Genome sequences of Acinetobacter phage At2 and its host Acinetobacter tandoii W4-4-4
Soon Keong Wee, Biaoguo Yan, Su'Aidah Binte Mustaffa, Eric Peng Huat Yap

TL;DR
This paper reports the genome sequences of a phage and its bacterial host found in a Singapore mangrove, including resistance genes.
Contribution
The study provides new genome sequences of a phage and its Acinetobacter host with antimicrobial resistance features.
Findings
The genome sequences of Acinetobacter phage At2 and its host were determined.
Antimicrobial resistance genes were identified in the host genome.
Abstract
We present the genome sequences of Acinetobacter phage At2 and its host Acinetobacter tandoii strain W4-4-4, both isolated from a tropical mangrove swamp in Singapore. This report highlights the genetic features of both organisms, including antimicrobial resistance genes.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
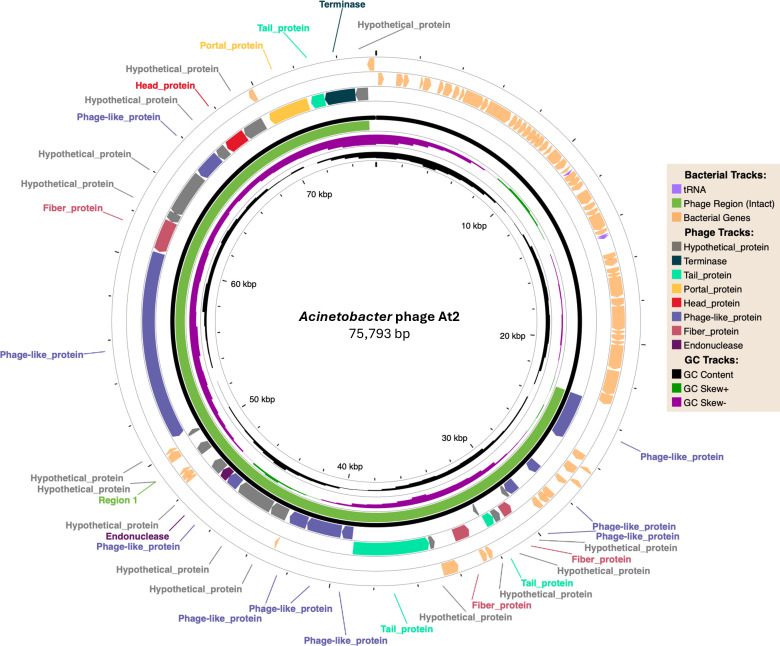 Fig 1
Fig 1| Class | Phenotype |
|---|---|
| Aminoglycoside | S: amikacin and gentamicin |
| Beta-lactam | S: ampicillin/sulbactam, piperacillin/tazobactam, doripenem, imipenem, meropenem, ceftazidime, and cefotaxime |
| Quinolone | S: ciprofloxacin |
| Sulfonamide | S: trimethoprim/sulfamethoxazole |
| Tetracycline | S: tetracycline |
| Trimethoprim | S: trimethoprim/sulfamethoxazole |
- —Ministry of Education - Singaporehttp://dx.doi.org/10.13039/501100001459
- —Lee Kong Chian School of Medicine, Nanyang Technological Universityhttp://dx.doi.org/10.13039/501100011738
- —Nanyang Technological Universityhttp://dx.doi.org/10.13039/501100001475
- —Nanyang Technological Universityhttp://dx.doi.org/10.13039/501100001475
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Genomics and Phylogenetic Studies · Antibiotic Resistance in Bacteria
ANNOUNCEMENT
First identified from activated sludge in Australia (1), Acinetobacter tandoii has also been found in termite gut microbiota and exhibits phenol degradation properties (2). Bacteriophages coexist with their host bacteria in the natural environment (3, 4), driving evolution and potentially enhancing its bioremediation applications, such as phenol degradation. We report the draft genomes of Acinetobacter tandoii strain W4-4-4 and lytic Acinetobacter phage At2 co-isolated from brackish water at an urban coastal swamp in Singapore.
