JNK signaling coordinates epithelial cell turnover through exocytosis in Drosophila ribosomal protein mutants
Nanami Akai, Yoshimasa Yagi, Tatsushi Igaki, Shizue Ohsawa

TL;DR
The study reveals how JNK signaling and exocytosis work together to control cell death and growth in fruit fly wing development.
Contribution
The discovery of a JNK-exocytosis feedback loop that coordinates cell turnover and Wg secretion in Drosophila Minute/+ mutants.
Findings
JNK signaling increases exocytosis and Wg secretion in dying cells during Drosophila wing development.
Elevated exocytosis promotes caspase activation through a feedback loop involving Dronc.
The JNK-exocytosis-Wg pathway is universally active downstream of JNK signaling, regardless of genetic background.
Abstract
Robust tissue growth is orchestrated by the precise coordination of cell death and cell proliferation. In the developing wing pouches of Drosophila Minute/+ animals, both cell death and compensatory cell proliferation are increased, thereby contributing to robust growth of mutant tissue. Here, we show that JNK-mediated elevation of exocytosis in dying cells is crucial for triggering cell turnover in M/+ wing morphogenesis. Mechanistically, elevated JNK signaling in dying cells upregulates exocytosis-related genes and Wingless (Wg), leading to enhanced Wg secretion. Notably, increased exocytosis promotes caspase activation via an exocytosis-Dronc amplification loop, sustaining apoptotic signaling while reinforcing Wg secretion through Dronc activation. Furthermore, this exocytosis-mediated Wg secretion and apoptotic feedback loop universally occurs downstream of JNK signaling, regardless…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
