In darkness, remember the light: Chlamydomonas retains low- and high-light-induced acclimatory phenotypes in the dark
Guy Levin

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
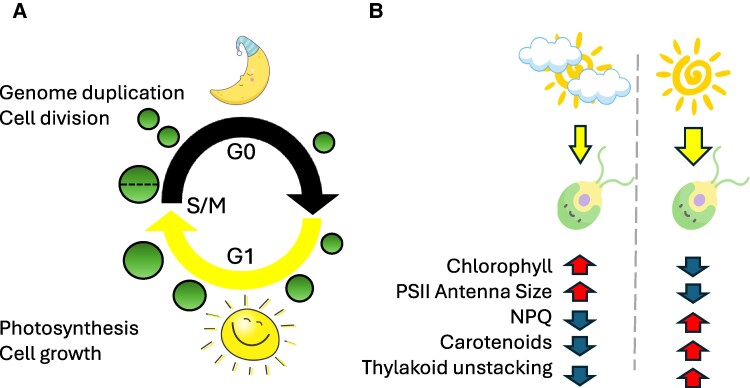 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhotoreceptor and optogenetics research · Circadian rhythm and melatonin · Advanced Thermodynamics and Statistical Mechanics
Plants encounter a wide variety of light conditions throughout their life cycle. For example, during winter, they may experience long periods of limited light, while in summer, light may be in excess. Light availability also changes during the day, where light is limited at dawn and dusk, while plentiful during midday. Photoautotrophs' metabolism is tightly synchronized with diurnal light via multiple mechanisms, including circadian regulation and responses to external cues (Oravec and Greenham 2022; De Barros Dantas et al. 2023). In the green alga Chlamydomonas (C. reinhardtii), cell growth occurs during the day when light is available, while genome replication and cell division occur at the beginning of the night (Figure) (Umen and Liu 2023). Low light can lead to retarded growth due to energy limitation, but excessive light may cause a similar effect due to enhanced reactive oxygen species formation (Erickson et al. 2015). Photosynthetic organisms can respond to such changes in light availability by increasing light-harvesting under limited light and both limiting light absorption and enhancing energy dissipation under excess light (Figure) (Croce 2020). To date, it is unclear if the diurnal programming of photosynthetic metabolism in Chlamydomonas plays a role in protecting the photosynthetic machinery from excess light or if exposure to suboptimal light intensities affects the diurnal program.
In new work, Sunnyjoy Dupuis, Valle Ojeda, and colleagues (Dupuis et al. 2025) use a systems approach to analyze the effect of diurnal light intensity on Chlamydomonas cells synchronized with 12-hour-light/-dark (day/night) and temperature cycles. The growth of cells under both limiting low light (LL) or excessive high light (HL) was retarded compared to cells grown under optimal, medium light (ML). However, light intensity had little impact on the Chlamydomonas cell cycle, which maintained synchronized cell division upon transition to the dark. Similarly, LL-, ML-, and HL-grown cells showed similar diurnal rhythmic gene expression programs—that is, gene and protein expression peaks simultaneously in cells grown under optimal or suboptimal light. Nevertheless, the expression of many specific genes and proteins was altered in LL- and HL-grown cells compared to ML, including enhanced accumulation of photoprotective proteins in the latter.
Interestingly, the variation in expression was not limited to the light phase but persisted after 10 h of darkness, showing that the daytime light intensity affects gene and protein expression at night. Unstacking of thylakoid membranes in HL-grown cells, which allows for efficient repair of photodamaged photosynthetic machinery, was also retained during the dark phase. The capacity to maintain high levels of nonphotochemical quenching (NPQ), where excess light is harmlessly dissipated as heat via proteins and carotenoids (Erickson et al. 2015), was also maintained in HL-grown cells during the night, with evidence suggesting lutein is the key quencher of the fastest NPQ component, q_E_. Unlike q_E_, other components of NPQ, including state transitions (q_T_) and Zeaxanthin-dependent quenching (q_Z_), were mostly active during the day but less prevalent at night. During q_T_, subunits of PSII antenna, the light-harvesting complex II (LHCII), are transferred to PSI to reduce excitation pressure from PSII. In q_Z_, zeaxanthin accumulates and provides a pathway for dissipation of excess energy (Erickson et al. 2015).
