When the heat is on, HYL1 steps in: Regulating miRNA biogenesis for plant thermotolerance
Regina Mencia

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
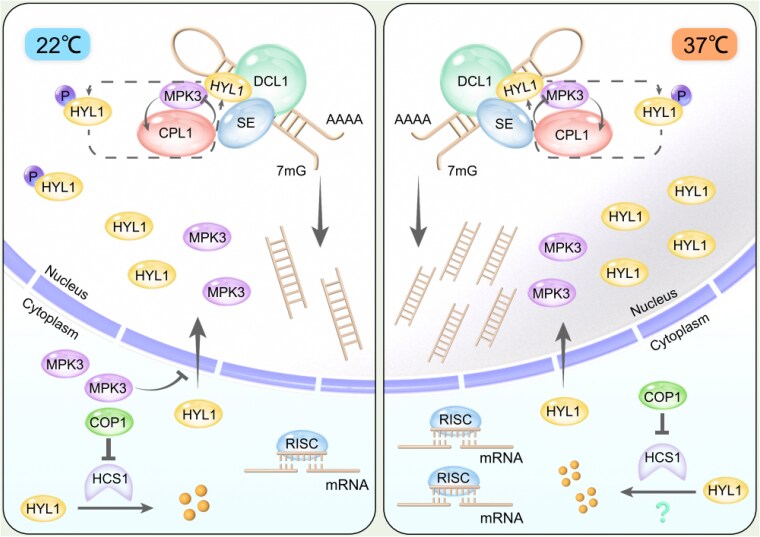 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Light effects on plants · Photosynthetic Processes and Mechanisms
Changes in environmental temperature are a major source of stress for plants. Plants have evolved various adaptive strategies that combat heat stress (HS) and mitigate potential negative effects on development and productivity. One such strategy involves microRNAs (miRNAs), small single-stranded RNAs, typically 21 nucleotides long, that play key regulatory roles, primarily through post-transcriptional gene silencing. In this process, miRNAs, once loaded into a protein complex, recognize target transcripts, leading to their cleavage and subsequent degradation (Huntzinger and Izaurralde 2011).
While specific miRNAs have been linked to HS responses, Yiming Cao, Jiaxin Zhang, and colleagues (Cao et al. 2025) set out to explore the broader, global role of miRNAs in plant adaptation to HS, using Arabidopsis (A. thaliana) as a model organism. Their findings reveal that most miRNAs are upregulated under HS, leading to the downregulation of their target genes. However, an intriguing observation emerged: pri-miRNA (hairpin-folded primary miRNA transcript) levels did not follow the same pattern.
MicroRNA synthesis is a multi-step process. In the nucleus, the core microprocessor complex comprising DICER-LIKE1 (DCL1), SERRATE (SE), and HYPONASTIC LEAVES 1 (HYL1) recognizes pri-miRNAs and processes them into mature miRNAs (Mencia et al. 2023). The observed discrepancy between pri-miRNA and mature miRNA levels suggests that HS may influence the global miRNA biogenesis pathway rather than just individual miRNAs.
A closer look at the miRNA biogenesis pathway revealed a surprising finding: although overall HYL1 levels decreased under HS, its nuclear localization increased. The routing of HYL1 to the nucleus led to greater binding to pri-miRNAs in HS-treated plants, which could explain the higher miRNA accumulation due to increased microprocessor activity. Furthermore, mutants with defects in the microprocessor displayed greater sensitivity to HS than wild-type plants, underscoring the crucial role of miRNA biogenesis in HS responses.
It was previously shown that nuclear localization of HYL1 is orchestrated by its phosphorylation state (Gonzalo et al. 2024). This led the authors to investigate whether HS could affect HYL1 phosphorylation, thereby triggering increased miRNA biogenesis. Analysis of mutant plants carrying a nonphosphorylated version of HYL1 revealed reduced nuclear localization, lower miRNA induction, and increased sensitivity to HS, further supporting the central role of HYL1 phosphorylation in the HS response.