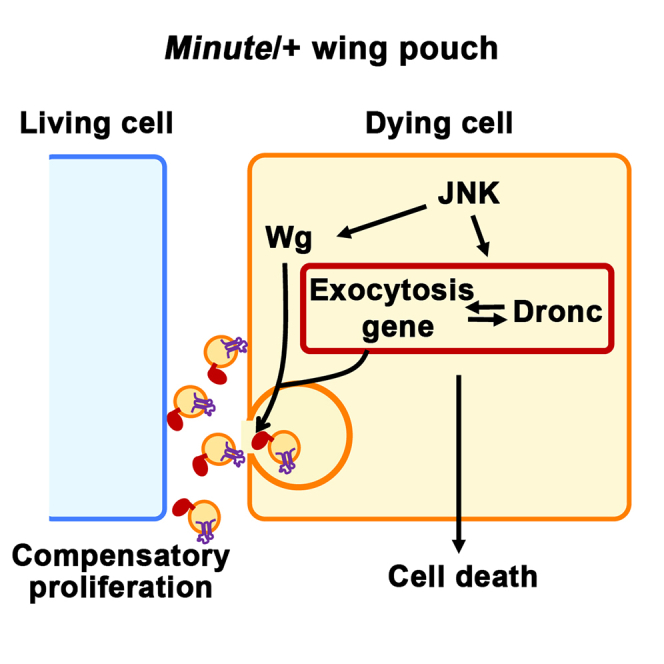 Figure 1
Figure 1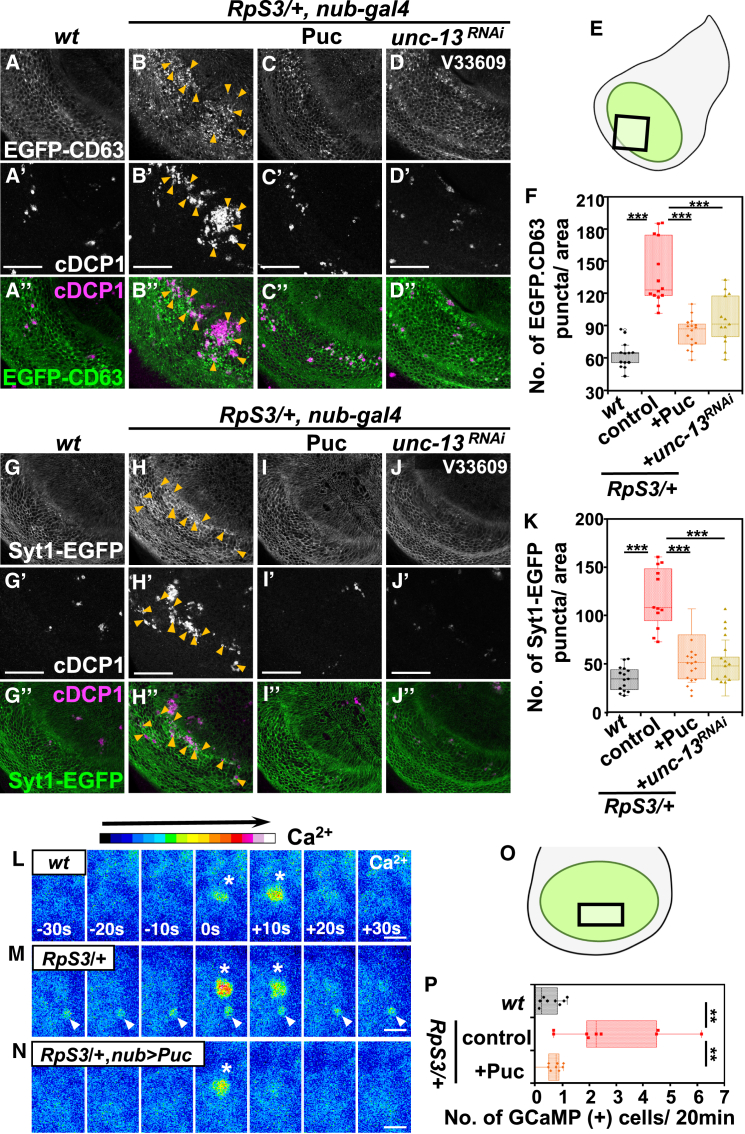 Figure 2
Figure 2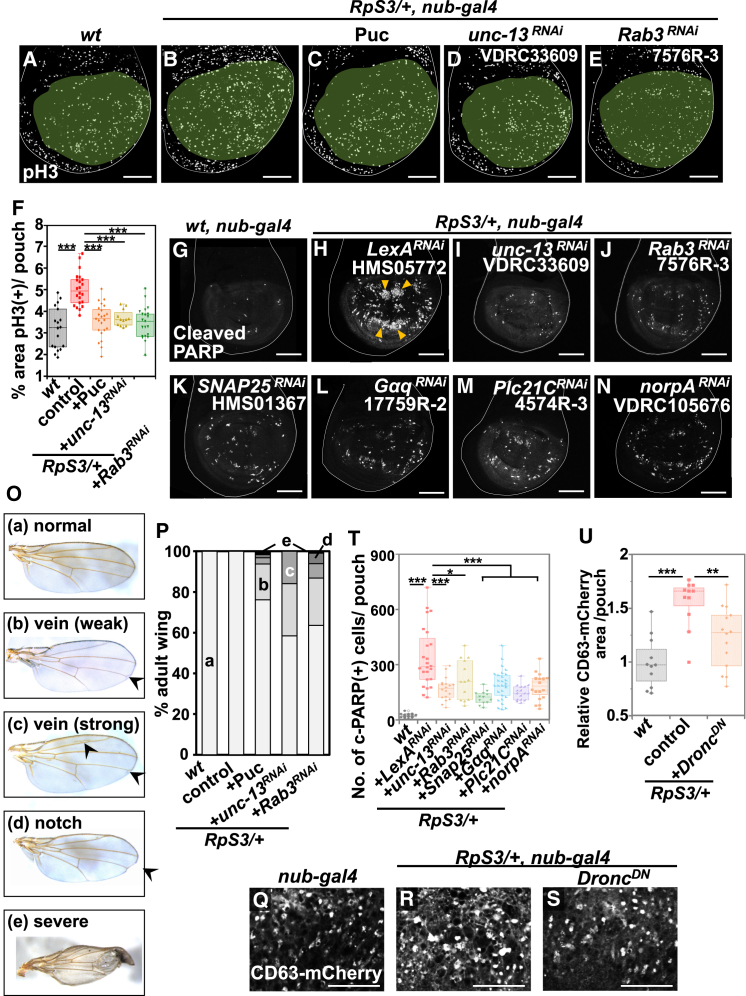 Figure 3
Figure 3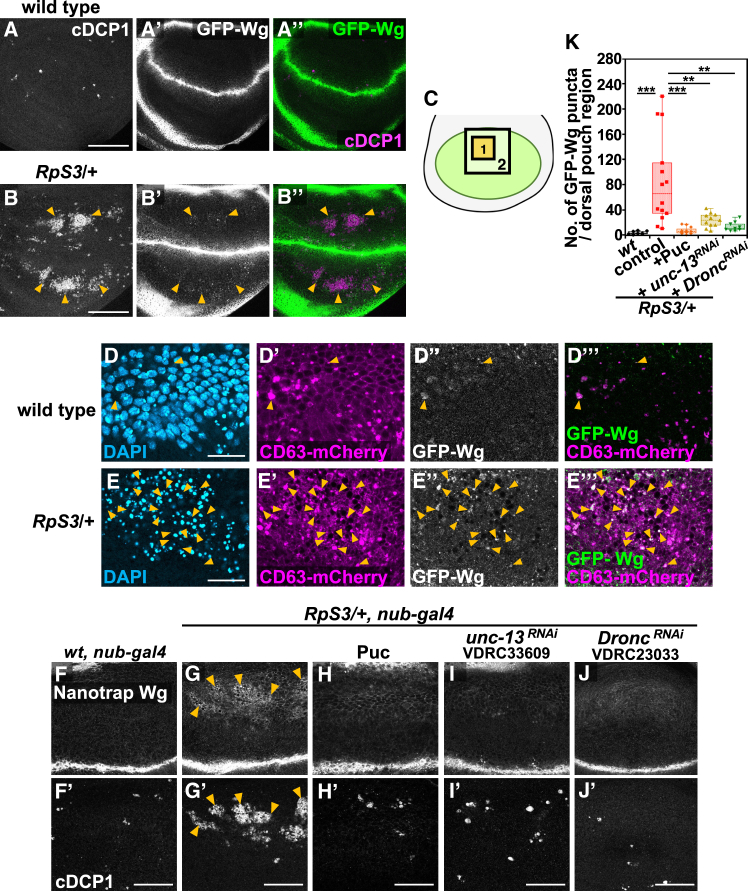 Figure 4
Figure 4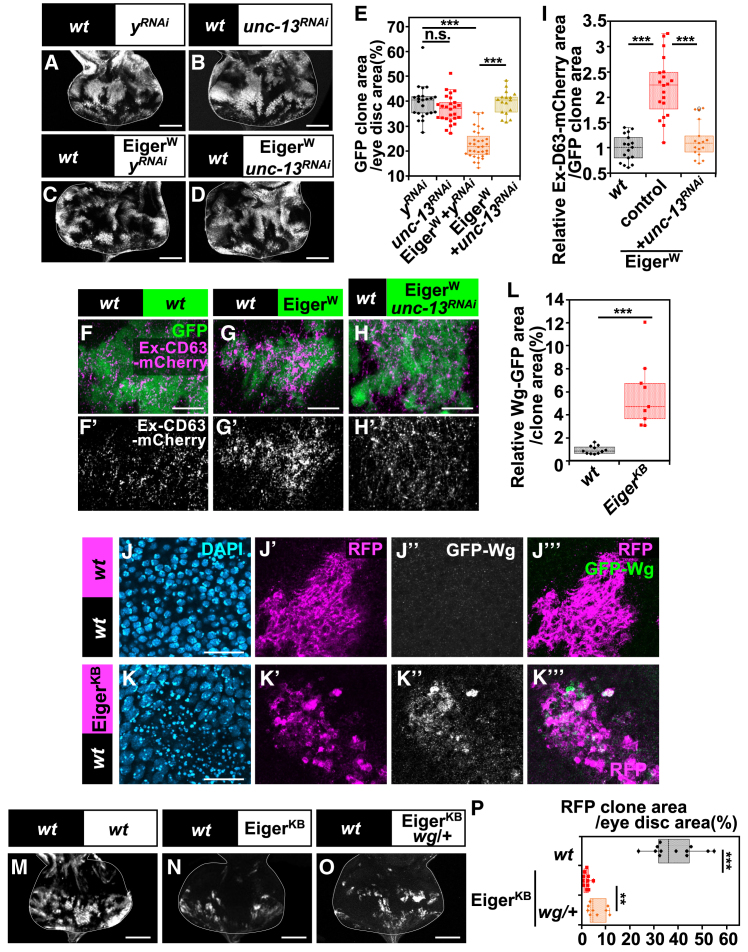 Figure 5
Figure 5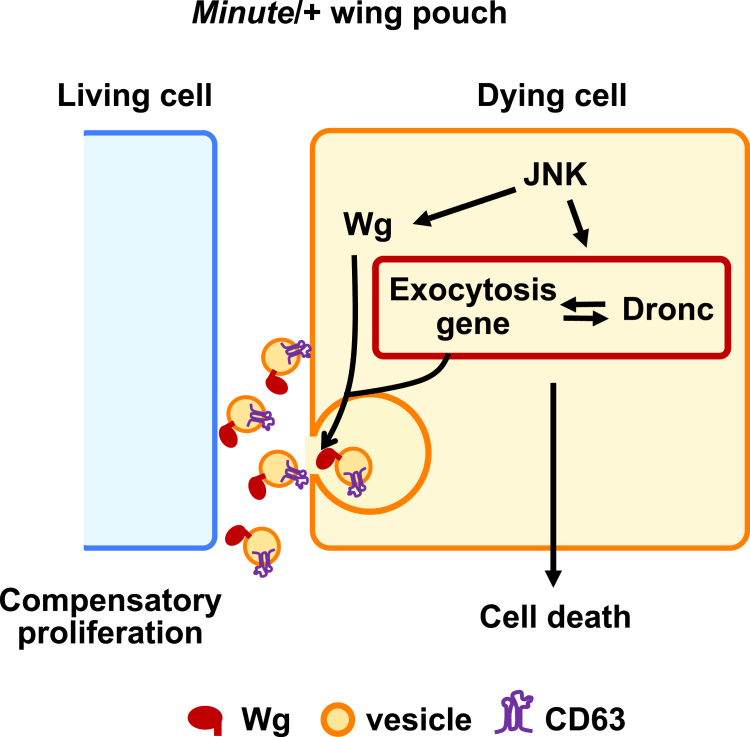 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCRISPR and Genetic Engineering · Invertebrate Immune Response Mechanisms · Cellular transport and secretion
Introduction
Ribosomes are essential molecular machines responsible for protein synthesis in living cells, and thus fundamental to life. In humans, heterozygosity of genes related to ribosomal proteins or ribosomal biogenesis factors leads to a group of genetic disorders collectively termed ribosomopathies. For instance, heterozygous mutations in various ribosomal protein genes, including RPS17,1 RPS26,2 RPS19,3 RPS24,4 and RPL27,5 are associated with Diamond-Blackfan anemia (DBA), with some patients exhibiting tissue-specific developmental anomalies, such as limb defects, cleft palate, and abnormal heart development.6^,^7 In addition, heterozygous mutations in the RpSA gene are linked to isolated congenital asplenia (ICA), a disorder characterized by the absence of a spleen at birth.8 However, the precise mechanisms by which a reduction in ribosomal protein gene dosage leads to these genetic disorders remain elusive.
