Plasmid sequences and availability of a two-plasmid system for CRISPRi knockdown of Clostridioides difficile genes without antibiotic selection
Michelle Chua, Daniel Erickson, James Collins

TL;DR
A new CRISPRi system for Clostridioides difficile was developed that doesn't need antibiotics to maintain gene knockdown.
Contribution
A two-plasmid CRISPRi system for C. difficile was created without requiring antibiotic selection.
Findings
The system uses an optimized tetracycline-inducible dCas9 for gene knockdown.
A toxin-antitoxin system ensures plasmid stability without antibiotics.
Successful knockdown of the essential gene walA was demonstrated.
Abstract
A two-plasmid CRISPRi system for Clostridioides difficile that does not require antibiotic maintenance was developed. pJAK184.tetR.PT5-3.dCas9 contains an optimized tetracycline-inducible dCas9 for chromosomal insertion. pJC.15A.sgRNA.TA encodes a toxin-antitoxin system for stable maintenance, and mCherry, which is exchangeable for a customized sgRNA. We demonstrate the knockdown of the essential gene walA.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
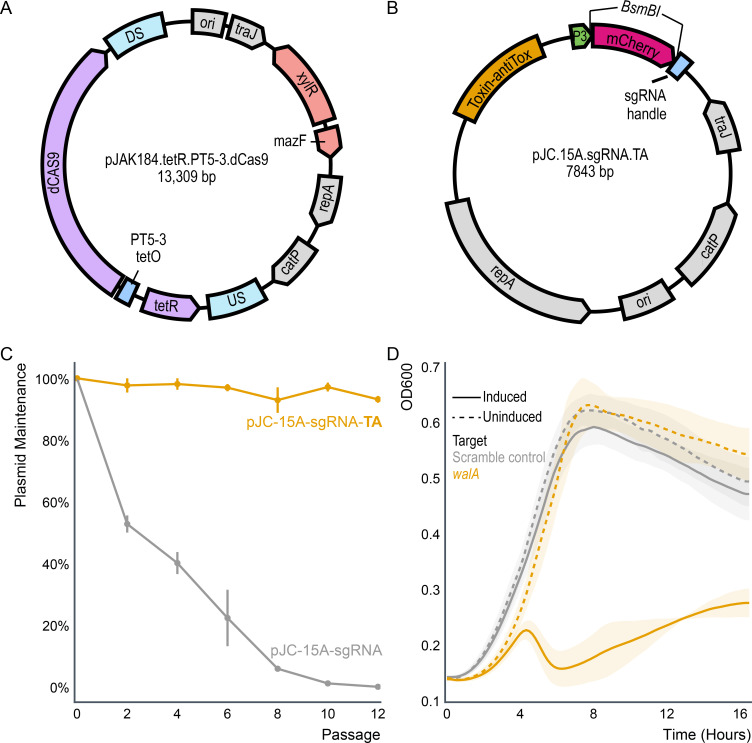 Fig 1
Fig 1| Strains/plasmids/primers | Relevant features | Source |
|---|---|---|
| pJAK184.tetR.PT5-3.dCas9 | This work | |
| pJAK184 | Xylose-inducible | ( |
| pRPF185 | ( | |
| pIA33 | dCas9 gene | ( |
| pJC.15A.sgRNA.TA | This work | |
| pPEPZ-sgRNAclone | mCherry gene | ( |
| pGEN222 | p15A ori | ( |
| pRPF185 | ( | |
| Toxin-antitoxin system | ||
| oMC396 |
| |
| oMC397 |
| |
| oJC842 |
| |
| oJC714 |
| |
| oJC850 |
| |
| oJC845 |
| |
| oJC846 |
| |
| oMC221 |
| |
| oJC715 |
| |
| oJC851 |
| |
| oMC207 |
| |
| oMC220 |
| |
- —National Institute of General Medical Scienceshttp://dx.doi.org/10.13039/100000057
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsClostridium difficile and Clostridium perfringens research · Viral gastroenteritis research and epidemiology · Bacteriophages and microbial interactions
ANNOUNCEMENT
Tools for the genetic manipulation of C. difficile are essential for understanding its pathogenesis. Here, we describe a CRISPRi system that does not require antibiotic maintenance, enabling the study of essential genes both in vitro and in vivo.
pJAK184.tetR.PT5-3.dCas9 contains catalytically inactive Cas9 (dCas9) under the control of an optimized tetracycline-inducible promoter (Fig. 1A). Homologous regions enable integration into the C. difficile chromosome downstream of the bacitracin permease gene, and a xylose-inducible mazF enables the selection of double crossovers, as described by Fuchs et al. (1). The plasmid pJC.15A.sgRNA.TA encodes mCherry flanked by BsmBI restriction sites directly upstream of the tracrRNA (Fig. 1B). The restriction sites enable the rapid replacement of mCherry with a CRISPR RNA target constructed by annealing oligo pairs with an overhang compatible with the backbone vector. To allow stable replication without the need for antibiotic selection, the plasmid contains a toxin-antitoxin (TA) system derived from a putative chromosomal TA system in C. difficile CD630, which is absent in the majority of C. difficile strains. The TA system enables stable plasmid maintenance without the need for antibiotics (Fig. 1C). This system replicates stably and can knock down essential genes without incurring fitness costs in the absence of induction (Fig. 1D).
