Phylogeny, phylogeography, and conservation of a rediscovered gecko from the Galápagos Islands
Omar Torres-Carvajal, Paula A. Castaño, Enrique Rincón, Fernando Ayala-Varela, Karl Campbell, Wilson Cabrera, Francisco Moreno

TL;DR
Scientists rediscovered a rare gecko species on Rábida Island in the Galápagos and studied its evolutionary history and conservation status.
Contribution
The study presents the first vouchered specimens of Phyllodactylus on Rábida and identifies them as a distinct evolutionary significant unit.
Findings
Vouchered specimens of Phyllodactylus were collected for the first time on Rábida Island.
Rábida gecko populations are recognized as a separate evolutionary significant unit within P. maresi.
The eradication of invasive rodents likely contributed to the rediscovery of the gecko.
Abstract
The rediscovery of species in iconic conservation sites like the Galápagos is eye-catching and can lead to quick and effective conservation actions. With 11 species occurring on most islands, Galápagos leaf-toed geckos (Phyllodactylus) are among the least known terrestrial vertebrates of the archipelago. Strikingly, reliable records of Phyllodactylus from Rábida Island prior to this study are limited to Holocene subfossils and a single photograph from 2012. Here we report the first vouchered specimens of Phyllodactylus from Rábida and present their phylogenetic and phylogeographic affinities with other gecko populations in the archipelago. Despite taxonomic uncertainty, we recognize Rábida gecko populations as a separate ESU within P. maresi, which also occurs in nearby islands (Santiago, Bartolomé, Mares), as well as in Marchena. Finally, we suggest that Rábida gecko populations…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
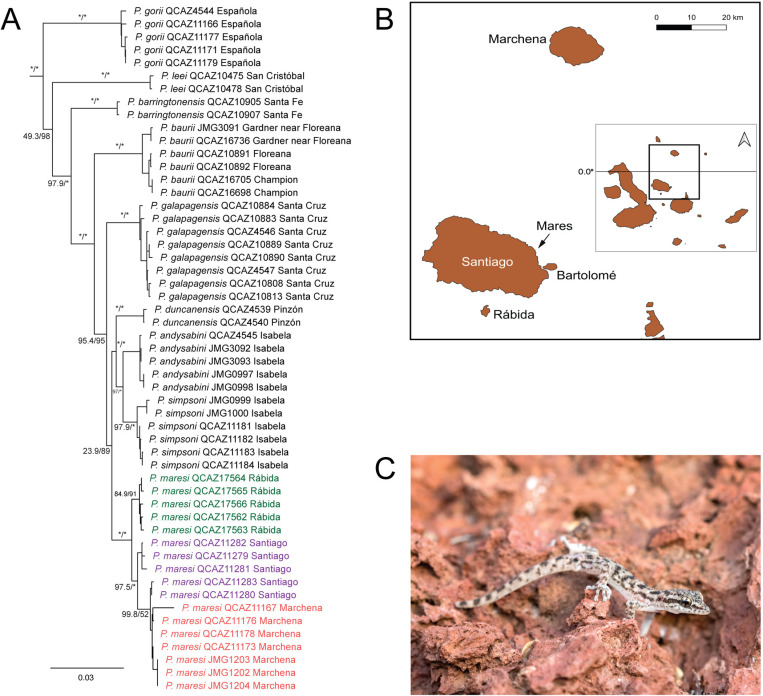 Fig 1
Fig 1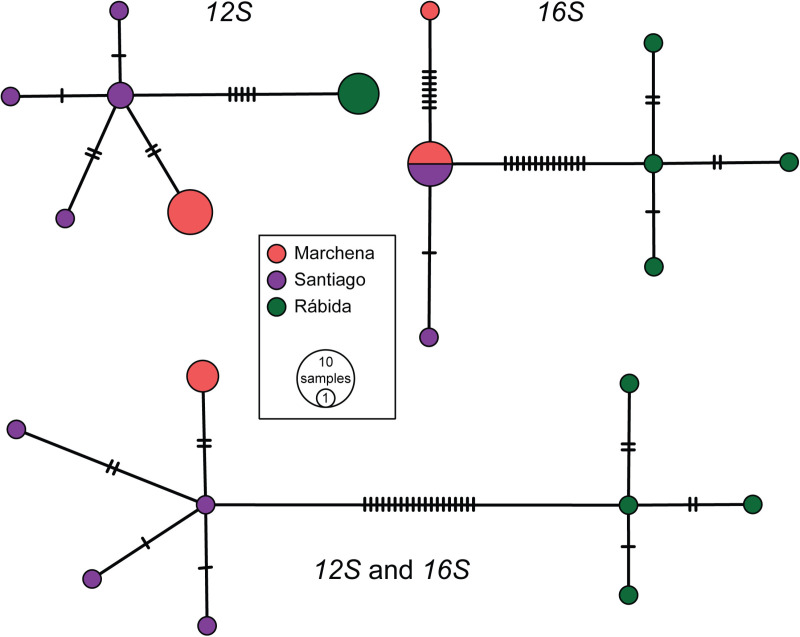 Fig 2
Fig 2- —http://dx.doi.org/10.13039/501100011749Pontificia Universidad Católica del Ecuador
- —Island Conservation
- —http://dx.doi.org/10.13039/501100016404Galapagos Conservation Trust
- —Galapagos National Park Directorate
- —Re:wild
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSpecies Distribution and Climate Change · Insect and Arachnid Ecology and Behavior · Plant and animal studies
Introduction
The rediscovery of species previously thought to be extinct has received considerable attention in recent years [1,2]. Some of the factors influencing rediscovery are (1) range and population size, (2) distribution within tropical megadiverse countries, (3) incorrect declaration of “extinct” status, (4) lifeform (plants), (5) accessibility to habitats of lost species, (6) species’ charisma, and (7) search effort [1–4]. The rediscovery of species in iconic conservation areas, such as the Galápagos Islands, attracts special attention and can result in quick conservation actions. For example, soon after the rediscovery in 2019 of the giant tortoise Chelonoidis phantasticus in Fernandina Island, the only female found, Fernanda, was transferred to the Galápagos National Park Tortoise Center, and further expeditions aimed at finding males for captive breeding were initiated [5].