In Drosophila, a series of heterozygous mutants for ribosomal protein genes, called Minute/+ (M/+), exhibit a pronounced developmental delay during larval stages.9 Despite this significant delay, M/+ animals are essentially normal flies without noticeable morphological defects, except for the thinner bristles, suggesting that M/+ animals exert certain mechanisms to overcome developmental perturbations caused by reduced ribosomal protein levels. It has been reported that extensive cell death,10 as well as cellular stress,10^,^11^,^12^,^13 is a distinctive characteristic of the M/+ wing pouch. Our previous study14 has shown that apoptotic cell death and the subsequent cell proliferation are dramatically increased in the M/+ wing pouch. Blocking this cell turnover by inhibiting cell death resulted in morphological abnormalities, indicating the essential role of cell turnover in M/+ wing morphogenesis. Genetic analyses have revealed that the induction of this cell turnover depends on activation of JNK (c-Jun N-terminal kinase) signaling. However, downstream events of JNK activation have remained to be elucidated.
The coordination of cell death and proliferation through cell-cell communications is crucial for proper development and homeostasis of multicellular organisms. Apoptotic cells, for example, can secrete mitogens such as Wingless (Wg; a Wnt homolog), dpp (a BMP homolog), and Hh, which could promote the proliferation of nearby cells in the Drosophila epithelium.15^,^16^,^17^,^18^,^19 Cell competition, a phenomenon in which cells with higher fitness (“winners”) eliminate neighboring less fit cells (“losers”) by inducing cell death,20 is another aspect of cell turnover. It has been reported that secretory factors from prospective loser cells contribute to cell competition. For instance, the Drosophila cytokine Unpaired 3 (Upd3), which is produced by losers, promotes cell competition between wild-type (winners) and M/+ cells (losers) in the epithelium.12 Furthermore, Madin-Darby canine kidney cells depleted of the polarity regulator gene scribble (scrib^KD^ MDCK cells) secrete fibroblast growth factor 21 (FGF21), which in turn promotes cell competition with neighboring wild-type MDCK cells.21 However, the mechanism by which these secreted factors are released from dying cells is still not fully understood.
Here, we found that exocytosis, acting as a downstream event of JNK signaling in dying cells, contributes to cell turnover in the M/+ wing pouch, which is essential for robust wing development in M/+ animals. Our genetic analyses suggest that increased exocytosis forms a signal amplification loop with Dronc, sustaining apoptotic signaling and reinforcing the release of Wg from dying cells, which in turn stimulates their own cell death and the proliferation of neighboring living cells. Furthermore, we found that this exocytosis-mediated Wg release is not restricted to M/+ context but appears to represent a universal event downstream of JNK signaling. Our findings provide new mechanistic insights into how dying cells coordinate robust development through cell-cell interactions.
Results
M/+ wing pouch elevates exocytosis downstream of JNK signaling
To elucidate the downstream event of JNK signaling in the M/+ wing pouch, we conducted RNA sequence (RNA-seq) analyses on GFP-labeled fluorescence-activated cell sorting (FACS)-sorted wing pouch cells from one of the M/+ mutants RpS3/+, compared to GFP-labeled wild-type control or RpS3/+ wing pouch cells overexpressing JNK inhibitor Puckered (Puc) (Figure S1A; the wing pouch is green-marked oval domain that becomes the adult wing blade). Among the 1,097 genes that were upregulated or downregulated in the RpS3/+ wing pouch relative to the wild-type control, we identified JNK targets or positive regulators including reaper22 and Gadd45,23 JAK/STAT targets including Socs36E24 and chinmo,25 Gr64 gustatory receptors and numerous genes associated with oxidative stress and DNA repair (Figure S1B; Tables S1 and S2), which aligns with previous reports on genes upregulated in the RpS3/+ or RpS17/+ wing disc.12^,^13 These data validate our experimental conditions for identifying genes required for JNK-mediated events in the M/+ wing pouch.