CRISPRi system. (A) Map of pJAK184.tetR.PT5-3.dCas9. This plasmid contains dCas9 driven by an optimized promoter (PT5-3) and was designed to be integrated into the C. difficile genome using mazF as the counter-selectable marker. (B) Map of pJC.15A.sgRNA.TA. This plasmid contains mCherry, which can be removed to integrate the sgRNA of interest. Once transformed into C. difficile, pJC.15A.sgRNA.TA remains stable due to the presence of a toxin-antitoxin system. (C) The introduction of a putative C. difficile toxin-antitoxin system allows for stable plasmid maintenance without the need for antibiotics. Three biological replicates of each strain were passaged every 12 h in BHIS medium without selection and subsequently plated onto BHIS agar with and without antibiotics. (D) Growth in the TY medium was inhibited when walA was targeted via CRISPRi. Strains were grown in a medium with or without anhydrotetracycline (50 ng/mL) to induce dCas9.
The plasmids were constructed as follows: For Gibson assembly and in vivo recombination, DNA fragments were synthesized or amplified using primers containing 25 nucleotide homologous ends. The promoter sequence driving dCas9 (PT5-3) was derived from Sorg et al. (2) and synthesized by Twist Bioscience. The xylose-inducible mazF, origin of replication, repA, and catP genes were amplified from pJAK184 using primers FFO-362 and FFO-363 (1). The bacitracin permease upstream and downstream arms and tetR and dCas9 genes were assembled via Gibson assembly from their respective sources, and the entire region was amplified using primers oMC396 and oMC397.
The mCherry gene was amplified from the pPEPZ-sgRNAclone plasmid using primers oJC842 and oJC714. Primers oJC850 and oJC845 were used to amplify the p15A ori gene from pGEN222. C. difficile replication and traJ/catP genes were amplified from pRPF185 using primers oJC846/oMC221 and oJC715/oJC851, respectively. Primers oMC207 and oMC220 were used to amplify the toxin-antitoxin system from C. difficile CD630 genomic DNA. Fragments were assembled via in vivo recombination (3). Strains, plasmids, and primers are listed in Table 1.
Following assembly, pJAK184.tetR.PT5-3.dCas9 was conjugated into C. difficile R20921 and chromosomally inserted downstream of the bacitracin permease gene. Clean insertions were confirmed by Sanger sequencing. The plasmid pJC.15A.sgRNA.TA was digested with BsmBI, sgRNAs targeting either walA or a scrambled control were inserted, and the plasmids were conjugated into dCas9 containing C. difficile. Although the negative (scramble) control grew like wild-type, regardless of induction, the R20291 strain with the walA sgRNA plasmid displayed significantly reduced growth when induced but grew normally when uninduced (Fig. 1D). Growth studies were conducted in the absence of antibiotics.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Fuchs M, Lamm-Schmidt V, Sulzer J, Ponath F, Jenniches L, Kirk JA, Fagan RP, Barquist L, Vogel J, Faber F. 2021. An RNA-centric global view of Clostridioides difficile reveals broad activity of Hfq in a clinically important gram-positive bacterium. Proc Natl Acad Sci U S A 118:25. doi:10.1073/pnas.2103579118 PMC 823759534131082 · doi ↗ · pubmed ↗
- 2Sorg RA, Gallay C, Van Maele L, Sirard J-C, Veening J-W. 2020. Synthetic gene-regulatory networks in the opportunistic human pathogen Streptococcus pneumoniae Proc Natl Acad Sci U S A 117:27608–27619. doi:10.1073/pnas.192001511733087560 PMC 7959565 · doi ↗ · pubmed ↗
- 3Huang F, Spangler JR, Huang AY. 2017. In vivo cloning of up to 16 kb plasmids in E. coli is as simple as PCR. P Lo S One 12:e 0183974. doi:10.1371/journal.pone.018397428837659 PMC 5570364 · doi ↗ · pubmed ↗
- 4Fagan RP, Fairweather NF. 2011. Clostridium difficile has two parallel and essential Sec secretion systems. J Biol Chem 286:27483–27493. doi:10.1074/jbc.M 111.26388921659510 PMC 3149341 · doi ↗ · pubmed ↗
- 5Müh U, Pannullo AG, Weiss DS, Ellermeier CD. 2019. A Xylose-inducible expression system and a CRISPR interference plasmid for targeted knockdown of gene expression in Clostridioides difficile. J Bacteriol 201:14. doi:10.1128/JB.00711-18PMC 659739530745377 · doi ↗ · pubmed ↗
- 6Liu X, Kimmey JM, Matarazzo L, de Bakker V, Van Maele L, Sirard J-C, Nizet V, Veening J-W. 2021. Exploration of bacterial bottlenecks and Streptococcus pneumoniae pathogenesis by CRISP Ri-Seq. Cell Host Microbe 29:107–120. doi:10.1016/j.chom.2020.10.00133120116 PMC 7855995 · doi ↗ · pubmed ↗
- 7Galen JE, Nair J, Wang JY, Wasserman SS, Tanner MK, Sztein MB, Levine MM. 1999. Optimization of plasmid maintenance in the attenuated live vector vaccine strain Salmonella typhi CVD 908-htr A. Infect Immun 67:6424–6433. doi:10.1128/IAI.67.12.6424-6433.199910569759 PMC 97051 · doi ↗ · pubmed ↗