In contrast to the charismatic giant tortoises, many Galápagos organisms have received less attention. Due to their secretive habits and nocturnal activity, leaf-toed geckos (Phyllodactylus) are perhaps the least known terrestrial vertebrates from the Galápagos Archipelago. Their present diversity stems from three colonization events originating in South America [6]. Of these, two events (~0.69 Mya and ~3.03 Mya) led to a single extant species (Phyllodactylus giberti in Wolf Island and P. darwini in San Cristobal Island, respectively). The third and oldest radiation (P. galapagensis radiation), which started at ~5.49 Mya, resulted in nine species across most of the islands [7,8]. Interestingly, despite intense collecting efforts in the Galápagos, especially in the late 19^th^ and early 20^th^ centuries [9–11], no geckos were registered from one of its main islands, Rábida (a.k.a. Jervis). This 499-ha island emerged between 1.3–1.6 million years ago [12] and lies in the central part of the archipelago, about 4.5 km south of Santiago Island and 25 km east of Isabela Island. Prior to 2012, the only record of geckos from this island were Holocene subfossil bones (5,700–8,540 years B. P.) reported by Steadman et al. [13], who also mentioned that “the current absence on Rábida of the secretive Phyllodactylus may only be an artifact of inadequate collecting.” Subsequently, in their field guide to Galápagos vertebrates, Swash and Still [14] reported Phyllodactylus from Rábida as an undescribed, possibly extinct species without providing further details. Thus, leaf-toed geckos in Rábida were suspected of having gone extinct prior to humans discovering the archipelago.
In 2011, the Galápagos National Park and its conservation partners successfully implemented an invasive rodent eradication program targeting brown rats (Rattus norvegicus) [15]. Shortly after removing invasive rodents, a team of researchers and Galápagos National Park rangers monitoring Rábida Island in 2012 found a specimen of Phyllodactylus [15]. This specimen was collected and photographed, thus representing the first record in the literature of a living leaf-toed gecko from Rábida Island. Subsequent reports of Phyllodactylus from Rábida by other authors [14,16] are not reportedly based on vouchers, photographs or even sightings. In this paper, we report on the rediscovery of leaf-toed geckos from Rábida based on two recent expeditions that allowed us to collect both tissue samples and specimens. We present for the first time their phylogenetic and phylogeographic affinities to other gecko populations from the Galápagos based on analyses of DNA sequence data, as well as a general morphological characterization and comparison with related taxa.
