Application of hsp60 amplicon sequencing to characterize microbial communities associated with juvenile and adult Euprymna scolopes squid
Steph Smith, Clotilde Bongrand, Susannah Lawhorn, Edward G Ruby, Alecia N Septer

TL;DR
This study uses hsp60 sequencing to better understand the microbial communities in squid, showing that Vibrio fischeri is the main symbiont in their light organs.
Contribution
The study introduces hsp60 amplicon sequencing as a method to improve species-level resolution of squid-associated bacterial communities.
Findings
Vibrio fischeri dominates the light organ microbiome of E. scolopes, making up 94%-99% of amplicon sequence variants.
Non-V. fischeri taxa like Bradyrhizobium and Rhodobacterales are consistently present in both juvenile and adult squid.
hsp60 sequencing allows for non-invasive assessment of microbial communities in squid ventate samples.
Abstract
The symbiotic relationship between Vibrio (Aliivibrio) fischeri and the Hawaiian bobtail squid, Euprymna scolopes, serves as a key model for understanding host–microbe interactions. Traditional culture-based methods have primarily isolated V. fischeri from the light organs of wild-caught squid, yet culture-independent analyses of this symbiotic microbiome remain limited. This study aims to enhance species-level resolution of bacterial communities associated with E. scolopes using hsp60 amplicon sequencing. We validated our hsp60 sequencing approach using pure cultures and mixed bacterial populations, demonstrating its ability to distinguish V. fischeri from other closely related vibrios and the possibility of using this approach for strain-level diversity with further optimization. This approach was applied to whole-animal juvenile squid exposed to either seawater or a clonal V.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
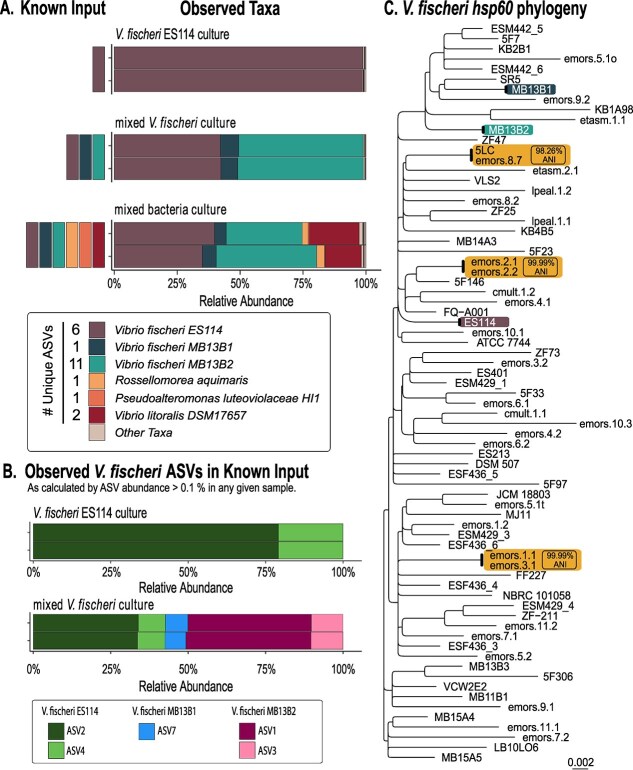 Figure 1
Figure 1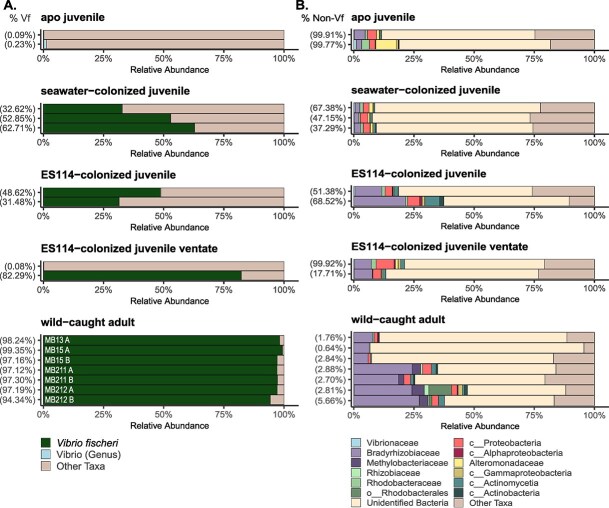 Figure 2
Figure 2- —UNC EMES Mills Brown Fellowship Award
- —National Science Defense and Engineering Graduate fellowship (NDSEG)
- —NIGMS10.13039/100000057
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCephalopods and Marine Biology · Gut microbiota and health · Parasite Biology and Host Interactions
Brief communication
Symbiosis model systems have been essential to expanding our understanding of how microbes interact with their hosts. The symbiosis between Vibrio fischeri and Euprymna scolopes (Hawaiian bobtail squid) is a model for studying host-bacterial interactions in the context of a beneficial association. Juvenile E. scolopes hatch without their bacterial symbiont, V. fischeri, which they acquire from the surrounding seawater. One or more distinct V. fischeri strains quickly colonize the bi-lobed symbiotic light organ, comprised of six physically separated epithelium-lined crypt spaces. After the cells proliferate in the crypts, the bacterial symbionts’ bioluminescence aids in the squid’s nocturnal activity [1]. Each day, the squid vents 90% of the bacterial cells within these colonized crypts into the surrounding water, resulting in a “ventate water” that can be collected and used to non-invasively assess the symbiotic population.