Mining the list of genes differentially expressed in the RpS3/+ wing pouch cells dependent on JNK signaling (Figure S1C; Table S3), we noticed that among the genes associated with the “secretion by cell” GO term (Figure S1B), the evolutionarily conserved exocytosis-related genes unc-13, SNAP25, and Cadps (calcium-dependent secretion activator) were upregulated in a JNK-dependent manner (2.29-fold increase, RpS3/+ compared to wild-type; 3.44-fold increase, RpS3/+ compared to RpS3/+ + Puc) (Figure S1D). Among these exocytosis-related genes, the upregulation of SNAP25 and unc-13 was consistent with the previous reports concerning genes upregulated in RpS3/+ or RpS17/+ cells.12^,^13
Calcium-dependent exocytosis is the process whereby cells release intracellular contents into the extracellular space. It has been shown that unc-13, SNAP25, and cadps collectively regulate the docking process of secretory vesicles to the plasma membrane during Ca^2+^-mediated exocytosis.26 Specifically, UNC-13, a conserved presynaptic protein with calcium-binding domains, interacts with syntaxin to prime vesicles for fusion, crucial for calcium-regulated exocytosis.27 SNAP-25, in conjunction with syntaxin-1 and synaptobrevin, forms the pivotal SNARE complex for neuronal exocytosis, assembling into a four-helix bundle that is essential for drawing vesicle and plasma membranes close together to enable membrane fusion.28^,^29 Like UNC-13, CAPS/Cadps possesses conserved C-terminal domains that are instrumental in the assembly of SNARE complexes, thus priming vesicles for Ca^2+^-induced exocytosis.30 First, to investigate whether exocytosis is upregulated in the M/+ wing pouch, we utilized the two exosome markers, CD63 tagged with EGFP and mCherry (EGFP-CD63 and CD63-mCherry).31^,^32^,^33^,^34^,^35^,^36^,^37 In addition, we utilized Syt1 tagged with EGFP (Syt1-EGFP), a widely used vesicle marker prevalent in both neuronal and non-neuronal cells.38^,^39^,^40 We observed a significant increase in EGFP-CD63-positive and Syt1-EGFP-positive vesicles in the RpS3/+ wing pouch compared to the wild-type control (Figures 1A, 1B, 1G, and 1H, corresponding to the boxed region in Figure 1E, quantified in Figures 1F and 1K). EGFP-CD63-positive vesicles were also found to be increased in the RpL19/+ wing pouch (Figure S1E, quantified in Figure S1F). Notably, these vesicles were predominantly observed in regions of the RpS3/+ wing pouch undergoing massive cell death (as indicated by arrowheads in Figures 1B, 1H, and S1E), a phenomenon we previously reported14 and also confirmed in this study (shown in Figure 2H, compared to Figure 2G, quantified in Figure 2T). Furthermore, we observed a significant increase in extracellular vesicles, including CD63-mCherry-positive and Syt1-EGFP-positive vesicles, in the RpS3/+ wing pouch compared to the wild-type control, as detected by staining with anti-mCherry or anti-GFP antibodies under non-permeabilizing conditions (Figures S1G, S1H, S3M, and S3N, quantified in Figure S1I and S3O).Figure 1M/+ wing pouch elevates exocytosis downstream of JNK signaling(A–D’’) The exosome marker EGFP-CD63 was expressed in the wing pouch of wild-type (A), RpS3/+ (B), RpS3/+, nub-Gal4, UAS-Puc (C), or RpS3/+, nub-Gal4, UAS-unc-13-RNAi (D) flies (white). The images correspond to the boxed area in the schematic diagram of the wing disc (E). Dying cells in the wing disc were visualized by anti-cleaved Dcp-1 staining (white). EGFP-CD63-positive vesicles were frequently observed in the area of morphological dying cells in the RpS3/+ wing pouch (Indicated by Orange arrowheads). Scale bar, 50 μm.(F) Boxplot with dots representing the number of EGFP-CD63-positive puncta in the pouch of genotypes shown in (A) (n = 13, number of wing pouches), (B) (n = 14), (C) (n = 16), and (D) (n = 13). Thick line, median; ∗∗∗, p < 0.001; Wilcoxon rank-sum test.(G–J’’) The vesicle marker Syt1-EGFP was expressed in the wing pouch of wild-type (G), RpS3/+ (H), RpS3/+, nub-Gal4, UAS-Puc (I), or RpS3/+, nub-Gal4, UAS-unc-13-RNAi (J) flies (white). The images correspond to the boxed area in the schematic diagram of the wing disc (E). Dying cells in the wing disc were visualized by anti-cleaved Dcp-1 staining (white). Syt1-EGFP-positive vesicles were frequently observed in the area of morphological dying cells in the RpS3/+ wing pouch (Indicated by Orange arrowheads). Scale bar, 50 μm.(K) Boxplot with dots representing the number of Syt1-EGFP-positive puncta in the wing pouch of genotypes shown in (G) (n = 15, number of wing pouches), (H) (n = 12), (I) (n = 14), and (J) (n = 16). Thick line, median; ∗∗∗p < 0.001; Wilcoxon rank-sum test.(L–N) Time-lapse imaging of Ca^2+^ signaling in cultured wing discs. The calcium reporter GCaMP6m (shown in pseudo color) was expressed in the wing pouch of wild-type (L), RpS3/+ (M), or RpS3/+, nub-Gal4, UAS-Puc (N) flies. Images were acquired at 10-s intervals. White arrowheads indicate the persistent activation of Ca^2+^ signaling. White asterisks indicate spontaneous Ca^2+^ flashes. The images correspond to the boxed area in the schematic diagram of the wing disc (O). Scale bar, 5 μm.(P) Quantification of cells exhibiting continuous Ca^2+^ activity for 20 min in the wing pouch. The respective genotypes are shown in (L) (n = 12), (M) (n = 10), and (N) (n = 9). These images are Z-stacked images from 3 confocal images (4 μm). Thick line, median; ∗∗p < 0.01; Wilcoxon rank-sum test.Figure 2. Exocytosis is required for massive cell turnover in the M/+ wing pouch(A–E) Wing discs of wild-type (A), RpS3/+ (B), RpS3/+, nub-Gal4, UAS-Puc (C), RpS3/+, nub-Gal4, UAS-unc-13-RNAi (D), or RpS3/+, nub-Gal4, UAS-Rab3-RNAi (E) flies were stained with anti-phospho-histone H3 (pH3) (Ser10) antibody (white). Wing pouches were marked in pale green. Scale bar, 100 μm.(F) Boxplot with individual dots representing pH3-positive areas in the pouch of genotypes shown in (A) (n = 18, number of wing pouches), (B) (n = 22), (C) (n = 23), (D) (n = 12), and (E) (n = 21). Thick line, median; ∗∗∗p < 0.001; Wilcoxon rank-sum test.(G–N) The activated-caspase-3 indicator CD8-PARP-Venus was expressed in the wing pouch of wild-type (G), RpS3/+, nub-Gal4, UAS-lexA-RNAi (H), RpS3/+, nub-Gal4, UAS-unc-13-RNAi (I), RpS3/+, nub-Gal4, UAS-Rab3-RNAi (J), RpS3/+, nub-Gal4, UAS-SNAP25-RNAi (K), RpS3/+, nub-Gal4, UAS- Gaq-RNAi (L), RpS3/+, nub-Gal4, UAS-Plc21C-RNAi (M), or RpS3/+, nub-Gal4, UAS-norpA-RNAi (N) flies, and dying cells in the wing pouch were visualized by anti-cleaved PARP staining (white). Orange arrowheads indicate massive cell death in the RpS3/+ wing pouch. Scale bar, 100 μm.(O) Adult wing phenotypes were classified as following five types: (a) normal, (b) vein (weak) (weakly bearing a brunched vein in the point of “landmark 14” (arrow)41 (c) vein (strong) (bearing other additional vein phenotypes (arrowheads) in addition to “landmark 14”), (d) notch, and (e) severe.(P) The rate of defective wings in the genotypes of wild-type (n = 55, number of adult wings), RpS3/+ (n = 44), RpS3/+, nub-Gal4, UAS-Puc (n = 63), RpS3/+, nub-Gal4, UAS-unc-13-RNAi (n = 101), and RpS3/+, nub-Gal4, UAS-Rab3-RNAi (n = 99).(Q–S) The exosome marker CD63-mCherry was expressed in the wing pouch of wild-type (Q), RpS3/+ (R), or RpS3/+, nub-Gal4, UAS-Dronc^DN^ (S) flies (white). The images correspond to the area enclosed by rectangle 1 in the schematic diagram of the wing disc (Figure 3C). Scale bar, 20 μm.(T) Boxplot with individual dots representing the number of cleaved-PARP-positive dying cells per pouch in genotypes shown in (G) (n = 18, number of wing pouches), (H) (n = 24), (I) (n = 18), (J) (n = 13), (K) (n = 13), (L) (n = 29), (M) (n = 16), and (N) (n = 19). Thick line, median; ∗∗p < 0.001, ∗p < 0.05; Wilcoxon rank-sum test.(U) Boxplot with dots representing the number of CD63-mCherry-positive puncta in the pouch of genotypes shown in (Q) (n = 11, number of wing pouches), (R) (n = 11), and (S) (n = 14). Thick line, median; ∗∗∗p < 0.001, ∗∗p < 0.01; Wilcoxon rank-sum test.