Materials and methods
Fieldwork and data sampling
Samples were obtained during two expeditions to Rábida Island. In October 2019, we sampled muscle tissue (tail tips) from nine specimens of leaf-toed geckos from Rábida and stored those samples in Longmire buffer [17] before releasing the specimens. These tissue samples were deposited at Museo de Zoología QCAZ, Pontificia Universidad Católica del Ecuador, Quito (QCAZ 17562–570). In a second expedition between 5–7 August 2021, we collected 10 whole body gecko specimens, which were captured by hand between 18:00–23:00 h and photographed the next morning. Most specimens were active on lava rocks. After lethal anesthetization with an intracelomic injection of benzocaine (2%), muscle and liver tissue samples were extracted and stored in Longmire buffer. Specimens were then fixated in 10% formalin and stored in 70% ethanol in the Vertebrate Collection of the Charles Darwin Foundation in Puerto Ayora, Galápagos (VCCDRS 3415–3424). This study was evaluated and approved by the DINV (Dirección de Investigación) of the Pontificia Universidad Católica del Ecuador in accordance with the guidelines for environmental and social impacts of research projects. The DINV committee evaluates projects to determine observance of its norms for ethical scientific research. Genetic data were obtained under the Genetic Resources Access Contract No MAE-DNB-CM-2016–0060 issued by the Ecuadorian Ministry of Environment, Water, and Ecological Transition. Science permit PC-56–23 and export permits 159–2019 DPNG and 070–2023 DPNG were issued by the Galápagos National Park Directorate (DPNG).
DNA sequence data and phylogenetic analyses
Tissue samples were mixed with Proteinase K and lysis buffer before overnight digestion. Total genomic DNA was extracted following a guanidinium isothiocyanate extraction protocol. DNA samples were quantified with a Nanodrop® ND-1000 (NanoDrop Technologies, Inc.), re-suspended and diluted to 25 ng/ul in ddH_2_O prior to amplification. We used the same primers and amplification protocols described in Torres-Carvajal et al. [8].
We obtained new DNA sequence data from five samples of Phyllodactylus (QCAZ 17562–566) from Rábida. We combined those sequences with 130 GenBank sequences of 23 species of Phyllodactylus from Galápagos and continental South America [6,8,18]. The species Phyllodactylus nocticolus, P. unctus and P. xanti were used as outgroups to root the tree [8]. We analyzed a character matrix of 135 terminals and 5,086 aligned nucleotides (nt) encompassing four mitochondrial and six nuclear genes. Mitochondrial genes included the NADH dehydrogenase subunit 4 (ND4, 601 nt) and a continuous fragment encompassing 12S rRNA, tRNA^Val^, and 16S rRNA (1,995 nt). Nuclear genes included brain-derived neurotrophic factor (BDNF, 630 nt), oocyte maturation factor MOS (CMOS, 390 nt), recombination-activating gene 1 (RAG1, 304 nt), recombination-activating gene 2 (RAG2, 384 nt), acetylcholinergic receptor M4 (ACM4, 383 nt) and phosducin (PDC, 399 nt). Sequences generated in this study are available on GenBank (Table 1). Although we were not able to obtain new sequences for ND4, we included this gene in the analyses because it is phylogenetically informative [8] and available in GenBank for most taxa included in our analyses.
Table 1: GenBank accession numbers for sequences obtained in this study.
We assembled and aligned DNA sequences in GENEIOUS PRIME 2022.1. (https://www.geneious.com) under default settings for MAFFT [19]. We translated all protein-coding sequences into amino acids for confirmation of alignment. In addition, we realigned ribosomal gene regions with multiple gaps in an attempt to minimize indels and optimize nucleotide identities across different individuals.
Phylogenetic relationships were inferred under the maximum likelihood optimality criterion in IQ-TREE [20]. We used MODELFINDER [21] to infer models of evolution for each gene, except for the tRNA^Val^ gene, which was included in the same partition as 12**S [8]. Branch support was also assessed in IQTREE with 1,000 replicates under both ultrafast bootstrapping [22] and SH approximate likelihood ratio test (SH-aLRT) [23]. We also ran a partitioned Bayesian analysis in MRBAYES v3.2.7 [24], with all parameters unlinked between partitions (except topology and branch lengths) and rate variation (prset ratepr = variable) invoked. We performed four independent runs of 5 million generations, each with four MCMC chains, sampling every 1,000 generations. After verifying convergence and effective sample sizes (≥ 200) in TRACER v1.7 [25], we discarded the initial 500 trees per run as “burn-in” before calculating posterior probabilities on a consensus tree.
