Predatory mites Amblydromalus limonicus and Amblyseius herbicolus (Acari: Phytoseiidae) as potential biocontrol agents of Eotetranychus sexmaculatus (Acari: Tetranychidae) in avocado: examining predation on different prey life stages
Yuhao Yang, Keshi Zhang, Zhi-Qiang Zhang

TL;DR
This study explores two predatory mite species as potential natural enemies to control the six-spotted spider mite, a damaging pest in avocado plantations.
Contribution
The study is the first to evaluate the biocontrol potential of Amblydromalus limonicus and Amblyseius herbicolus against Eotetranychus sexmaculatus in avocado.
Findings
Both predator species fed and reproduced on all life stages of E. sexmaculatus.
Amblydromalus limonicus showed higher predation and oviposition rates than Amblyseius herbicolus.
Both predators preferred earlier life stages of the prey, with stronger preference in A. herbicolus.
Abstract
The six-spotted spider mite, Eotetranychus sexmaculatus (Riley) (Acari: Tetranychidae), is a severe pest of avocado trees, causing excessive leaf drop and reduced yield. Two predators found to be sympatric with E. sexmaculatus in surveys, Amblydromalus limonicus (Garman & McGregor) and Amblyseius herbicolus (Chant) (Acari: Phytoseiidae), may serve as potential biological control agents. We examined their potential in 2 laboratory experiments. (i) In no-choice tests, each predator was presented with 40 eggs, 40 larvae, 40 nymphs, or 20 female adults of E. sexmaculatus, and predation and oviposition rates (per day) were measured. (ii) In choice tests, predators were given a mixture of 10 eggs, 10 larvae, 10 nymphs, and 10 female adults, and the same parameters were measured, but with prey stage preference also assessed using Manly’s preference index. Our results showed both A. limonicus…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
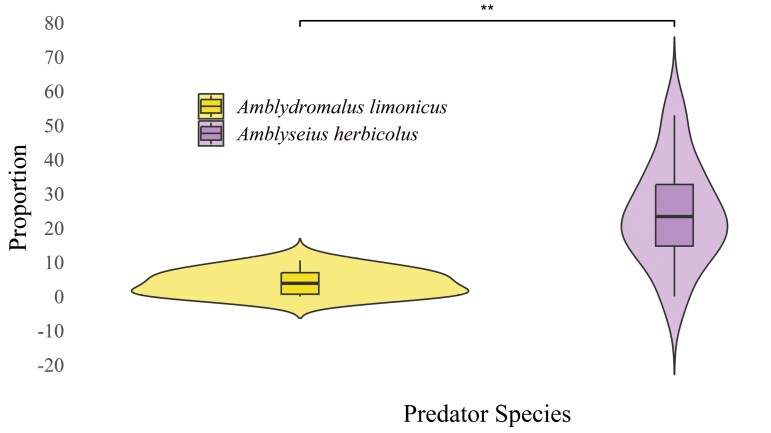 Fig. 1
Fig. 1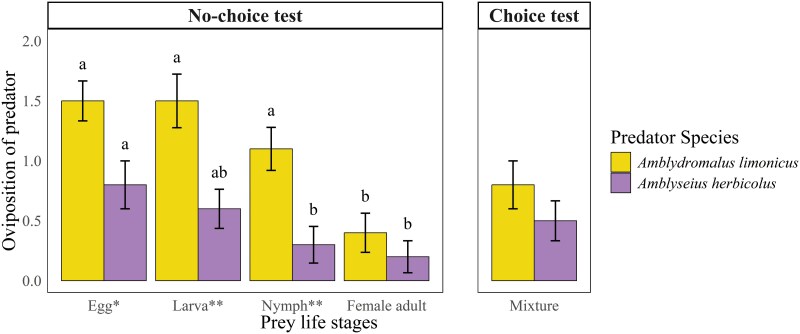 Fig. 2
Fig. 2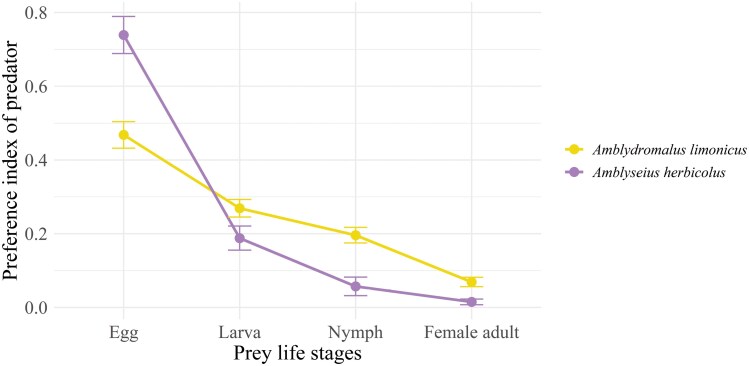 Fig. 3
Fig. 3| Prey | No. prey |
|
|
|---|---|---|---|
| Eggs | 40 | 38.8 ± 0.3 (37–40)a | 28.3 ± 2.2 (16–36)a |
| Larvae | 40 | 33.3 ± 1.4 (26–40)b | 20.8 ± 1.6 (14–28)b |
| Nymphs | 40 | 13.4 ± 0.9 (8–18)c | 9.2 ± 0.8 (5–14)c |
| Female adults | 20 | 4.8 ± 0.6 (2–8)d | 1.6 ± 0.2 (1–3)d |
| Prey | No. prey |
|
|
|---|---|---|---|
| Eggs | 10 | 7.6 ± 0.6 (4–10)a | 9.0 ± 0.4 (7–10)a |
| Larvae | 10 | 4.9 ± 0.3 (3–6)b | 3.2 ± 0.6 (0–6)b |
| Nymphs | 10 | 3.8 ± 0.4 (2–6)c | 1.1 ± 0.5 (0–5)c |
| Female adults | 10 | 1.4 ± 0.2 (0–2)d | 0.3 ± 0.1 (0–1)c |
- —Crown Research Institutes10.13039/501100019685
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect-Plant Interactions and Control · Insect and Pesticide Research · Insect Pheromone Research and Control