To further investigate the enhanced exocytosis in the RpS3/+ wing pouch, we utilized the GCaMP6m fluorescent calcium reporter42 to monitor Ca^2+^ activity, a known inducer of exocytosis. Persistent Ca^2+^ activity was observed in the area of massive cell death within the RpS3/+ wing pouch, along with spontaneous Ca^2+^ flashes similar to those present in wild-type control cells (Figures 1L and 1M, quantified in Figure 1P). Notably, these cells exhibiting persistent Ca^2+^ activity in the RpS3/+ wing pouch were SYTOX-positive, a dye that fluoresces upon binding DNA in cells with compromised plasma membranes, a characteristic of non-viable cells43 (Figure S1J). These observations suggest a potential link between persistent Ca^2+^ activity and an enhanced exocytic process in dying cells, potentially driven by elevated calcium levels in the RpS3/+ wing pouch.
We then examined whether exocytosis is the downstream event of JNK activation in the M/+ wing pouch. We found that blocking JNK signaling by overexpressing Puc significantly suppressed the increased number of EGFP-CD63/Syt1-EGFP-positive vesicles, as well as the emergence of cells exhibiting persistent Ca^2+^ signaling (Figure 1C, quantified in Figure 1F; Figure 1I, quantified in Figure 1K; Figure 1N, quantified in Figure 1P). Together, these observations suggest that exocytosis is elevated in dying cells within the M/+ wing pouch as a downstream event of JNK signaling.
Exocytosis is required for massive cell turnover in the M/+ wing pouch
We have previously demonstrated that JNK signaling is required for cell turnover in the M/+ wing pouch.14 Blocking JNK signaling by overexpressing Puc significantly reduced cell death and mitoses in the RpS3/+ wing pouch, as assayed by the CD8-PARP-Venus probe for caspase activity in the imaginal disc,44^,^45^,^46^,^47^,^48^,^49^,^50^,^51 and the M phase marker phospho-histone H3 (Figures 2A–2C, quantified in Figure 2F; Figures 2G, 2H, and S2A, quantified in Figure S2Q). In addition, we intriguingly found that downregulating unc-13, a docking factor involved in exocytosis, similarly diminished cell death and mitoses (Figures 2I and S2B, quantified in Figures 2T and S2Q, respectively; Figure 2D, quantified in Figure 2F), and also reduced EGFP-CD63-positive and Syt1-EGFP-positive vesicles in the RpS3/+ wing pouch (Figures 1D and 1J, quantified in Figures 1F and 1K, respectively). Knocking down rab3, a “Secretory GTPase Rabs” required for vesicle exocytosis, also significantly reduced cell death and mitoses in the RpS3/+ wing pouch (Figures 2J and S2C, quantified in Figures 2T and S2Q, respectively; Figure 2E, quantified in Figure 2F). Furthermore, overexpressing Puc or downregulating either unc-13 or rab3 led to morphological defects in the RpS3/+ adult wing, while their individual overexpression or downregulation did not affect wing development (Figures 2O, 2P, and S2R). These results suggest that exocytosis, as a downstream component of JNK signaling, is pivotal for cell turnover and subsequent normal morphogenesis in the M/+ wing pouch. Indeed, downregulating other docking factors such as SNAP25 or Syt1,52 another Secretory GTPase Rab27, or ALIX, an ESCRT (endosomal sorting complexes required for transport)-related protein involved in the biogenesis of extracellular vesicles,53^,^54 also significantly suppressed cell death in the RpS3/+ wing pouch (Figure 2K, quantified in Figure 2T; Figures S2D–S2J, quantified in Figure S2Q). Additionally, downregulating factors, such as Gαq, Plc21C, and norpA, which are required for intracellular Ca^2+^ release that triggers exocytosis process,39 also significantly suppressed cell death in the RpS3/+ wing pouch (Figures 2L–2N, quantified in Figure 2T; Figures S2K–S2P, quantified in Figure S2Q). Together, these data reinforce the vital role of exocytosis in cell turnover within the M/+ wing pouch, functioning as a downstream process of JNK signaling. Intriguingly, blocking Dronc activity by overexpressing Dronc^DN^ suppressed the elevation of exocytosis in the RpS3/+ wing pouch (Figure 2S, compared to Figure 2R, quantified in Figure 2U). This suggests the presence of a signal amplification loop between exocytosis and Dronc within the JNK signaling pathway, consistent with previous studies showing Dronc-JNK amplification loop that induce cell death.55^,^56
Dying cells secrete Wg via exocytosis in the M/+ wing pouch
We next sought to identify the protein secreted through exocytosis from dying cells in the M/+ wing pouch. We found that wg expression was specifically upregulated in dying cells within the RpS3/+ wing pouch, compared to the wild-type control (Figures S3A and S3B), as assayed by the wg-lacZ reporter.57 This elevated wg expression was significantly suppressed by overexpressing Puc in the RpS3/+ pouch (Figure S3C, quantified in Figure S3D), suggesting that wg expression is mediated through JNK signaling in this context, which is consistent with the previous studies showing that apoptotic cells produce Wg through JNK signaling.18^,^58^,^59 Additionally, our previous study14 suggested that cell turnover in the M/+ wing pouch is initiated by a mechanism akin to Wg-dependent cell competition, a phenomenon by which cells with higher Wg signaling activity eliminate neighboring cells with lower Wg signaling activity in the wing disc.60 Indeed, as demonstrated through previous genetic manipulations,14 and confirmed here, genetically reducing the Wg signaling gradient in the entire M/+ pouch significantly suppressed cell death (Figure S3F, quantified in Figure S3G). These observations led us to hypothesize that dying cells in the M/+ wing pouch might release Wg via JNK-mediated exocytosis, potentially conferring a survival advantage to their neighboring living cells. Supporting this hypothesis, it has been reported that Wnt/Wg can be released through extracellular vesicles including exosomes.33^,^61^,^62^,^63^,^64