Although V. fischeri is the only species isolated from the light organs of wild-caught animals, only one study has applied a culture-independent approach for examining the diversity of bacteria associated with adult animals. This study, which applied amplicon sequencing of the V3–V4 region of the 16S rRNA gene to the light organs of two wild-caught adult squid, found the majority of reads (79%) were most similar to V. fischeri, with another 5% most similar to Vibrio litoralis [2]. Although this study was the first to apply a culture-independent approach to the light organ symbiosis, it also revealed a need for using an amplicon target that better distinguishes between Vibrio species, to increase confidence for assigning species, and possibly strain-level taxonomy to amplicon sequence variants (ASVs).
This work investigates the application of hsp60 amplicon sequencing to achieve enhanced taxonomic resolution in the vibrio-squid symbiosis, particularly for distinguishing closely related symbiotic populations with the potential for non-invasive sampling approaches. The hsp60 gene encodes a conserved protein that serves as a better marker for distinguishing and identifying species-level classification within the closely related Vibrionaceae [3, 4]. We applied this approach to pure bacterial cultures, as well as whole-animal juvenile E. scolopes squid, juvenile vented water, and light organ cores from four wild-caught adults. Based on these results, we propose how this approach can be applied to the vibrio-squid system, its limitations, and how further optimization could expand its utility.
We first applied hsp60 amplicon sequencing to known culture samples. DNA extraction and amplicon sequencing were performed on three types of culture samples: (i) a single strain of V. fischeri, (ii) mixed cultures of multiple V. fischeri strains, and (iii) a multi-species culture, to validate that V. fischeri can be distinguished from other species and observe how strain-level variance might appear in the amplicon sequencing analysis. Because this approach involves amplification of the target gDNA sequence, amplification or sequencing errors could introduce nucleotide changes and thus result in distinct ASVs from a single, clonal copy of the target, or natural within-population variation of the target sequence could result in multiple ASVs, despite representing a single bacterial lineage [5–7].
Analysis of the culture samples indicates that V. fischeri ASVs can be distinguished from ASVs for other species (Fig. 1A), while strain-level resolution was less clear (Fig. 1B). Although most V. fischeri genomes encode a unique hsp60 sequence (Fig. 1C), including the strains used in our culture samples (ES114, MB13B1, and MB13B2), the hsp60 amplicon sequencing results yielded more than one unique ASV per input strain. At the species level, all 18 unique ASVs were assigned to V. fischeri. However, at the strain level, six unique ASVs were identified as ES114, 11 ASVs were identified as MB13B2, and one ASV was identified as MB13B1. When the dataset was filtered to exclude ASVs that represent <0.1% of total reads across any given sample, two dominant unique ASVs were identified for both ES114 and MB13B2, and a single unique ASV was identified for MB13B1 (Fig. 1B), suggesting that removal of low-abundance ASVs that may result from sequencing errors does not resolve the mismatch between the number of strains (three) and the number of unique ASVs (five) for the known culture samples.