Introduction
Avocado (Persea americana Mill.) is a unique tropical and subtropical plant from Central America, with fruit that has gained popularity worldwide as a nutrient dense and healthy food source (Ayala Silva and Ledesma 2014, Talavera et al. 2023). The unique chemicals within avocado fruit and their derivatives are used in the manufacture of some industrial products, such as guacamole (Majid et al. 2020, García et al. 2021, Talavera et al. 2023). The direct and indirect damage to avocado quality caused by pests feeding on its fruit and leaves has caused serious economic losses to the avocado industry worldwide (Erichsen and Schoeman 1992, Hoddle et al. 2003, Ramírez-Gil et al. 2019, Torres et al. 2023). One phytophagous pest that feed on avocado leaves, the six spotted spider mite Eotetranychus sexmaculatus (Riley) (Acari: Tetranychidae), has had a serious negative impact on the avocado growing industry (Hoddle and Morse 2012, Bragard et al. 2023). Infestation of avocado trees by E. sexmaculatus usually occurs in late spring and early summer, and occasionally in autumn (Froud et al. 2002, Bragard et al. 2023). The pests are usually found on the lower surface of leaves, colonizing along the midrib and main veins of the leaf (McMurtry 1989, Lu et al. 2016). After infesting an avocado leaf, E. sexmaculatus feed on the phloem with their piercing sucking mouthparts, causing yellowing of the leaves and purple spots at the feeding sites. Feeding damage caused by E. sexmaculatus can lead to severe defoliation and a subsequent decline in avocado yield (Stevens et al. 2001, Jamieson and Stevens 2007).
Currently, the most used method of combating E. sexmaculatus damage to avocado plantations is chemical control with broad-spectrum pesticides, such as abamectin, milbemectin, or fenpyroximate (Froud et al. 2002, Steven 2004). Although chemical control works rapidly to reduce population densities at a low cost, the toxicity of broad-spectrum pesticides to natural enemies of E. sexmaculatus may lead to more serious management issues, such as the recolonization of E. sexmaculatus in the future, and the resistance of E. sexmaculatus to pesticides should also be considered (Tomkins 2002, Learmonth 2019). A recent report on the number of E. sexmaculatus on avocado leaves that were either sprayed or unsprayed with pesticides, confirms and reinforces these concerns about side effects on natural enemies and pesticide resistance (Logan et al. 2023). Although some studies have targeted plant resistance (including its molecular mechanisms) as another method for E. sexmaculatus control (Lu et al. 2016, Liang et al. 2020, Wang et al. 2023), these approaches have not been proven on avocados. Therefore, other available control methods are urgently needed.