Consistently, we found that GFP-Wg-positive puncta, derived from a knock-in allele,65 were more abundant in the RpS3/+ wing pouch compared to the wild-type control (Figures 3A and 3B). This increase in GFP-Wg-positive puncta, particularly in the area with massive cell death within the RpS3/+ wing pouch (Figures S3I–S3J’’’, compared to Figure S3H), was more evident when using a membrane-tethered anti-GFP nanobody (Vhh4-CD8), which immobilizes GFP-Wg on the cell surface65 (Figures 3F and 3G, quantified in Figure 3K). Notably, these GFP-Wg-positive puncta, observed both without and with the Vhh4-CD8 morphotrap, sometimes colocalized with CD63-mCherry-positive vesicles (Figures 3E and S3L, indicated by arrowheads), suggesting that GFP-Wg might be secreted via extracellular vesicles through JNK-mediated exocytosis. Supporting this possibility, we found that blocking JNK signaling by overexpressing Puc or knocking down unc-13 significantly reduced the number of GFP-Wg-positive puncta in the RpS3/+ wing pouch (Figures 3H and 3I, quantified in Figure 3K). Additionally, consistent with the exocytosis-Dronc amplification loop, knocking down Dronc similarly reduced the number of GFP-Wg-positive puncta in the RpS3/+ wing pouch (Figure 3J, quantified in Figure 3K), suggesting that caspase activation reinforces Wg-vesicle accumulation.Figure 3. Dying cells secrete Wg via exocytosis in the M/+ wing pouch(A and B’’) Wing discs of GFP-Wg/+ (A), or RpS3/+, GFP-Wg/+ (B) flies, and dying cells in the wing pouch were visualized by anti-cleaved Dcp-1 staining (white). GFP-Wingless was visualized by anti-GFP staining (white). Arrowheads indicate massive cell death in the RpS3/+ wing pouch. Scale bar, 100 μm.(D–E”’) Immunofluorescent localization of the exosome marker CD63-mCherry, as visualized by anti-DsRed staining (magenta), and GFP-Wingless (white) in the wing pouch of GFP-Wg/+ (D), or RpS3/+, GFP-Wg/+ (E) flies expressing CD63-mCherry. The nuclei were visualized by DAPI staining (blue). The images correspond to the area enclosed by rectangle 1 in the schematic diagram of the wing disc (C). Orange arrowheads indicate the colocalization of GFP-Wingless-positive puncta with CD63-mCherry-positive puncta. Scale bar, 20 μm.(F–J’) The membrane-tethered anti-GFP nanobody (Vhh4-CD8-HA) was expressed in the wing pouch of GFP-Wg/+ (F), RpS3/+, GFP-Wg/+ (G), RpS3/+, GFP-Wg/+, nub-Gal4, UAS-Puc (H), RpS3/+, GFP-Wg/+, nub-Gal4, UAS-unc-13-RNAi (I), or RpS3/+, GFP-Wg/+, nub-Gal4, UAS-Dronc-RNAi (J) flies. GFP-Wingless was visualized by anti-GFP staining (white). Dying cells were visualized by anti-cleaved Dcp-1 staining in the wing discs (white). The images correspond to the area enclosed by the rectangle 2 in the schematic diagram of the wing disc (C). Immobilized GFP-Wg-positive puncta were frequently observed in the area of morphological dying cells in the RpS3/+ wing pouch (Indicated by orange arrowheads). Scale bar, 50 μm.(K) Boxplot with individual dots representing the number of GFP-wingless-positive puncta per rectangle-2 in the wing pouch (C). The respective genotypes are shown in (F) (n = 9, number of wing pouches), (G) (n = 14), (H) (n = 11), (I) (n = 11), and (J) (n = 9). Thick line, median; ∗∗∗p < 0.001, ∗∗p < 0.01; Wilcoxon rank-sum test.
Furthermore, we observed a significant increase in extracellular GFP-positive puncta and extracellular CD63-mCherry-positive vesicles in the RpS3/+ wing pouch when GFP-Wg was captured on the cell surface using the Vhh4-CD8 morphotrap, compared to the wild-type control (Figures S3M and S3N, quantified in Figures S3O and S3P). These extracellular GFP-Wg-positive puncta occasionally colocalized with extracellular CD63-mCherry-positive vesicles (6.88% in the wild-type, and 16.45% in the RpS3/+ wing pouch region, characterized by massive cell death, as indicated by the arrowheads in Figures S3M and S3N). Together, these data suggest that dying cells secrete Wg via exocytosis in the M/+ wing pouch.
JNK signaling triggers exocytosis-mediated Wg secretion
Finally, we investigated whether JNK activation universally upregulates exocytosis and subsequent Wg secretion in contexts beyond the M/+ wing pouch. To address this, we generated JNK-activating clones through the overexpression of Eiger, the Drosophila homolog of tumor necrosis factor (TNF), in the eye discs. Eiger has been shown to specifically activate the JNK pathway through dTAK1 (Drosophila JNKKK) and Hep (Hemipterous; Drosophila JNKK).66
Although dying cells were observed in Eiger-overexpressing clones generated in the eye discs (Figure S4B, compared to Figure S4A, quantified in Figure S4D), the overall tissue size remained unchanged (quantified in Figure S4E), suggesting that neighboring cells proliferate to maintain tissue homeostasis. We found that the elimination of Eiger-expressing clones and the number of dying cells were significantly suppressed by knocking down unc-13 in these clones (Figures 4D and S4C, quantified in Figures 4E and S4D), whereas unc-13-RNAi expression alone did not affect tissue growth (Figure 4B, compared to Figure 4A, quantified in Figures 4E and S4E). Similarly, in the wing disc, unc-13 knockdown also significantly suppressed the elimination of Eiger-expressing clones (Figures S4F–S4I, quantified in Figure S4J), suggesting that exocytosis-mediated elimination of JNK-activating cells is not tissue-specific.Figure 4JNK signaling universally triggers exocytosis-mediated Wg secretion(A–D) Eye disc bearing eyFLP-induced MARCM clones of UAS-yellow-RNAi (A), UAS-unc-13-RNAi (B), UAS-Eiger^W^ +UAS-yellow-RNAi (C), or UAS-Eiger^W^+UAS-unc-13-RNAi (D) cells. Scale bar, 100 μm.(E) Boxplot with individual dots representing the clone size (% of total clone area per eye disc area) in genotypes shown in (A) (n = 23, number of eye discs), (B) (n = 26), (C) (n = 32), and (D) (n = 17). Thick line, median; ∗∗∗p < 0.001; n.s., not significant; Wilcoxon rank-sum test.(F–H) Eye disc bearing eyFLP-induced MARCM clones of UAS-CD63-mCherry (F), UAS-Eiger^W^ +UAS-CD63-mCherry (G), or UAS-Eiger^W^+UAS-unc-13-RNAi +UAS-CD63-mCherry (H) cells, stained with anti-DsRed antibody (magenta) to detect extracellular CD63-mCherry. Scale bar, 10 μm.(I) Boxplot with individual dots representing the relative extracellular CD63-mCherry area in the eye disc for genotypes shown in (F) (n = 17, number of clones), (G) (n = 21), and (H) (n = 17). Thick line, median; ∗∗∗p < 0.001; Wilcoxon rank-sum test.(J–K’’’) Eye disc bearing eyFLP-induced MARCM clones of GFP-Wg/+ (J), or UAS-Eiger^KB^, GFP-Wg/+ (K) cells stained with anti-GFP antibody (white). Scale bar, 20 μm.(L) Boxplot with individual dots representing the relative GFP-Wg area per clone area of the eye disc for genotypes shown in (J) (n = 11, number of clones), and (K) (n = 9). Thick line, median; ∗∗∗, p < 0.001; Wilcoxon rank-sum test.(M–O) Eye disc bearing eyFLP-induced MARCM clones of wild-type (M), UAS-Eiger^KB^ (N), or UAS-Eiger^KB^, wg/+ (O) cells. Scale bar, 100 μm.(P) Boxplot with individual dots representing the clone size (% of total clone area per eye disc area) in genotypes shown in (M) (n = 12, number of eye discs), (N) (n = 11), and (O) (n = 9). Thick line, median; ∗∗∗p < 0.001, ∗∗p < 0.01; Wilcoxon rank-sum test.
