In vitro untargeted polar metabolomics data from B. cenocepacia and S. aureus biofilm supernatants
Hayden Skaggs, Deborah R. Yoder-Himes

TL;DR
This paper provides a dataset of 618 polar metabolites found in biofilm supernatants from two human pathogens.
Contribution
The study presents an untargeted polar metabolomics dataset from B. cenocepacia and S. aureus biofilms.
Findings
618 polar metabolites were identified using HPLC and HRMS.
The dataset includes metabolites from in vitro biofilm supernatants of two human pathogens.
Abstract
Here, we offer a metabolomics dataset generated via high-performance liquid chromatography and high-resolution mass spectrometry analysis of in vitro biofilm supernatant harvested from the human pathogens Burkholderia cenocepacia and Staphylococcus aureus. A total of 618 polar metabolites were identified across all experimental groups.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
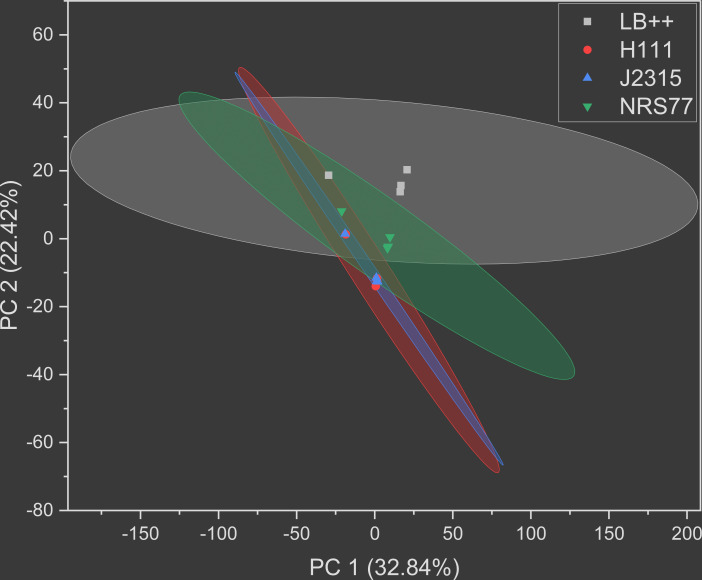 Fig 1
Fig 1- —U.S. Cystic Fibrosis Foundation
- —University of Louisvillehttp://dx.doi.org/10.13039/100007924
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial biofilms and quorum sensing · Cystic Fibrosis Research Advances · Vibrio bacteria research studies
ANNOUNCEMENT
Staphylococcus aureus and Burkholderia cenocepacia are well-studied pathogens (1–4) that can colonize individuals who are immunocompromised or with the genetic condition cystic fibrosis (CF) (5). The polymicrobial nature of the CF lung makes it difficult to obtain distinct in vivo metabolic profiles for specific pathogens (6–8). Further, freely accessible metabolomics data sets for bacteria, particularly in non-planktonic conditions, remain paltry compared with eukaryotes. This study aims to identify the extracellular metabolites produced by these pathogens under biofilm conditions in order to better understand the complex metabolome of the CF lung.
Collection of cell-free supernatants from epidemic isolates of B. cenocepacia, H111 and J2315, and S. aureus NRS77 from quadruplicate 7-day-old mono-culture biofilms was done as previously described (9). The sterile supernatant from each of the three cultures and the uninoculated medium (LB Lennox broth + 150 mM MOPS + 1% glucose) was lyophilized using a LabConco FreeZone 2.5L benchtop freeze dryer for 24 h. Dried supernatants were reconstituted in 2 mL 50% acetonitrile and injected into a Thermo DIONEX UltiMate 3000 HPLC system, then analyzed using a Thermo Q Exactive HF Hybrid Quadrupole-Orbitrap Mass Spectrometer. The LC system was equipped with a reversed phase column (Waters Acquity UPLC HSS T3 column, 2.1 × 150 mm, 1.8 µm) and hydrophilic interaction chromatography column (Millipore SeQuant ZIC-cHILIC column, 2.1 × 150 mm, 3 µm). Information regarding specific column conditions and solvent gradient can be found in our related work (6). To identify metabolites, 2D LC-MS/MS data were first matched to our own proprietary database that contains parent ion m/z, MS/MS spectra, and retention time of 363 authentic standards. Thresholds were set as spectral similarity ≥0.4, retention time difference ≤0.15, and m/z variation window ≤5 ppm. 2D LC-MS/MS data without a match with the metabolites in the proprietary database were further analyzed using Compound Discoverer software (v 3.1, Thermo Fisher Scientific, Germany) as described in (9). A total of 618 polar metabolites were identified between the three culture supernatants and the uninoculated culture media. Principal component analysis (PCA) was performed using Origin(pro) graphing software v2024b. For all software listed above, default parameters were used unless otherwise noted.