Compared to other control strategies that have been utilized for spider mites, biological control is considered an environmentally friendly alternative (McMurtry et al. 2015, Knapp et al. 2018, Assouguem et al. 2022). Predatory mites in the family Phytoseiidae (Acari: Mesostigmata) are important biological control agents of many spider mite pest species on a variety of crops (McMurtry et al. 2013, Tixier 2018, Wang et al. 2024). Previous studies have confirmed that some phytoseiids are effective against E. sexmaculatus. For example, Hao et al. (2021), and Chen et al. (2023) found that Neoseiulus barkeri Hughes and Amblyseius swirskii Athias-Henriot were effective predators of E. sexmaculatus in rubber trees and were good candidate biological control agents for field releases. Caceres and Childers (1991) showed that Galendromus helveolus (Chant) could develop to adults with E. sexmaculatus as the only food source on citrus crops in Florida. However, because they have not been commonly found in wild avocado leaves until now, their effectiveness and adaptablity in artificial release is in doubt (McMurtry et al. 2013, Learmonth 2019, Logan et al. 2023), and its potential threat for biological invasion is also worth considering (Babendreier 2007, Hajek et al. 2016).
In a recent survey of E. sexmaculatus and its potential predators on sprayed and unsprayed “Hass” avocado trees in New Zealand, 2 predatory mite species were commonly sympatric with E. sexmaculatus: Amblydromalus limonicus (Garman & McGregor) (Acari: Phytoseiidae) and Amblyseius herbicolus (Chant) (Acari: Phytoseiidae) (Logan et al. 2023). Amblydromalus limonicus is a type III-b generalist predator that can feed and reproduce on a wide range of prey (McMurtry et al. 2013) and was reported as a potential predator of the phytophagous mite, Oligonychus punicae (Hirst) (Acari: Tetranychidae), as early as the 1960s (McMurtry and Scriven 1971). Compared to other predatory mites of the family Phytoseiidae, the commercialization of A. limonicus in the Americas, Europe and Oceania regions (Knapp et al. 2013, Schoeller et al. 2020), as well as its natural association with avocado leaves, makes it a good potential biocontrol agent against E. sexmaculatus on avocado. Previous studies have shown A. limonicus to be an effective biological control agents for thrips (Knapp et al. 2013, Lam et al. 2019, Schoeller et al. 2020, Mouratidis et al. 2023), psyllids (Liu et al. 2019), and whiteflies (Knapp et al. 2013, Cuthbertson 2014, Medd and GreatRex 2014, Lee and Zhang 2018), as well as some spider mite species (Knapp et al. 2013, Samaras et al. 2019, Wang et al. 2024). Its potential role as a biological control agent against E. sexmaculatus is unknown and worth investigating. Amblyseius herbicolus is also a type III-c generalist predator (McMurtry et al. 2013), and it is the most common predatory sympatric species for E. sexmaculatus in avocado orchards in New Zealand (Logan et al. 2023). In recent years, A. herbicolus has been shown to be a potential biological control agent for some pests, such as mites (Reis et al. 2007, Rodríguez-Cruz et al. 2013), whiteflies (Xin and Zhang 2021, Cardoso et al. 2024), psyllids (Jorge et al. 2021), and thrips (Lam et al. 2019). Fortunately, Zhang and Zhang (2021) provided a better method for large-scale cultivation of A. herbicolus by using Carpoglyphus lactis (Linnaeus) (Acari: Carpoglyphidae) as food, which is conducive for the potential large-scale commercialization of A. herbicolus.
Additionally, both A. limonicus and A. herbicolus can develop and reproduce on supplementary food, such as pollen, which makes it possible for predators to remain on avocado leaves when E. sexmaculatus population densities are low (Liu and Zhang 2017, Xin and Zhang 2021).
Although A. limonicus and A. herbicolus have been found to inhabit avocado leaves when E. sexmaculatus are present, it is unknown whether they can attack all stages of this prey. Therefore, this study simulates the artificial release of predators in a controlled laboratory environment with modified Munger cells and addresses the following objectives: (i) In no-choice tests, determine whether A. limonicus or A. herbicolus can prey on the single life stages of E. sexmaculatus and determine the rate of predation over 24 h. (ii) In choice tests, determine the predation preferences of A. limonicus or A. herbicolus for E. sexmaculatus at different life stages and the rate of predation over 24 h in an arena with mixed life stage of E. sexmaculatus. (iii) In both choice and no-choice tests, determine the number of eggs produced by A. limonicus or A. herbicolus, to assess whether these species can reproduce and thus demonstrate their potential as sustainable and environmentally friendly biological control agents.