HYL1 phosphorylation is regulated by the dynamic balance between the phosphatase CPL1 and the kinase MPK3 (Gonzalo et al. 2024). In their study, Cao et al. (2025) examined the interaction between CPL1 and MPK3, determining that the 2 proteins physically interact. Additionally, they found that MPK3 phosphorylates CPL1, regulating its stability. Interestingly, mutants lacking CPL1 exhibited increased MPK3 kinase activity, suggesting a reciprocal regulatory mechanism between these 2 proteins (Figure). Moreover, when analyzing mutant plants lacking MPK3, they observed increased nuclear accumulation of HYL1, higher miRNA abundance, and enhanced tolerance to HS. Overall, the work by Cao et al. provides valuable insights into the role of miRNA biogenesis in HS adaptation, shedding light on the regulation of HYL1 and its contribution to plant stress responses.
Recent related articles in The Plant Cell
Li et al. (2023) demonstrate that HS induces the miR165/166–PHB regulatory module, which modulates HSFA1, controlling HS responses in Arabidopsis thaliana through transcriptional reprogramming and antagonistic interactions between PHB and HSFA1s. Field et al. (2023) review recent advances in understanding biomolecular condensates in plant development, examining their potential roles throughout the plant life cycle, their molecular functions, and how they respond to environmental changes. Pan et al. (2024) revealed that the SMXL4/SMXL5–HSFA2 regulatory module modulates plant thermotolerance by repressing HSFA2 transcription and antagonizing HSFA1 activity, while HSFA2, in turn, induces SMXL4 and SMXL5 expression during heat stress recovery, establishing a feedback mechanism in Arabidopsis thaliana. Park et al. (2023) showed that HYL1 mediates the ABA-induced recruitment of the HOS15-HDA9 complex to MIRNA loci, linking pri-miRNA-structure recognition to the transcriptional repression of miRNA biogenesis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cao Y, Zhang J, Zhao Z, Tang G, Yan J. Heat stress triggers enhanced nuclear localization of HYL 1 to regulate mi RNA biogenesis and thermotolerance in plants. Plant Cell. 2025. 10.1093/plcell/koaf 092PMC 1214259040266261 · doi ↗ · pubmed ↗
- 2Field S, Jang G-J, Dean C, Strader LC, Rhee SY. Plants use molecular mechanisms mediated by biomolecular condensates to integrate environmental cues with development. Plant Cell. 2023:35(9):3173–3186. 10.1093/plcell/koad 06236879427 PMC 10473230 · doi ↗ · pubmed ↗
- 3Gonzalo L, Giudicatti AJ, Manavella PA. HYL 1's multiverse: a journey through mi RNA biogenesis and beyond canonical and non-canonical functions of HYL 1. Curr Opin Plant Biol. 2024:80:102546. 10.1016/j.pbi.2024.10254638718678 · doi ↗ · pubmed ↗
- 4Huntzinger E, Izaurralde E. Gene silencing by micro RN As: contributions of translational repression and m RNA decay. Nat Rev Genet. 2011:12(2):99–110. 10.1038/nrg 293621245828 · doi ↗ · pubmed ↗
- 5Li J, Cao Y, Zhang J, Zhu C, Tang G, Yan J. The mi R 165/166–PHABULOSA module promotes thermotolerance by transcriptionally and posttranslationally regulating HSFA 1. Plant Cell. 2023:35(8):2952–2971. 10.1093/plcell/koad 12137132478 PMC 10396384 · doi ↗ · pubmed ↗
- 6Mencia R, Gonzalo L, Tossolini I, Manavella PA. Keeping up with the mi RN As: current paradigms of the biogenesis pathway. J Exp Bot. 2023:74(7):2213–2227. 10.1093/jxb/erac 32235959860 · doi ↗ · pubmed ↗
- 7Pan Y, Yu B, Wei X, Qiu Y, Mao X, Liu Y, Yan W, Linghu Q, Li W, Guo H, et al Suppression of SMXL 4 and SMXL 5 confers enhanced thermotolerance through promoting HSFA transcription in Arabidopsis. Plant Cell. 2024:36(10):4557–4575. 10.1093/plcell/koae 22439102897 PMC 11449109 · doi ↗ · pubmed ↗
- 8Park J, Giudicatti AJ, Bader ZE, Han MK, Møller C, Arce AL, Xu Z-Y, Yang SW, Manavella PA, Yun D-J. The HIGH EXPRESSION OF OSMOTICALLY RESPONSIVE GENE 15-HISTONE DEACETYLASE 9 complex associates with HYPONASTIC LEAVES 1 to modulate micro RNA expression in response to abscisic acid signaling. Plant Cell. 2023:35(8):2910–2928. 10.1093/plcell/koad 13237195876 PMC 10396366 · doi ↗ · pubmed ↗