In addition, we observed a significant increase in extracellular CD63-mcherry-positive vesicles and GFP-Wg-positive puncta, derived from a knock-in allele,65 in Eiger-overexpressing clones in the eye discs, compared to the control (Figures 4F, 4G, 4J’’, and 4K’’, quantified in Figures 4I and 4L). Furthermore, the elimination of Eiger-expressing clones in the eye discs was significantly suppressed to a moderate extent when the Wg signaling gradient was reduced by deleting one copy of the wg gene (Figures 4M–4O, quantified in Figure 4P), raising the intriguing possibility that exocytosis-mediated Wg secretion from dying cells may contribute to their cell death and proliferation of neighboring cells.
Interestingly, knocking down Dronc in the clones of cells activating JNK signaling by overexpressing the constitutive active form of Hep (Hep^CA^) reduced the increased extracellular CD63-mcherry-positive vesicles (Figures S4K and S4L, quantified in Figure S4M). This suggest that Dronc is crucial for promoting exocytosis downstream of JNK signaling, and that JNK activation alone is not sufficient to maintain exocytic activity in the absence of caspase activation.
Together, these data suggest that JNK signaling promotes exocytosis and the subsequent Wg secretion, which may play a role for maintaining tissue homeostasis. Notably, our findings also highlight a reciprocal interplay between caspase activation and exocytosis, where each process may reinforce the other to sustain apoptotic signaling downstream of JNK signaling.
Discussion
Our current study reveals that exocytosis is crucial for massive cell turnover in M/+ wing morphogenesis, acting as a downstream event of JNK signaling. In dying cells, JNK signaling upregulates the expression of exocytosis-related genes and Wg, leading to increased Wg secretion. In parallel, elevated exocytosis, in turn, promotes caspase activation via an exocytosis-Dronc amplification loop, wherein exocytosis enhances Dronc activity, which in turn reinforces exocytosis, driving sustained apoptotic signaling (Figure 5). This exocytosis-mediated Wg secretion and apoptotic feedback loop is a key downstream event of JNK signaling in both M/+ and Eiger-overexpressing clones, suggesting that exocytosis actively reinforces the apoptotic response, potentially by enhancing JNK signaling to trigger cell death.Figure 5A model for ensuring robust coordination of tissue growth in M/+ animalsDuring the larval period of M/+ animals, JNK signaling is activated in the M/+ wing pouch, which induces the expression of exocytosis-related genes and Wg, leading to the secretion of Wg through exocytosis in dying cells. In addition, elevated exocytosis induces cell death via an exocytosis-Dronc amplification loop. This process subsequently promotes the proliferation of neighboring cells, establishing their apoptotic/proliferative status and resulting in significant cell turnover in the M/+ wing pouch. Wg (red), vesicles (yellow), and CD63 (purple) are indicated in the schematic.
Notably, impairment of exocytosis in dying cells in RpS3/+ animals leads to phenotypic variations in adult wings (Figures 2O and 2P in this study). This highlights the importance of proper exocytosis in maintaining cell turnover and ensuring developmental robustness, which may provide the flexibility needed to adapt developmental programs to varying genetic and environmental conditions.
We identified Wg as an important factor secreted through exocytosis from dying cells, potentially essential for the proliferation of neighboring living cells to maintain tissue homeostasis. Previous studies have shown that “undead” cells, stimulated toward apoptosis but kept alive by the caspase inhibitor p35, are able to promote the proliferation of neighboring cells in the Drosophila epithelium.67^,^68^,^69 Several mitogens, including Wg, Dpp, and Spitz (a EGF homolog) have been identified as being produced by apoptotic cells in a JNK-dependent manner.18^,^58^,^70 In addition, Hedgehog (Hh)15 have also been identified as products of apoptotic cells. Furthermore, during cell competition between wild-type (winners) and M/+ cells (losers) in the Drosophila epithelium*,* prospective loser cells secrete upd3, which promotes competitive elimination by winner cells.12 Hence, in addition to Wg, other mitogens could also be secreted thorough exocytosis from dying cells, potentially contributing to massive cell turnover in the M/+ wing pouch.
Interestingly, it has been reported that apoptosis triggers a wave of ERK/Akt activity pulses that radially propagate across approximately three healthy cells in a cultured human MCF10A monolayer.71 Notably, caspase-activated dying cells have also been shown to induce ERK activation in neighboring cells, which is essential for inhibiting caspase activation and preventing the elimination of clustered cells in the Drosophila pupal notum.72 Together, these previous studies raise the intriguing possibility that non-autonomous ERK activation and JNK-mediated exocytosis, leading to Wg secretion, may cooperate to maintain tissue homeostasis by modulating the balance between cell death and subsequent cell proliferation. How these pathways interact to orchestrate such a balance remains an open question, requiring further investigation to uncover the precise mechanisms underlying this coordination.
Previous studies have identified exosomes as carriers of Wnt/Wg in the extracellular space of both mammalian and Drosophila cells, including wing disc cells.33^,^61^,^62^,^63^,^64^,^73 Concurrently, alternative mechanisms for Wg transport, such as those involving lipoprotein particles or a lipocalin Swim in the Drosophila wing disc, have been reported.32^,^74 Additionally, the cell-surface proteoglycan Dally-like-protein (Dlp) has been reported to enable long-range signaling of the palmitoylated Wg.65 Intriguingly, Wnt/Wg transport has also been observed through filopodia-like cellular extensions known as cytonemes.75^,^76^,^77^,^78 In our study of the M/+ wing disc, a model characterized by massive cell turnover, we observed partial colocalization of GFP-Wg puncta with extracellular CD63-mCherry-positive vesicles. Our data also suggest that Wg secretion through exocytosis may not uniformly occur among all dying cells within this context. This disparity suggests that while exosomes from dying cells significantly contribute to Wg transport within the M/+ wing pouch, other pathways may also be operative.
Rp genes are crucial in a broad range of organisms, from yeast to humans, frequently exhibiting haploinsufficient mutant phenotypes. Notably, heterozygous mutations in genes encoding ribosomal proteins or ribosomal biogenesis factors are linked to tissue-specific human diseases known as ribosomopathies.79 In patients affected by these conditions, some tissues may preserve normal patterning and functions, potentially by buffering stresses caused by ribosomal mutations. Given the evolutionary conservation of the molecules we identified in Drosophila, regulating cell turnover through exocytosis in dying cells may represent evolutionarily conserved strategy to prevent developmental abnormalities in ribosomal protein mutants.
Limitations of the study
Our genetic data suggest that JNK signaling is required for elevated exocytosis, but sustained exocytic activity depends on caspase activation, making it difficult to fully separate the effects of JNK signaling and apoptosis on exocytosis. While our findings support a positive feedback loop between exocytosis and caspase activity, the molecular basis of this interplay remains unclear. Furthermore, although we show that Wg is secreted from dying cells via exocytosis, how this signal is received by neighboring cells and leads to compensatory proliferation is not fully understood. It also remains unclear how the balance between cell proliferation and cell death is regulated to maintain coordinated tissue growth. Future studies investigating the molecular mechanisms as well as the spatial and temporal dynamics of Wg reception and proliferative responses will be essential for a comprehensive understanding of tissue growth coordination.
