Pumping iron, keeping FIT: How MYB30 regulates FIT stability during plant iron deficiency
Linhan Sun

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
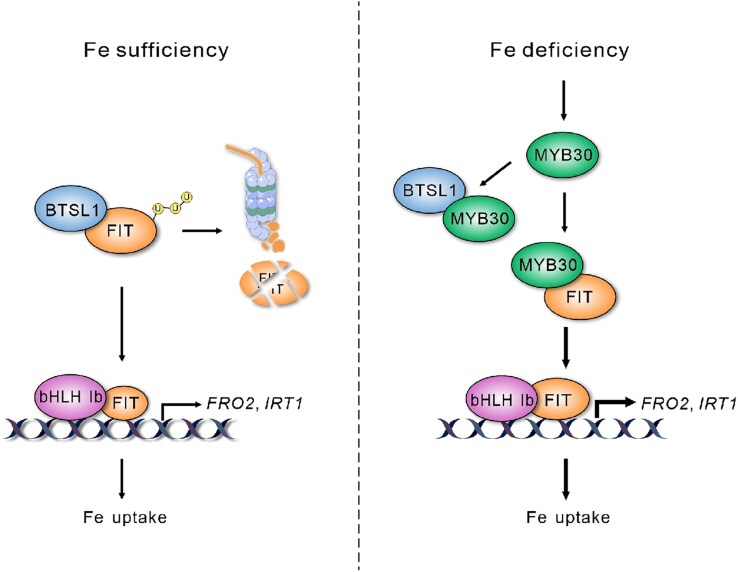 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Micronutrient Interactions and Effects · Banana Cultivation and Research
The well-known cartoon character Popeye eats spinach for its high iron (Fe) content, which gives him the strength to deal with his nemesis, Brutus. Plants also rely on this critical element to grow properly. In the model plant Arabidopsis (A. thaliana), a pivotal transcription factor FER-LIKE IRON DEFICIENCY-INDUCED TRANSCRIPTION FACTOR (FIT) helps plants respond to Fe deficiency by inducing the expression of iron transporter genes, including FERRIC REDUCTASE OXIDASE2 (FRO2) and IRON TRANSPORTER1 (IRT1; Colangelo and Guerinot 2004). When Fe stores are replenished, 2 BRUTUS-LIKE E3 ubiquitin ligases, BTSL1 and BTSL2, mediate the ubiquitination and degradation of FIT, thus preventing excessive iron uptake (Rodríguez-Celma et al. 2019). Surprisingly, even under Fe deficiency, the expression of BTSL1 and BTSL2 also increases (Rodríguez-Celma et al. 2019). This paradox raises the question of how FIT escapes the fate of being degraded under Fe deficiency, when it is needed the most.
In new work, Hongyun Zhao and colleagues (Zhao et al. 2025) reveal a role for another transcription factor, MYB30, in the stabilization of FIT during Fe deficiency. First, they observed that 2 MYB30 T-DNA insertion loss-of-function mutants (myb30-1 and myb30-2) exhibited more severe Fe deficiency phenotypes, relative to the wild type, such as stronger inhibition of root elongation and diminished expression of FRO2 and IRT1. Conversely, the overexpression lines (OX-#1 and OX-#2) showed enhanced resistance to Fe deficiency stress and elevated levels of FRO2 and IRT1. The mRNA transcript and protein levels of MYB30 were also significantly upregulated under Fe deficiency. These results pointed to the role of MYB30 as a positive regulator of Fe deficiency. To further explore how MYB30 is involved in this process and identify other regulators that work in concert with MYB30, the authors performed a yeast 2-hybrid screening of MYB30-interacting proteins and identified BTSL1 and BTSL2 as candidate interacting partners. Using protein-protein interaction assays, the authors further confirmed that the N-terminus of MYB30 (containing its DNA-binding domain) interacted with the C terminus region of BTSL1. The overlapping expression patterns of MYB30 and BTSL1/2 at the root suggested that MYB30 functions in concert with BTSL1/2 in the Fe-deficiency pathway.
Counterintuitively, the authors discovered that these 2 E3 ubiquitin ligases did not appear to mediate the degradation of MYB30 itself, indicating an unconventional link between BTSL1/2 and MYB30. Given the well-established interaction between BTSL1/2 and FIT (Rodríguez-Celma et al. 2019), the authors hypothesized that the missing link between BTSL1/2 and MYB30 could be FIT. Indeed, the authors discovered that FIT interacted with MYB30 both in yeast and in planta and that the N-terminal DNA-binding region of MYB30 alone could physically interact with FIT. The authors used a series of biochemical experiments to demonstrate that the absence of MYB30 resulted in accelerated degradation of FIT under Fe deficiency, while overexpression of MYB30 exhibited the opposite effects on FIT stability. More importantly, using in vivo co-immunoprecipitation experiments in both tobacco leaves and Arabidopsis protoplasts, they showed that the addition of MYB30 reduced the strength of the interaction between BTSL1 and FIT, and overexpression of MYB30 also curtailed the ubiquitination and degradation of FIT mediated by BTSL1. Collectively, these data suggested that MYB30 functions to interfere with the interaction between BTSL proteins and FIT, thus stabilizing FIT under iron deficiency conditions.