Phylogeography
To investigate haplotype diversity between leaf-toed geckos from Rábida and other related populations, we calculated haplotype networks for mitochondrial genes 12S and 16S using the software PopArt [26]. Because sequences of both 12S and 16S were either incomplete or not available for some samples, we trimmed individual 12S (n = 16, 315 nt) and 16S (n = 12, 1,259 nt) matrices until all sequences had < 5% of missing data. We also analyzed a concatenated 12S-16S dataset after removing samples missing either 12S or 16S sequences (n = 11, 1,574 nt). We used the median-joining haplotype network algorithm, with parameter epsilon set to 0 [27]. We also addressed genetic differentiation within the Phyllodactylus galapagensis radiation [8] by calculating uncorrected genetic distances for 12S and 16S in PhyML 3.3 [23] using DIVEIN [28].
Morphological data
We studied external morphology in all specimens obtained in the 2021 expedition and used published data [11] for comparisons with closely related taxa. We followed terminology for measurements and scutellation from the literature [11,29,30] to record the following characters: (1) number of scales between nostril and eye including the one or two scales on anterior margin of preorbital groove; (2) number of scales across snout at level of third labials; (3) number of scales across interorbital region at level of center of eyes; (4) number of supralabials from rostral to a point below center of eye; (5) number of infralabials from mental to a point below center of eye; (6) number of postmentals; (7) number of scales in contact with postmentals posteriorly; (8) number of scales in contact with scales behind postmentals; (9) number of longitudinal rows of dorsal tubercles at midbody; (10) number of dorsal tubercles counted on the medialmost (paravertebral) row from head to base of tail; (11) number of paravertebral tubercles between axilla and groin; (12) number of scales around midbody; (13) number of lamellae on Finger IV; (14) number of lamellae on Toe IV; (15) eye diameter; (16) head length between snout and anterior margin of ear opening; (17) snout length between snout and anterior margin of orbit; (18) length between axilla and groin; (19) snout–vent length (SVL); and (20) tail length (TL). SVL and TL measurements were taken with a ruler and recorded to the nearest millimeter. Other measurements were made with digital calipers and recorded to the nearest 0.1 mm. Sex was determined by gonad inspection through dissection, or by noting the presence or absence of (1) hemipenes by eversion or (2) shelled eggs visible through the skin.
Results
The topologies of the maximum likelihood and Bayesian trees obtained in this study (Fig 1 and S1 Fig, respectively) are very similar and support previous hypotheses [8,18]. All species of Phyllodactylus, except for P. darwini and P. gilberti are nested in a single clade, the “galapagensis clade” [6]. The new specimens of Phyllodactylus from Rábida form a strongly supported clade sister to P. maresi from Santiago and Marchena with maximum support. Therefore, we used all available samples from Rábida, Santiago and Marchena in the phylogeographic analyses.
Phylogenetic relationships of leaf-toed geckos (Phyllodactylus) from Rábida, Galápagos Islands.A. Maximum-likelihood phylogenetic tree of leaf-toed geckos (Phyllodactylus) from Galápagos. Only the P. galapagensis radiation (i.e., all Galapagean species except P. darwini-San Cristóbal and P. gilberti-Wolf) is shown for clarity; the full tree is shown in S2 Fig. New samples from Rábida are shown in green. Numbers next to branches are SH-aLRT/ultrafast bootstrap support values, with asterisks representing values ≥ 99%; numbers on short branches were removed for clarity. B. Map of the Galápagos archipelago showing islands included in this study. The approximate location of Mares Islet is indicated with an arrow. This map was created using the Free and Open Source QGIS; base map and data from OpenStreetMap and OpenStreetMap Foundation. C. Phyllodactylus maresi from Rábida (photograph by Fernando Ayala-Varela, Bioweb.bio).
The inferred networks show a close genetic similarity between populations from Marchena and Santiago; however, samples from Rábida are more divergent (Fig 2). The concatenated 12S-16S network shows four haplotypes from Santiago separated from each other by 1–2 mutations, one haplotype from Marchena separated by two mutations from Santiago, and four haplotypes from Rábida separated from each other by 1–2 mutations and from Santiago by 19 mutations. The 12S-only network is different in that Rábida contains a single haplotype, separated from Santiago by four mutations. The 16S-only network shows a shared haplotype between Santiago and Marchena, separated from single haplotypes from Santiago and Marchena by one and seven mutations, respectively; haplotypes from Rábida are separated from the shared Santiago/Marchena haplotype by 14 mutations.
Haplotype networks of leaf-toed geckos (Phyllodactylus) from Rábida, Santiago and Marchena islands.The haplotype networks are based on both individual 12S and 16S genes and a concatenated dataset. Bars on branches indicate mutation steps. The colors match the clades in Fig 1.