Principal component analysis (PCA) was performed on the raw chromatographic peak intensities for each of the metabolomic data sets. The PCA scores plot depicts strong similarity between the metabolic profiles of the B. cenocepacia clinical isolates in the form of overlapping 95% confidence interval (CI) ellipticals (Fig. 1). The S. aureus laboratory strain exhibits a slight similarity in its metabolic profile with the Burkholderia, but is otherwise independent. The control condition was as expected.
Principal component analysis (PCA) scores plot of biofilm supernatant metabolic profiles. Principal component analysis was performed on the raw chromatographic peak intensities of each metabolite identified across all experimental conditions. One data point represents a single biological replicate. LB++ (gray), B. cenocepacia H111 (red), B. cenocepacia J2315 (blue), and S. aureus NRS77 (green). Sterile supernatant of uninoculated LB + 1% glucose + 150 mM MOPS (LB++) serves as the control condition, with all further samples being characterized in this same media. The axes represent the percentage of variance for that respective principal component. The colored ellipses represent the 95% CI of PCA scores. Samples with similar scores will appear close to each other.
Overall, the untargeted polar metabolomics data set provides a basis for understanding the extracellular biofilm metabolome of these pathogens. Identifying the metabolic secretions of these organisms is paramount to understanding their pathogenicity within the CF lung. These data could be used to explore differences between in vitro and in vivo CF metabolomes, other microbial interactions in the CF lung, interactions between these pathogens and the human host immune systems, and in a broader sense, other polymicrobial infections.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Li PUMA JJ, Spilker T, Gill LH, Campbell PW III, Liu L, Mahenthiralingam E. 2001. Disproportionate distribution of Burkholderia cepacia complex species and transmissibility markers in cystic fibrosis . Am J Respir Crit Care Med 164:92–96. doi:10.1164/ajrccm.164.1.201115311435245 · doi ↗ · pubmed ↗
- 2Ahn Y, Kim JM, Kweon O, Kim S-J, Jones RC, Woodling K, Gamboa da Costa G, Li Puma JJ, Hussong D, Marasa BS, Cerniglia CE. 2016. Intrinsic resistance of Burkholderia cepacia complex to benzalkonium chloride. M Bio 7:e 01716-16. doi:10.1128/m Bio.01716-1627879334 PMC 5120141 · doi ↗ · pubmed ↗
- 3Bal AM, Gould IM. 2005. Antibiotic resistance in Staphylococcus aureus and its relevance in therapy. Expert Opin Pharmacother 6:2257–2269. doi:10.1517/14656566.6.13.225716218886 · doi ↗ · pubmed ↗
- 4Monaco M, Pimentel de Araujo F, Cruciani M, Coccia EM, Pantosti A. 2017. Worldwide epidemiology and antibiotic resistance of Staphylococcus aureus. Curr Top Microbiol Immunol 409:21–56. doi:10.1007/82_2016_327025380 · doi ↗ · pubmed ↗
- 5Lipuma JJ. 2010. The changing microbial epidemiology in cystic fibrosis. Clin Microbiol Rev 23:299–323. doi:10.1128/CMR.00068-0920375354 PMC 2863368 · doi ↗ · pubmed ↗
- 6Twomey KB, Alston M, An SQ, O’Connell OJ, Mc Carthy Y, Swarbreck D, Febrer M, Dow JM, Plant BJ, Ryan RP. 2013. Microbiota and metabolite profiling reveal specific alterations in bacterial community structure and environment in the cystic fibrosis airway during exacerbation. P Lo S One 8:e 82432. doi:10.1371/journal.pone.008243224358183 PMC 3866110 · doi ↗ · pubmed ↗
- 7Jørgensen KM, Wassermann T, Johansen HK, Christiansen LE, Molin S, Høiby N, Ciofu O. 2015. Diversity of metabolic profiles of cystic fibrosis Pseudomonas aeruginosa during the early stages of lung infection. Microbiology (Reading) 161:1447–1462. doi:10.1099/mic.0.00009325873584 · doi ↗ · pubmed ↗
- 8Muhlebach MS, Sha W. 2015. Lessons learned from metabolomics in cystic fibrosis. Mol Cell Pediatr 2:9. doi:10.1186/s 40348-015-0020-826542299 PMC 4883209 · doi ↗ · pubmed ↗