In this comprehensive work, Zhao et al. (2025) uncovered an exciting mechanism by which Arabidopsis responds to Fe deficiency. This response resembles the textbook “competitive inhibition” mechanism: When Fe levels are sufficient, BTSL targets FIT for degradation. However, when Fe levels drop, MYB30 functions as a “decoy” for BTSL1/2, protecting FIT from being degraded by BTSL1/2 under Fe-deficiency. Fine-tuning FIT protein levels and ensuring proper cellular responses mediated by FIT is at the heart of this response (Fig.). This work also opens the door for exploring similar mechanisms regulating the stability of other transcription factors, for example, other key bHLH transcription factors involved in the Fe-deficiency response that are targeted by other E3 ligases (Selote et al. 2015). When plants “pump iron,” they might be able to combat BRUTUS through different mechanisms but with similar underlying themes.
Recent related articles in The Plant Cell:
Zhao et al. (2024) reported that MYB30 also modulates nitric oxide (NO)-induced seed germination in Arabidopsis. Paffrath et al. (2024) showed that a group of secondary metabolites called coumarins is involved in the Fe reduction process in roots mediated by FERRIC REDUCTION OXIDASE2 (FRO2). Chia et al. (2023) reported on Arabidopsis OLIGOPEPTIDE TRANSPORTER 3 (OPT3), a Fe transporter localized in the phloem, that mediates systemic responses to both Fe deficiency and copper (Cu) deficiency, highlighting the complex crosstalk between Fe and Cu homeostasis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chia J-C, Yan J, Rahmati Ishka M, Faulkner MM, Simons E, Huang R, Smieska L, Woll A, Tappero R, Kiss A, et al Loss of OPT 3 function decreases phloem copper levels and impairs crosstalk between copper and iron homeostasis and shoot-to-root signaling in Arabidopsis thaliana. Plant Cell. 2023:35(6):2157–2185. 10.1093/plcell/koad 05336814393 PMC 10226573 · doi ↗ · pubmed ↗
- 2Colangelo EP, Guerinot ML. The essential basic helix-loop-helix protein FIT 1 is required for the iron deficiency response. Plant Cell. 2004:16(12):3400–3412. 10.1105/tpc.104.02431515539473 PMC 535881 · doi ↗ · pubmed ↗
- 3Paffrath V, Tandron Moya YA, Weber G, von Wirén N, Giehl RFH. A major role of coumarin-dependent ferric iron reduction in strategy I-type iron acquisition in Arabidopsis. Plant Cell. 2024:36(3):642–664. 10.1093/plcell/koad 27938016103 PMC 10896297 · doi ↗ · pubmed ↗
- 4Rodríguez-Celma J, Connorton JM, Kruse I, Green RT, Franceschetti M, Chen Y-T, Cui Y, Ling H-Q, Yeh K-C, Balk J. Arabidopsis BRUTUS-LIKE E 3 ligases negatively regulate iron uptake by targeting transcription factor FIT for recycling. Proc Natl Acad Sci U S A. 2019:116(35):17584–17591. 10.1073/pnas.190797111631413196 PMC 6717287 · doi ↗ · pubmed ↗
- 5Selote D, Samira R, Matthiadis A, Gillikin JW, Long TA. Iron-binding E 3 ligase mediates iron response in plants by targeting basic helix-loop-helix transcription factors. Plant Physiol. 2015:167(1):273–286. 10.1104/pp.114.25083725452667 PMC 4281009 · doi ↗ · pubmed ↗
- 6Zhao H, Jiang J, Shen M, Zhang Y, Zhang Y, Liu H, Zhou H, Zheng Y. The transcription factor MYB 30 promotes iron homeostasis by maintaining the stability of the FIT transcription factor. Plant Cell. 2025. 10.1093/plcell/koaf 090PMC 1212340640244930 · doi ↗ · pubmed ↗
- 7Zhao H, Ma L, Shen J, Zhou H, Zheng Y. S-nitrosylation of the transcription factor MYB 30 facilitates nitric oxide-promoted seed germination in Arabidopsis. Plant Cell. 2024:36(2):367–382. 10.1093/plcell/koad 27637930821 PMC 10827312 · doi ↗ · pubmed ↗