Mean 12S and 16S interspecific genetic distances (S1 and S2 Tables, respectively) among species of the P. galapagensis radiation vary between 0.02 (P. duncanensis/ P. simpsoni) and 0.11 (e.g., P. maresi/ P. leei, P. gorii/ P. baurii), and 0.01 (P. simpsoni/ P. andysabini) and 0.10 (e.g., P. duncanensis/ P. leei, P. galapagensis/ P. gorii), respectively. The mean 12S genetic distance is 0.01 between specimens of P. maresi from Santiago and Marchena, 0.01 between Santiago and Rábida, and 0.02 between Marchena and Rábida. The mean 16S genetic distance is 0.002 between specimens of P. maresi from Santiago and Marchena, 0.01 between Santiago and Rábida, and 0.01 between Marchena and Rábida.
The specimens of Phyllodactylus from Rábida are similar in morphology and color pattern to those from Santiago and Mares islet (Table 2). Specimens from Rábida are more similar to those from Santiago in having less paravertebral tubercles and less scales around midbody. Given the lack of DNA samples from Mares, lack of specimens from Marchena, and generally low sample sizes, we assign the samples of leaf-toed geckos from Rábida to P. maresi based on our phylogenetic analysis (Fig 1).
Table 2: Geographical variation in morphology of Phyllodactylus maresi from Mares, Santiago and Rábida islands.
Discussion
The Galápagos Archipelago represents an iconic conservation site. Therefore, it is crucial for conservation practice that biodiversity data is based on evidence instead of anecdote [31], and that conservation interventions (e.g., removal of invasive species) have a robust monitoring program [32] that can provide attribution of changes detected to specific interventions. This may include changes in density, distribution, recording new species or rediscovering others. The recent discovery of new species of land iguanas, giant tortoises, snakes, and lizards [8,33–35], as well as the rediscovery of species that were thought to be extinct [5], underscores the fact that the task of documenting the biodiversity of terrestrial vertebrates in the Galápagos is far from complete. The best scenario to overcome this problem in common species like lava lizards and leaf-toed geckos is to collect whole-organism voucher specimens with associated DNA samples in areas determined by expert taxonomists. Except for endangered species or populations, whole organism specimens are essential for species descriptions and conservation actions [36]. Although leaf-toed geckos have been collected in many islands across the Galápagos archipelago [7,9–11], records from Rábida have only been anecdotal [14,16], except for subfossils [13] and a photograph of a specimen recorded in 2012 [15]. The 2012 specimen, although collected, does not appear in collections where it was reported as deposited and now appears to be lost. Here we confirm the existence of Phyllodactylus in Rábida through collection of voucher specimens as part of the monitoring program following the 2011 invasive rodent eradication program. Teams (including authors KC, WC) searching for geckos were unable to find geckos or any evidence of them (e.g., footprints, shed skin, egg shells) during and prior to 2011. Lizard populations subject to rodent predation in the Caribbean and offshore islands of New Zealand have recovered after rodent eradication [37,38]. Although the ecological relationships between rodents and geckos in Rábida are unknown, we suggest that populations of Phyllodactylus benefited from the eradication of rodents in Rábida and that population recovery and expanded distribution due to rodent eradication facilitated their rediscovery and the collection efforts reported here.
Our phylogenetic analyses suggest that the population of Phyllodactylus from Rábida corresponds to P. maresi, a species that was recently elevated from subspecies status by Arteaga et al. [16]. Although we accept P. maresi as a valid species, we argue that Arteaga et al.‘s proposal is questionable because they did not include in their analyses samples from Mares islet (Fig 1), the type locality of P. maresi [11]. In a comprehensive phylogenetic study of Phyllodactylus from the Galápagos, Torres-Carvajal et al. [8] recognized a clade with six samples from Santiago and Marchena as “P. sp. 2”. Upon addition of mitochondrial gene sequences of three samples from Marchena, Arteaga et al. [16] obtained a similar topology and assigned arbitrarily populations from Marchena, Santiago and the surrounding Rábida, Bartolomé and Mares Islet to Phyllodactylus maresi. However, it is worth mentioning that Lanza [11] proposed the subspecies P. galapagensis maresi precisely because he noticed morphological differences between specimens from Mares and Santiago. We found similar differences between specimens from Rábida and Mares (Table 2). Strikingly, there are only five whole-body voucher specimens of Phyllodactylus from Mares, three from Santiago and no specimens from Marchena in museums worldwide as far as we know. Therefore, further and possibly more refined (e.g., genomic data) analyses based on additional sampling is needed to elucidate the taxonomic status of leaf-toed geckos from Marchena, Rábida, Santiago, Mares and surrounding islets. Phylogenomic analyses are particularly promising as demonstrated by a previous study on Phyllodactylus geckos from Mexico that revealed substantial cryptic diversity [39].