Materials and Methods
Mite Cultures
Prey Mites
The dry fruit mite C. lactis was used for rearing predatory mites, and originated from a commercial supplier (Bioforce Limited, Karaka, Auckland, New Zealand) (Zhang and Zhang 2021). The prey was reared in 250 ml plastic containers with screw caps fitted with meshed holes for ventilation. The rearing medium consisted of a mixture of wheat bran, dry yeast, and icing sugar. The cultures were maintained in a controlled environment at 25 °C ± 1 °C, 80% ± 5% relative humidity (RH), and a 16:8 h light: dark photoperiod at the Manaaki Whenua—Landcare Research, St Johns, Auckland, New Zealand.
The initial population of E. sexmaculatus used as prey in this study was collected on avocado leaves from an orchard in Katikati, Bay of Plenty, New Zealand (−37.5307815, 175.9171147) and maintained on the leaves of avocado seedlings (c. 1 m in height) at 25 °C ± 3 °C, 40% ± 5% RH, under natural daylight. The avocado leaves were gently washed with water before introducing E. sexmaculatus. To prevent contamination from other predators, a thick layer of Vaseline was applied to the base of the avocado seedlings.
Predatory Mites
Both predatory mite species used in this study were originally collected on avocado and plum leaves from Rototuna, Hamilton, New Zealand in March 2024. The 2 predator species were reared using identical rearing set-ups: a petri dish was placed on top of a plastic sheet, which was then placed on a slightly larger piece of sponge (see Wang et al. 2024 for details). The sponge was soaked in water-filled containers. Mites had access to water but were unable to escape from their cultures, preventing contamination. The wheat bran mixture with C. lactis was added to the rearing set-ups of predatory mites. Four square plastic sheets (c. 10 mm long), folded twice, were added to each culture to provide refuge for the predatory mites.
For the experiments, similarly aged cohorts of both predatory species were established by transferring eggs (<24 h old) into new cultures with ad libitum supply of C. lactis. The eggs of predatory species were collected by placing sewing threads (c. 15 mm long) + into the main cultures overnight (Zhang and Zhang 2022). All adults of both species used in the experiments were young adults (less than 7 d after reaching maturity).
Experimental Arenas
For the experimental arenas used in the pretest starvation treatment, the cell consisted of 2 transparent plexiglass slides (L38 × W25 × H2 mm), and the aperture in this cell was conical (a diameter of 6 mm at the top, 10 mm at the bottom). A black plastic sheet was placed between 2 plexiglass slides, then 4 layers of filter paper were placed under the plastic sheet for adding water, and 7 holes were pierced in the plastic sheet using a size 3 insect pin to allow access to water for the predators (see Wang et al. 2024 for details). The entire cell assembly was secured using a pair of metal clips. A layer of food wrap was used to seal the top of the cylindrical hole. Seven evenly spaced holes were pierced into the wrap using a size 0 insect pin to allow ventilation.
The arena used in predation bioassays was similar to the cell described above but used transparent plexiglass square slides (L38 × W38 × H3 mm) and a cylindrical aperture with a diameter of 15 mm in the middle of the upper slide, which was the arena space for testing. A fresh avocado leaf disc (lower leaf surface upward) with a diameter of 20 mm was used instead of the black plastic sheet, and the freshness of this leaf disc ensured by adding water to the filter paper.
Experiment 1: No-choice Tests
Each no-choice test lasted for 24 h and occurred in the same controlled laboratory environment as the predator cultures. Each predatory species was replicated 10 times for each prey stage treatment. Before trials began, an adult (confirmed gravid) female (<7 d old) was transferred into an empty arena and starved for 24 h to standardize the predator’s hunger level; water was added to the 4 layers of filter paper before the test. Then, before the end of starvation treatment, one of 4 prey stage treatments was applied as follows: (i) 40 eggs, (ii) 40 larvae, (iii) 40 nymphs, or (iv) 20 adult females. Starved predators were placed in the center of the test arena and the top of the cell was immediately sealed with the piece of food wrap, and water was added to 4 layers of filter paper. The following data were recorded: (i) number of prey consumed; (ii) number of eggs laid by the predator; (iii) number of new eggs laid by female adult prey in a treatment group (to compare the differences in possible non-feeding effects on E. sexmaculatus oviposition due to A. limonicus and A. herbicolus); and (iv) the status of eggs in the egg treatment group (ie possible partial consumption, referring to an egg which was destroyed but not completely consumed).