hsp60 amplicon sequencing differentiates V. fischeri from other closely related vibrios at the species level. (A) Known input corresponds to bacterial isolates added to each known community, and observed taxa reports the relative abundance (%) of taxa identified from each known community using hsp60 amplicon sequencing. All known input strains were detected in their corresponding samples, including in mixed communities comprised of multiple V. fischeri strains and closely related vibrios. The number of unique ASVs that were assigned to each taxonomic identifier is reported here. (B) Relative abundance (%) of V. fischeri ASVs comprising >0.1% of total abundance in any given sample. Of ASVs that meet this criterion, two ASVs were identified as V. fischeri ES114, two ASVs were identified as V. fischeri MB13B2, and one ASV was identified as MB13B1. Abundance of any additional unique ASVs identified in panel a fell below the abundance cutoff of 0.1%. (C) Neighbor-joining phylogenetic tree analysis based on distance matrix calculated by multiple sequence alignment (MSA) of 73 V. fischeri hsp60 sequences obtained from NCBI and trimmed to the region amplified by the primers used in this study. Orange boxes represent strains with 100% identity between hsp60 sequences compared to the average nucleotide identity (ANI) between the corresponding strains at the whole-genome level as calculated by FastANI (98.26%–99.99% ANI).
We conclude that, although hsp60 amplicon sequencing can distinguish V. fischeri from other bacterial species, further work is needed to identify appropriate computational thresholds to separate true strain-level sequence variants from ASVs that may arise from errors in amplification or sequencing. Since we are currently unable to determine whether minority ASVs represent unique strains or artifacts of the approach, we chose not to apply strain-level diversity analyses to the animal samples in the following section.
We next applied hsp60 amplicon sequencing to animal samples. DNA extraction and amplicon sequencing was performed on three types of samples: (i) homogenized whole animal juveniles colonized with either Hawaiian seawater or a single V. fischeri strain, (ii) paired ventate water from juveniles colonized by a single V. fischeri strain, or (iii) paired cores of light organ lobes (A and B) from four wild-caught adult squid. Although only between 31% and 62% of the ASVs associated with juvenile squid were identified as V. fischeri, most remaining ASVs had no assigned taxonomy, making V. fischeri the majority of identified reads in these samples. While the majority V. fischeri ASVs in ES114-colonized juveniles confirms that these animals were symbiotic, the paired ventate sample results were variable: one sample returned 82% of reads identified as V. fischeri while the second ventate sample was comparable to juvenile squid that were not exposed to V. fischeri (aposymbiotic; apo) (Fig. 2A). These results suggest this approach could be optimized by plating a subsample of the ventate prior to extraction to verify the animals did in fact vent. For adult samples, V. fischeri ASVs comprised 94%–99% of the total reads, confirming that V. fischeri is the primary symbiont in the light organ using a culture-independent technique (Fig. 2A).
Relative abundance of V. fischeri and other conserved taxa associated with juvenile and adult squid. (A) Relative abundance of V. fischeri, other vibrios, and non-vibrio taxa in juvenile squid, juvenile squid ventate, and adult squid. The percentage of reads identified as V. fischeri in each sample is indicated on the y-axis (% Vf). In a wild-caught adult panel, MB indicates animal identifier, with A or B indicating paired samples from each lobe of the same light organ. (B) Corresponding relative abundance of non-V. fischeri taxa in the samples described in panel A. Taxa are plotted at the family level, and Vibrionaceae excludes ASVs identified as V. fischeri. Unidentified bacteria correspond to ASVs that were not assigned taxonomy past the kingdom level, likely due to a lack of similar sequences in available hsp60 reference sequences. Other taxa correspond to ASVs that were assigned taxonomy to at least the family level but make up <0.1% of the total read abundance. Sample order is consistent between panel A and panel B.*
Finally, we explored the ASVs that were not identified as V. fischeri in animal samples, which comprised 37%–68% of reads for juveniles and 0.6%–5% of reads for adult light organ cores. For juvenile squid, we employed whole-animal homogenization – an approach commonly used for culture-based quantification that typically yields almost exclusively V. fischeri colonies on selective media. However, our results demonstrate that culture-independent hsp60-amplicon sequencing captures a more diverse community from these same homogenates, reflecting both V. fischeri and other bacteria that may be associated with other tissues or external surfaces. This distinction indicates that in future studies, the choice between whole-animal homogenization and organ-specific sampling should be matched to specific research questions. For adult squid, our light organ core approach substantially reduced but did not eliminate non-V. fischeri ASVs, as this method would still capture bacteria associated with the light organ surface.