The delimitation of evolutionarily significant units (ESUs)—divergent intraspecific lineages [40]—is crucial for the conservation of insular species, which are represented by relatively small and isolated populations that are generally more prone to extinction than their mainland relatives partly because of limited genetic variation [41]. Genetic variability in Phyllodactylus is lower in the Galápagos than in mainland South America [8]; therefore, recognition of ESUs are important for the conservation of these lizards. Torres-Carvajal et al. [42] recognized two ESUs within P. baurii, one occurring in Floreana and Champion, and the other from Gardner. Despite taxonomic uncertainty, both our phylogenetic and phylogeographic analyses suggest that leaf-toed geckos from Rábida represent a separate ESU from the Marchena and Santiago populations (Figs 1 and 2).
Supporting information
S1 Table12S uncorrected pairwise distances among species of the Phyllodactylus galapagensis radiation.Number of sequences (N) of each taxon, mean, standard error, minimum (Min), Q1, median, Q3, and maximum (Max) values are presented.(XLSX)
S2 Table16S uncorrected pairwise distances among species of the Phyllodactylus galapagensis radiation.Number of sequences (N) of each taxon, mean, standard error, minimum (Min), Q1, median, Q3, and maximum (Max) values are presented.(XLSX)
S1 FigBayesian phylogenetic tree of leaf-toed geckos (Phyllodactylus).Numbers on branches are posterior probability values.(PDF)
S2 FigMaximum-likelihood phylogenetic tree of leaf-toed geckos (Phyllodactylus).Numbers on branches are SH-aLRT/ultrafast bootstrap support values.(PDF)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ladle R, Jepson P, Malhado AC, Jennings S, Barua M. The causes and biogeographical significance of species’ rediscovery. Front Biogeogr. 2011;3(3):111–8.
- 2Lindken T, Anderson CV, Ariano-Sánchez D, Barki G, Biggs C, Bowles P, et al. What factors influence the rediscovery of lost tetrapod species? Global Change Biology. 2024;30(1):e 17107.10.1111/gcb.1710738273552 · doi ↗ · pubmed ↗
- 3Scheffers BR, Yong DL, Harris JBC, Giam X, Sodhi NS. The world’s rediscovered species: back from the brink? P Lo S One. 2011;6(7):e 22531. doi: 10.1371/journal.pone.0022531 21818334 PMC 3144889 · doi ↗ · pubmed ↗
- 4Humphreys AM, Govaerts R, Ficinski SZ, Nic Lughadha E, Vorontsova MS. Global dataset shows geography and life form predict modern plant extinction and rediscovery. Nat Ecol Evol. 2019;3(7):1043–7. doi: 10.1038/s 41559-019-0906-2 31182811 · doi ↗ · pubmed ↗
- 5Jensen EL, Gaughran SJ, Fusco NA, Poulakakis N, Tapia W, Sevilla C, et al. The Galapagos giant tortoise Chelonoidis phantasticus is not extinct. Commun Biol. 2022;5(1):546. doi: 10.1038/s 42003-022-03483-w 35681083 PMC 9184544 · doi ↗ · pubmed ↗
- 6Torres-Carvajal O, Rodríguez-Guerra A, Chaves JA. Present diversity of Galápagos leaf-toed geckos (Phyllodactylidae: Phyllodactylus) stems from three independent colonization events. Mol Phylogenet Evol. 2016;103:1–5. doi: 10.1016/j.ympev.2016.07.006 27400628 · doi ↗ · pubmed ↗
- 7Wright JW. The evolution and biogeography of the lizards of the Galápagos Archipelago: evolutionary genetics of Phyllodactylus and Tropidurus populations. In: Bowman RI, Berson M, Levinton AE, editors. Patterns of evolution in Galápagos organisms. San Francisco, CA: American Association for the Advancement of Science, Pacific Division; 1983. p. 123–55.
- 8Torres-Carvajal O, Barnes CW, Pozo‐Andrade MJ, Tapia W, Nicholls G. Older than the islands: origin and diversification of Galápagos leaf‐toed geckos (Phyllodactylidae: Phyllodactylus) by multiple colonizations. J Biogeogr. 2014;41(10):1883–94.