Maximal PSII efficiency was lower in HL-grown cells at the beginning of the light phase but recovered after 10 h, suggesting that Chlamydomonas adjusts and repairs its photosynthetic machinery after long exposure to diurnal HL. Higher chlorophyll a/b ratios in HL-grown cells indicated a reduction of the PSII antenna size, as chlorophyll b mainly resides in LHCII. A smaller antenna size (fewer LHCII subunits and chlorophylls) protects photosynthesis by reducing light absorption under HL conditions and was recently suggested to be a key photoprotective mechanism in the HL-tolerant alga Chlorella ohadii (Levin et al. 2024). The authors also noted the accumulation of the LHCII subunit LHCBM9 in HL-grown cells after 10 h of light exposure. LHCBM9 was shown to enhance the stability and energy dissipation of PSII (Grewe et al. 2014) and may be key to the recovered PSII efficiency observed after 10 h of light exposure.
This work shows that the Chlamydomonas diurnal program is robust and maintained under challenging light conditions whether limiting or in excess. Moreover, Chlamydomonas seems to retain HL-dependent acclimatory phenotypes during the night, as indicated by unstacked thylakoid membranes, reduced chlorophyll and photosynthetic protein abundance, and elevated levels of lutein, NPQ capacity, and the photoprotective q_E_-inducing protein LhcSR3 (Figure). These findings suggest that Chlamydomonas “remembers” the light intensity it encountered during the day, perhaps as a mechanism to prepare for the light levels that are expected on the following day. This complex mechanism, coordinated by a crosstalk between multiple regulatory processes, including circadian regulation and light and temperature sensing, would be beneficial for optimizing light harvesting over long periods, such as seasonal changes. In future work, it would be interesting to investigate the individual roles of each component of this mechanism, which are poorly understood in unicellular phototrophs.
Recent related articles in The Plant Cell
(Ostermeier et al. 2024) reviewed the latest knowledge of the structure, biogenesis, and evolution of thylakoid membranes, including their repair post-light-induced damage.(Romero-Losada et al. 2025) used a multiomics approach to determine the effect of different daylight durations on the green alga Ostreococcus tauri.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Croce R . Beyond ‘seeing is believing’: the antenna size of the photosystems in vivo. New Phytol. 2020:228(4):1214–1218. 10.1111/nph.1675832562266 PMC 7689736 · doi ↗ · pubmed ↗
- 2De Barros Dantas LL, Eldridge BM, Dorling J, Dekeya R, Lynch DA, Dodd AN. Circadian regulation of metabolism across photosynthetic organisms. Plant J. 2023:116(3):650–668. 10.1111/tpj.1640537531328 PMC 10953457 · doi ↗ · pubmed ↗
- 3Dupuis S, Ojeda V, Gallaher SD, Purvine SO, Glaesener AG, Ponce R, Nicora CD, Bloodsworth K, Lipton MS, Niyogi KK, et al Too dim, too bright, and just right: systems analysis of the Chlamydomonas diurnal program upon acclimation to limiting and excess light. Plant Cell. 2025; 10.1093/plcell/koaf 086PMC 1213697340251989 · doi ↗ · pubmed ↗
- 4Erickson E, Wakao S, Niyogi KK. Light stress and photoprotection in Chlamydomonas reinhardtii. Plant J. 2015:82(3):449–465. 10.1111/tpj.1282525758978 · doi ↗ · pubmed ↗
- 5Grewe S, Ballottari M, Alcocer M, D’Andrea C, Blifernez-Klassen O, Hankamer B, Mussgnug JH, Bassi R, Kruse O. Light-harvesting complex protein LHCBM 9 is critical for photosystem II activity and hydrogen production in Chlamydomonas reinhardtii. Plant Cell. 2014:26(4):1598–1611. 10.1105/tpc.114.12419824706511 PMC 4036574 · doi ↗ · pubmed ↗
- 6Levin G, Yasmin M, Liran O, Hanna R, Kleifeld O, Horev G, Wollman F-A, Schuster G, Nawrocki WJ. Processes independent of nonphotochemical quenching protect a high-light-tolerant desert alga from oxidative stress. Plant Physiol. 2024:197(1):kiae 608. 10.1093/plphys/kiae 60839520699 PMC 11663709 · doi ↗ · pubmed ↗
- 7Oravec MW, Greenham K. The adaptive nature of the plant circadian clock in natural environments. Plant Physiol. 2022:190(2):968–980. 10.1093/plphys/kiac 33735894658 PMC 9516730 · doi ↗ · pubmed ↗
- 8Ostermeier M, Garibay-Hernández A, Holzer VJC, Schroda M, Nickelsen J. Structure, biogenesis, and evolution of thylakoid membranes. Plant Cell. 2024:36(10):4014–4035. 10.1093/plcell/koae 10238567528 PMC 11448915 · doi ↗ · pubmed ↗