The bacterial host was isolated through sequential culture in Baumann enrichment medium (5), Leeds acinetobacter medium agar (6), and lysogeny broth agar at 37°C incubation. The phage was subsequently isolated from the same sample by plaque formation on the host monoculture using double-layer agar with 0.6% soft agar. Antibiotic susceptibility was assessed by disk diffusion following CLSI standards (7). DNA from the bacterial colony and phage lysate was extracted using QIAGEN DNA Mini Kit. Concentrations were measured by Nanodrop and Qubit HS dsDNA Assay. Genomes were sequenced using Nextera XT DNA library preparation on the Illumina HiSeq platform with 150 bp paired-end reads, yielding 5,293,391 host reads and 773,186 phage reads. Quality check and adaptor trimming were performed using FastQC v0.72 and Trimmomatic v0.36.5 (8, 9). De novo assembly was performed with Unicycler Galaxy version 0.4.6.0 (10, 11). Host species identification was done using dDDH analysis in TYGS Server (12). Host and phage genomes were annotated using NCBI Prokaryotic Genome Annotation Pipeline 4.9 (13) and Prokka 1.13 (14), respectively. Antimicrobial resistance genes, virulence factors, CRISPR arrays, and phage regions were predicted using ResFinder 3.2 (15), VFDB VFanalyzer (16), CRISPRCasFinder (17), and PHASTEST (18). Default parameters were used for all software tools.
Acinetobacter tandoii strain W4-4-4 has a 3,575,445 bp genome (GC content, 40.15%). The draft assembly, in 50 contigs (coverage depth, 373×; N50, 312,393 bp), contains 3,276 coding DNA sequence (CDS) genes, 60 pseudogenes, 66 tRNAs, and 3 rRNAs. It carries the blaOXA gene, sharing 92.63% nucleotide identity with blaOXA-664, an intrinsic carbapenemase in Acinetobacter tandoii (19). Phenotypically, it is susceptible to various antibiotics including carbapenems (Table 1). No CRISPR arrays were detected, but two intact prophage regions (34.5 kb to 38 CDS and 39.1 kb to 43 CDS) and one questionable prophage region were identified.
Acinetobacter phage At2 has a 75,793 bp circular genome (GC content, 39.47%; coverage depth, 3,131×) (Fig. 1). It encodes 99 CDS and 2 tRNAs, with 87 (87.8%) being hypothetical proteins. It features two N4-like RNA polymerases (virion-associated RNA polymerase and RNA polymerase II) and a single-stranded DNA-binding protein, suggesting a genome injection mechanism akin to phage N4 (20). Other replication-associated genes encode DNA polymerase I, DNA helicase, DNA primase, ATPase, ribonucleotide reductase, and thymidylate synthetase. Structural proteins identified include baseplate hub, tail fibers, and tail terminase components. Homology with N4 proteins suggests a conserved N4-like genome injection and replication mechanism (21). Classified within the Caudoviricetes class, Phage At2 shares 67% and 70% amino acid identity with two Acinetobacter phages from the Schitoviridae family infecting Acinetobacter beijerinckii (phage nACB1) and Acinetobacter pittii (phage VB_ApiP_XC38), respectively.
Gene annotation plot of Acinetobacter phage At2 generated using PHASTEST (18).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Carr EL, Kämpfer P, Patel BKC, Gürtler V, Seviour RJ. 2003. Seven novel species of Acinetobacter isolated from activated sludge. Int J Syst Evol Microbiol 53:953–963. doi:10.1099/ijs.0.02486-012892111 · doi ↗ · pubmed ↗
- 2Van Dexter S, Boopathy R. 2019. Biodegradation of phenol by Acinetobacter tandoii isolated from the gut of the termite. Environ Sci Pollut Res Int 26:34067–34072. doi:10.1007/s 11356-018-3292-430264343 · doi ↗ · pubmed ↗
- 3Koskella B, Brockhurst MA. 2014. Bacteria-phage coevolution as a driver of ecological and evolutionary processes in microbial communities. FEMS Microbiol Rev 38:916–931. doi:10.1111/1574-6976.1207224617569 PMC 4257071 · doi ↗ · pubmed ↗
- 4Simmons EL, Drescher K, Nadell CD, Bucci V. 2018. Phage mobility is a core determinant of phage-bacteria coexistence in biofilms. ISME J 12:531–543. doi:10.1038/ismej.2017.19029125597 PMC 5776469 · doi ↗ · pubmed ↗
- 5Baumann P. 1968. Isolation of Acinetobacter from soil and water. J Bacteriol 96:39–42. doi:10.1128/jb.96.1.39-42.19684874313 PMC 252249 · doi ↗ · pubmed ↗
- 6Jawad A, Hawkey PM, Heritage J, Snelling AM. 1994. Description of leeds Acinetobacter medium, a new selective and differential medium for isolation of clinically important Acinetobacter spp., and comparison with Herellea agar and Holton’s agar. J Clin Microbiol 32:2353–2358. doi:10.1128/jcm.32.10.2353-2358.19947814465 PMC 264065 · doi ↗ · pubmed ↗
- 7CLSI. 2018. Performance standards for antimicrobial susceptibility testing. 29th ed. CLSI supplement M 100. Clinical and Laboratory Standards Institute, Wayne, PA.
- 8Andrews S. 2010. Fast QC a quality control tool for high throughput sequence data. Available from: https://www.bioinformatics.babraham.ac.uk/projects/fastqc