Experiment 2: Choice Tests
The choice tests used the same set-up as the no-choice tests and lasted for 24 h, but the prey treatments included a mixture of 10 eggs, 10 larvae, 10 nymphs, and 10 female adults in each experimental arena. As in the no-choice test, there were 10 replicates for each predator species for each treatment. At the end of test, the same information as the no-choice test was recorded.
Statistical Analysis
The software R, Version 4.4.0 (R Core Team 2024) was used for statistical analysis in this study. The results were summarized as means and standard errors of the mean (SEMs). Due to the non-normal distribution of the whole data recorded, aligned Rank Transform (ART) analysis of variance (ANOVA) in the package ARTool (Kay et al. 2021) was used to compare the number of prey consumed by different treatment groups, the number of eggs laid by predators, and the number of eggs laid by prey female adults. A Wilcoxon rank-sum test was used to compare all the numerical differences in this study of the 2 predators under the same treatment.
The package selectapref (Richardson 2020) was used to generate the Manly’s preference index (Manly 1974) in the choice test (Experiment 2), which was used to assess the preference of A. limonicus and A. herbicolus for different prey stages. Using the preference index for eggs as an example, the formula is given below:
where α refers to the preference index; e1, e2, e3, and e4 refer to the numbers of consumed E. sexmaculatus eggs, larvae, nymphs and female adults, respectively; A1, A2, A3, and A4 refer to the initial numbers of E. sexmaculatus placed in the cell at the start of each test (in this study, A1, A2, A3, and A4 each = 10). A higher α value represents the higher preference of the predator for that prey’s life stage. The package ggplot2 (Wickham 2016) was used for graphical representation. Statistical significance was set at P < 0.05.
Results
Experiment 1: No-choice Test
Consumption of each of the 4 prey stages (ie eggs, larvae, nymphs, and adults) was observed in both predator species. Overall, adult A. limonicus females consumed significantly more prey than adult A. herbicolus females (ART ANOVA: F1,72 = 93.92, *P *< 0.001; Table 1). Prey consumption by both predator species was significantly affected by the prey life stage (F3,72 = 186.71, *P *< 0.001), with the highest consumption observed for eggs (Table 1). There was a significant interaction between predator species and prey life stage (F3,72 = 7.42, *P *< 0.001): the difference in prey consumption between A. limonicus and A. herbicolus was more pronounced for the earlier prey life stages than later stages (Table 1). Specially, the comparison of the results of 2 treatment groups of predators feeding on prey eggs showed that some of the eggs were not completely consumed by both predators, and the proportion of eggs partially consumed by A. limonicus was significantly lower than that for A. herbicolus (Fig. 1). Additionally, in the treatment group involving 20 adult E. sexmaculatus females, there were a small number of replicates, n = 3 (A. limonicus) and n = 2 (A. herbicolus), in which the predators completely sucked up one newly laid E. sexmaculatus egg.
*Experiment 1 (no-choice) egg treatment group data showing the proportion of prey eggs partially consumed by Amblydromalus limonicus and Amblyseius herbicolus out of all prey eggs consumed over a 24-h period. Asterisks denote the significant differences between two predator species (Wilcoxon rank-sum test: *P < 0.01). The “cloud” represents the kernel density estimation (KDE), showing the smoothed distribution of the data. The width of the “cloud” reflects the density of the data (the wider the clouds, the more possible data point estimates, or vice versa).
When feeding on E. sexmaculatus adult females, A. limonicus consumed more individuals than A. herbicolus (Table 1). However, the difference between 2 predators’ interference with oviposition rates of the remaining adult E. sexmaculatus females during tests was not significant (F1,18 = 0.56, P = 0.464): the number of eggs laid by adult E. sexmaculatus females in the 2 treatment groups were 0.94 ± 0.10 eggs in A. limonicus groups; and 1.10 ± 0.12 eggs in A. herbicolus groups.
Experiment 2: Choice Test
When adult females of A. limonicus and A. herbicolus were given a mixture of prey life stages, while the numbers of prey eggs/larvae consumed by 2 predator species was not significantly different (Table 2), the difference in overall consumption was significant (F1,72 = 15.56, P < 0.001). The consumption of prey at different life stages by both A. limonicus and A. herbicolus was highly significant (F3,72 = 94.12, P < 0.001), with higher consumption of prey at earlier life stages (Table 2). The interaction between predator species and prey life stage was also significant (F3,72 = 7.62, P = 0.012); this was reflected in A. herbicolus’ stronger preference for eggs than A. limonicus (Table 2).