Analysis of these non-V. fischeri ASVs revealed two important findings: (i) the majority of these ASVs were not assigned a taxonomy (likely due to lack of similar sequences available in hsp60 databases), and (ii) several conserved taxa appeared in both juvenile and adult animals that were dominated by members of the Rhodobacterales order, including Bradyrhizobium spp. (Fig. 2B). Importantly, none of these conserved ASVs were detected in either of our negative extraction control (NEC) samples, which consisted of sterile PBS processed concurrently through DNA extraction, library preparation, and sequencing alongside experimental samples. This absence from negative controls strongly suggests these taxa represent genuine biological associations rather than contaminants introduced during sample processing. These conserved taxa may represent another important microbial association outside of the primary light organ symbiosis. Bradyrhizobium have primarily been studied as plant symbionts known for their ability to fix nitrogen, with a recent isolate encoding genes to assimilate ammonium [8], which is also excreted by the squid host [9, 10]. Future work may permit the cultivation of such taxa and the exploration of their potential localization and interactions with the squid host.
Methods
See supplemental methods for details pertaining to squid collection and experimentation, bacterial strains, DNA extraction, sequencing, and bioinformatic analyses.
Supplementary Material
Smith-2024_Supplemental_Final_ycaf085
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Nyholm SV, Mc Fall-Ngai MJ. A lasting symbiosis: how the Hawaiian bobtail squid finds and keeps its bioluminescent bacterial partner. Nat Rev Microbiol 2021;19:666–79. 10.1038/s 41579-021-00567-y 34089010 PMC 8440403 · doi ↗ · pubmed ↗
- 2Bongrand C, Koch E, Mende D. et al. Evidence of genomic diversification in a natural symbiotic population within its host. Front Microbiol 2022;13:854355. 10.3389/fmicb.2022.85435535300477 PMC 8922018 · doi ↗ · pubmed ↗
- 3Jesser KJ, Noble RT. Vibrio ecology in the Neuse River estuary, North Carolina, characterized by next-generation amplicon sequencing of the gene encoding heat shock protein 60 (Hsp 60). Appl Environ Microbiol 2018;84:e 00333–18. 10.1128/AEM.00333-1829678912 PMC 6007114 · doi ↗ · pubmed ↗
- 4King WL, Siboni N, Kahlke T. et al. A new high throughput sequencing assay for characterizing the diversity of natural vibrio communities and its application to a Pacific oyster mortality event. Front Microbiol 2019;10:2907. 10.3389/fmicb.2019.0290731921078 PMC 6932961 · doi ↗ · pubmed ↗
- 5Pan P, Gu Y, Sun D-L. et al. Microbial diversity biased estimation caused by Intragenomic heterogeneity and interspecific conservation of 16S r RNA genes. Appl Environ Microbiol 2023;89:e 02108–22. 10.1128/aem.02108-2237129483 PMC 10231250 · doi ↗ · pubmed ↗
- 6Schirmer M, Ijaz UZ, D’Amore R. et al. Insight into biases and sequencing errors for amplicon sequencing with the Illumina Mi Seq platform. Nucleic Acids Res 2015;43:e 37–7. 10.1093/nar/gku 134125586220 PMC 4381044 · doi ↗ · pubmed ↗
- 7Schloss PD . Amplicon sequence variants artificially split bacterial genomes into separate clusters. m Sphere 2021;6:e 00191–21. 10.1128/m Sphere.00191-2134287003 PMC 8386465 · doi ↗ · pubmed ↗
- 8Paudel D, Liu F, Wang L. et al. Isolation, characterization, and complete genome sequence of a Bradyrhizobium strain Lb 8 from nodules of peanut utilizing crack entry infection. Front Microbiol 2020;11:93. 10.3389/fmicb.2020.0009332117123 PMC 7020250 · doi ↗ · pubmed ↗