Oviposition Rates in Choice and no-choice Bioassays
Amblydromalus limonicus laid significantly more eggs than A. herbicolus (F1,72 = 22.90; P < 0.001) in treatments of all 4 prey life stages in both the choice and no-choice experiments (Fig. 2). The number of eggs laid by A. limonicus and A. herbicolus in the experimental arena was significantly affected by the life stage of the prey (F3,72 = 10.77; P < 0.001), with this number gradually decreasing as the life stage of the prey increased (Fig. 2). However, there were no significant interactions between predator species and prey stage (F3,72 = 1.26; P = 0.296).
The number (Mean ± SEM) of eggs laid by Amblydromalus limonicus and Amblyseius herbicolus in the arena during no-choice (left) and choice (right) tests. Asterisks denote significant differences in oviposition rate within the same prey life stage between two species of predatory mites (Wilcoxon rank-sum test: ** P < 0.01; * P < 0.05). ab Means ART ANOVA pairwise comparisons in oviposition rate within the same species of predatory mites, different letters denote significant differences.
Most females of both predator species laid eggs when feeding on a mixture of prey life stages, but the difference in the number of eggs laid was non-significant between the 2 predator species (Fig. 2). As in the no-choice tests, there was no significant difference in the oviposition rates by the remaining adult E. sexmaculatus females in the 2 treatment groups (F1,18 = 1.05, P = 0.319): (1.10 ± 0.11 eggs in A. limonicus groups; 0.93 ± 0.11 eggs in A. herbicolus groups).
In this choice test, the calculated Manly’s preference index indicated a preference for earlier prey life stage by both A. limonicus and A. herbicolus (F3,72 = 105.43; P < 0.001; Fig. 3). There was a significant interaction between predator species and prey life stage (F3,72 = 19.27; P < 0.001), reflected in the stronger preference for eggs shown by A. herbicolus compared to A. limonicus (Fig. 3).
Choice test (Experiment 2): Manly’s preference index curves for adult females of Amblydromalus limonicus and Amblyseius herbicolus consuming different life stages of the prey Eotetranychus sexmaculatus. Bars show mean ± SEM values.
Discussion
In this study, we demonstrated for the first time that both A. limonicus and A. herbicolus can feed on the eggs, larvae, nymphs, and adult females of E. sexmaculatus on avocado leaf discs and that both species preferred earlier life stages of E. sexmaculatus. We also showed that both predator species could reproduce when feeding on different life stages of E. sexmaculatus. These results provide the first laboratory evidence that these 2 predatory species have the potential for use in biological control against E. sexmaculatus.
Predation
In the no-choice tests (Experiment 1), we observed that A. limonicus consumed significantly more prey than A. herbicolus across all E, sexmaculatus life stages (Table 1). Within the limits of the amount of prey being tested, young female A. limonicus adults were able to consume almost all the 40 E. sexmaculatus eggs or most of the 40 E. sexmaculatus larvae within 24 h, although the consumption might be greater if more prey eggs/larvae were provided. In the egg treatment, we observed that some prey eggs were partially consumed by A. limonicus or A. herbicolus, with such eggs being more frequently observed when A. herbicolus was the predator. This could indicate that the eggs of E. sexmaculatus might not provide sufficient nutrients to A. herbicolus. However, partially consumed eggs cannot hatch, so A. herbicolus still had some biological control effect on these E. sexmaculatus eggs. In phytoseiid-spider mites’ predator–prey interactions, the rate of prey consumption usually decreases with increasing life stage, although sometimes larval consumption is higher than egg consumption (Ali et al. 2011). In both E. sexmaculatus nymph and female adult treatment groups, the consumption rate by A. limonicus and A. herbicolus seemed to decrease more in comparison with egg and larva treatment groups (Table 1). Additionally, as E. sexmaculatus grows in size with subsequent life stages (McGregor 1950), the webs produced by E. sexmaculatus nymphs and adults seemed to hinder and entangle A. limonicus and A. herbicolus, reducing their rates of predation (Pers, observation of Y. Yang). This is consistent with previous reports of these 2 predators being hampered by webs from other spider mites (Sabelis and Bakker 1992, Wang et al. 2024). Although the prey in the female adult treatment groups laid some new eggs, the predators rarely consumed these eggs for food. This may also be because the newly laid eggs were protected by webs, making it difficult for predators to reach them.
In the choice test (Experiment 2), the predation efficiency of A. limonicus appeared to be similar to that of the no-choice test over the same test time (24 h), but the efficiency of A. herbicolus seemed to decrease (Table 2). Compared to other choice tests for spider mites, the predation efficiency of A. limonicus and A. herbicolus in this study was obviously lower than that reported in Hao et al. (2021). There are 4 possible explanations for this difference. (i) Differences exist in predation efficiency among different genera of predators. (ii) The host influences on the performance of E. sexmaculatus may be different on rubber versus avocado leaves (Lu et al. 2016, Liang et al. 2020). (iii) The 3-dimensional and relatively closed set-ups of the test arena simulated in this study might strengthen the effects of E. sexmaculatus webs in limiting predator mobility (Sabelis and Bakker 1992, Shimoda et al. 2010). (iv) Compared with the study of Hao et al. (2021), the higher prey density provided by this study might interfere with the predator’s hunting efficiency (Reis et al. 2003).
Prey Preference
In the choice test (Experiment 2), A. limonicus and A. herbicolus clearly favored earlier prey stages, which is similar to the results reported for A. swirskii (Hao et al. 2021). The preference was more marked in A. herbicolus. Many studies have investigated the prey preferences of species in the family Phytoseiidae, and variable nutritional value of the different predation strategies in the face of different spider mite prey and thus show different predation preferences (Blackwood et al. 2001, Xiao and Fadamiro 2010, Jyothis and Ramani 2019, 2024). Possible factors include the probability of encountering prey (Sabelis 1990), how easy it is to hold onto the prey (Furuichi et al. 2005), and the availability of nutritional value at different stages of the prey’s life (Sabelis 1990, Blackwood et al. 2001, Xiao and Fadamiro 2010). Different predation preferences may also be related to the degree to which predators are specialized for hunting different prey (Blackwood et al. 2001, McMurtry et al. 2013). The possible interference from the E. sexmaculatus webs with predator behavior may have changed the predation preferences of A. limonicus or A. herbicolus in this study, consistent with several previous studies (Sabelis and Bakker 1992, Shimoda et al. 2010, McMurtry et al. 2013). In no replicate in the choice test, did we find even one newly laid egg consumed by predators, but the eggs which were artificially set without the protection of webs were heavily consumed. Active prey could leave the protection of the webs and be consumed; thus, the number of prey remaining seems to reflect the ability of prey to detect the predator and hide in the protection of the webs. Under this assumption, A. herbicolus might be less successful in capturing and consuming E. sexmaculatus than A. limonicus, resulting in A. herbicolus having to consume more artificially set eggs for survival and showing more extreme preferences.
Oviposition Rate
In this study, young female predators in the no-choice experiments laid an average of 1.5 (A. limonicus) and 0.8 (A. herbicolus) eggs, respectively, when provided with the eggs of E. sexmaculatus, but the rate of oviposition decreased with increasing prey life stage (Fig. 2). In the choice experiments, young female predators laid an average of 0.8 (A. limonicus) and 0.5 (A. herbicolus) eggs, respectively (Fig. 2). Our follow-up observations showed that these eggs were able to hatch. Oviposition rates reflect the ability of a predator to reproduce using the nutrients in its prey, and it can be used to predict whether artificially released predators can control E. sexmaculatus prey effectively and sustainably in the avocado leaf habitat. Compared to the other predation tests, the oviposition rate of A. limonicus consuming E. sexmaculatus in our study was similar to that of A. limonicus when feeding on other suitable prey species in other laboratory studies (Davidson et al. 2016, Patel and Zhang 2017, Lam et al. 2019), which seems to indicate that A. limonicus can effectively use and convert the nutrients of E. sexmaculatus into offspring. However, the oviposition rate of A. herbicolus consuming E. sexmaculatus was significantly lower than that of A. herbicolus when consuming other suitable prey species in other laboratory studies (Lam et al. 2019, Liu et al. 2024). This seems to suggest that A. herbicolus is not well suited to consuming E. sexmaculatus as prey for total nutrition alone. This makes it is uncertain whether A. herbicolus, if released, would choose to consume E. sexmaculatus, other prey, or possibly even A. herbicolus larvae (Liu et al. 2023) as the food source.
A further consideration is that our review of previous studies on A. limonicus found that the daily oviposition rate reported in studies using the same prey in long-term rearing was significantly higher than the reported single-day oviposition rate of studies simulating artificial release (McMurtry and Scriven 1965, Samaras et al. 2015). This was consistent with the interference of the previous prey on subsequent short-term oviposition rate, as reported in Sabelis (1990).
Predator Lifestyle
In this study, A. limonicus and A. herbicolus, 2 biological control agents commonly used against thrips, psyllids, and whiteflies, showed statistically significantly different results against the spider mite E. sexmaculatus. Differences in lifestyle may explain this: A. limonicus, a type III-b predator, prefers glabrous leaves as a habitat, while A. herbicolus, a type III-c predator, prefers confined space on dicotyledonous plants (McMurtry et al. 2013). Therefore, the less hairy and more open avocado leaves may be more suitable for A. limonicus. This is also consistent with our observations when collecting predators. The location from where we collected A. limonicus and A. herbicolus included neighboring avocado and plum trees: most A. limonicus were observed on avocado leaves, while most A. herbicolus observed on plum leaves.
Limitations of Current Study and Suggestion for Future Studies
This study was a laboratory simulation of predator release in the field, and, as such, certain factors confound interpretation. (i) The test arena was more enclosed than the avocado leaves, which could have led to excessive interference from predators with the 3-dimensional webbing structure; (ii) The experimental set-up involved simpler interspecific interactions than natural niches, and predators may prefer to consume prey other than E. sexmaculatus (eg A. herbicolus may prefer thrips) after release in the field. (iii) The higher density of prey in our laboratory study probably interfered with the predator’s living space. Expanded greenhouse experiments or field experiments should then be conducted as follow ups to provide better evidence of whether these predators can act as effective biological control agents.
Overall, this study demonstrates that A. limonicus and A. herbicolus can consume E. sexmaculatus at various life stages on avocado leaves. Of the 2 predators, A. limonicus, which has been commercialized, can be more quickly and easily applied to actual avocado production. However, the webs of E. sexmaculatus may be a key factor in reducing the predation efficiency of both predators. Future studies will need to consider the extent to which the webs hinder A. limonicus or A. herbicolus, and whether both of these predators can be used as practical biological control agents in integrated pest management programs.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ali M , Naif AA, Huang D. 2011. Prey consumption and functional response of a phytoseiid predator, Neoseiulus womersleyi, feeding on spider mite, Tetranychus macfarlanei. J. Insect Sci. 11:167. https://doi.org/10.1093/jis/11.1.167 · doi ↗
- 2Assouguem A , Kara M, Mechchate H, et al 2022. Current situation of Tetranychus urticae (Acari: Tetranychidae) in Northern Africa: the sustainable control methods and priorities for future research. Sustainability. 14:2395. https://doi.org/10.3390/su 14042395 · doi ↗
- 3Ayala Silva T , Ledesma N. 2014. Avocado history, biodiversity and production. In: Nandwani D editor. Sustainable horticultural systems. sustainable development and biodiversity. Vol 2. Springer. p. 157–205.
- 4Babendreier D. 2007. Pros and cons of biological control. In: Nentwig W editor. Biol. invasions. Springer. p. 403–418.
- 5Blackwood J , Schausberger P, Croft B. 2001. Prey-stage preference in generalist and specialist phytoseiid mites (Acari: Phytoseiidae) when offered Tetranychus urticae (Acari: Tetranychidae) eggs and larvae. Environ. Entomol. 30:1103–1111. https://doi.org/10.1603/0046-225X-30.6.1103 · doi ↗
- 6Bragard C , Baptista P, Chatzivassiliou E, et al; EFSA Panel on Plant Health (EFSA PLH Panel). 2023. Pest categorisation of Eotetranychus sexmaculatus. EFSA J. 21:e 07898. https://doi.org/10.2903/j.efsa.2023.789837009445 PMC 10052452 · doi ↗ · pubmed ↗
- 7Caceres S , Childers CC. 1991. Biology and life tables of Galendromus helveolus (Acari: Phytoseiidae) on Florida citrus. Environ. Entomol. 20:224–229. https://doi.org/10.1093/ee/20.1.224 · doi ↗
- 8Cardoso AC , Marcossi I, Fonseca MM, et al 2024. A predatory mite as potential biological control agent of Bemisia tabaci on tomato plants. J. Pest Sci. 1–13. https://doi.org/10.1007/s 10340-024-01809-7 · doi ↗