Resource availability
Lead contact
Further information and requests for resources and reagents should be directed to and will be fulfilled by the lead contact, Shizue Ohsawa ([email protected]).
Materials availability
This study did not generate new unique reagents.
Data and code availability
- •All data reported in this paper will be shared by the lead contact upon request. The sequencing data have been deposited at DDBJ: PRJDB20450.
- •This paper does not report original code.
- •Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
Acknowledgments
We thank Takefumi Kondo, Keisuke Ikawa, Kiichiro Taniguchi, Rina Nagata, and Yukari Sando for discussions, Susumu Tsutsumi, Nana Watanabe, Mina Hoshino, Tomoko Furukawa, and Sayako Suzuki for technical support, and NGS core facility of the Graduate Schools of Biostudies, Kyoto University for supporting the RNA-seq analysis. We also thank Yash Hiromi, Gary Struhl, Maoto Sato, Jean-Paul Vincent, Sharad Kumar, Konrad Basler, the Bloomington Drosophila Stock Center (Indiana), the Vienna Drosophila Resource Center (Vienna), the National Institute of Genetics Stock Center (Mishima), and the Drosophila Genomics and Genetic Resources (Kyoto) for fly stocks. Cell sorting using 10.13039/100017412BD FACS Aria II were performed at the Medical Research Support Center, 10.13039/100022395Graduate School of Medicine, 10.13039/501100005683Kyoto University, supported by Basis for Supporting Innovative Drug 10.13039/100002806Discovery and Life Science Research (BINDS) from 10.13039/100009619AMED (grant no. JP22ama121034). This work was supported in part by 10.13039/501100001700MEXT grant-in-aid (10.13039/501100001691KAKENHI) for Transformative Research Area (A) (grant nos. 20H05945 to S.O., 21H05284 to T.I.), the Scientific Research (B) (grant no. 22H02616 to S.O.), and Challenging Exploratory Research (grant no. 21K19257 and 24K21967 to S.O.), 10.13039/501100002241Japan Science and Technology Agency (10.13039/501100020963Moonshot Research & Development: Grant Number JPMJPS2022 to SO), AMED-10.13039/501100003382CREST, 10.13039/100009619Japan Agency for Medical Research and Development (22gm1710002h0001 to T.I.), 10.13039/100009619Japan Agency for Medical Research and Development (21gm5010001 to T.I.), and 10.13039/100007449Takeda Science Foundation to S.O.
Author contributions
Conceptualization: N.A., T.I., and S.O. data curation: N.A., T.I., and S.O. formal analysis: N.A. funding acquisition: S.O. and T.I. investigation: N.A., Y.Y., and S.O. methodology: N.A. project administration: S.O. resources: S.O. supervision: S.O. visualization: N.A., T.I., and S.O. writing – original draft: S.O. and N.A. writing – review and editing: N.A., T.I., and S.O.
Declaration of interests
The authors declare no competing interests.
STAR★Methods
Key resources table
REAGENT or RESOURCESOURCEIDENTIFIERAntibodiesRat anti-GFP antibodyNacalai TesqueCat#04404-84; RRID: AB_10013361Rabbit anti-cleaved PARP (Asp214) antibodyCell Signaling TechnologyCat#9541; RRID: AB_331426Rabbit anti-phospho-histone H3 (Ser10) antibodyCell Signaling TechnologyCat#9706; RRID: AB_331748Living Colors® dsRed Polyclonal AntibodyTakara BioCat#632496; RRID: AB_10013483Rabbit anti-GFP antibodyThermo Fisher ScienceCat#A6455; RRID: AB_221570Cleaved Drosophila Dcp-1 (Asp216) AntibodyCell Signaling TechnologyCat#9578; RRID: AB_2721060mouse anti-β-galactosidase antibodySigma-AldrichCat#G8021; RRID: AB_259970anti-rabbit Alexa 488Thermo Fisher ScienceCat#A-11034; RRID: AB_2576217anti-rabbit Alexa 546Thermo Fisher ScienceCat#A-11035; RRID: AB_2534093anti-rat Alexa 488Thermo Fisher ScienceCat#A-11006; RRID: AB_2534074anti-chicken Alexa 488Thermo Fisher ScienceCat#A-11039; RRID: AB_2534096anti-rabbit Alexa 647Thermo Fisher ScienceCat#A-21245; RRID: AB_2535813anti-mouse Alexa 647Thermo Fisher ScienceCat#A-21237; RRID: AB_2535806Chemicals, peptides, and recombinant proteinsSlowFade Diamond Antifade MountantThermo Fisher ScienceCat#S36972DAPISigma-AldrichCat#D9542GlycerolSigma-AldrichCat#G5516n-Propyl GallateKANTO CHEMICALCat#32465-31Schneider’s Drosophila MediumThermo Fisher ScienceCat#21720024SYTOX Blue Nucleic Acid StainThermo Fisher ScienceCat#11348Normal Goat SerumJackson ImmunoResearchCat#005-000-121; RRID: AB_2336990InsulinSigma-AldrichCat#I0516; CAS: 11070-73-810x TrypLE SelectGibcoCat#A1217701Critical commercial assaysNucleoSpin RNA XS KitTaKaRaCat#740902NEBNext Ultra II Directional RNA Library Prep Kit for IlluminaNEW ENGLAND BiolabsCat#E7760SDeposited dataRNA-sequencing dataThis paperDDBJ: PRJDB20450Experimental models: Organisms/strainsDrosophila melanogaster: UAS-EGFP.CD63Bloomington Drosophila Stock CenterBDSC91390Drosophila melanogaster: UAS-CD63.mCherryBloomington Drosophila Stock CenterBDSC91389Drosophila melanogaster: UAS-Syt1-eGFPBloomington Drosophila Stock CenterBDSC6925Drosophila melanogaster: 20XUAS-IVS-GCaMP6mBloomington Drosophila Stock CenterBDSC42750Drosophila melanogaster: RpS3^Plac92^Bloomington Drosophila Stock CenterBDSC5627Drosophila melanogaster: RpL19^K03704^Kyoto Stock Center (DGRC)DGRC102285Drosophila melanogaster: UAS-unc-13-RNAiBloomington Drosophila Stock CenterVDRC33609Drosophila melanogaster: UAS-unc-13-RNAiNational Institute of GeneticsNIG2999R-2Drosophila melanogaster: UAS-LexA-RNAiBloomington Drosophila Stock CenterBDSC67947Drosophila melanogaster: UAS-Rab3-RNAiNational Institute of GeneticsNIG7576R-3Drosophila melanogaster: UAS-Rab3-RNAiBloomington Drosophila Stock CenterBDSC31691Drosophila melanogaster: UAS-SNAP25-RNAiNational Institute of GeneticsNIG HMS01367Drosophila melanogaster: UAS-SNAP25-RNAiBloomington Drosophila Stock CenterBDSC27306Drosophila melanogaster: UAS-Syt1-RNAiNational Institute of GeneticsNIG3139R-1Drosophila melanogaster: UAS-Syt1-RNAiBloomington Drosophila Stock CenterBDSC31668Drosophila melanogaster: UAS-Rab27-RNAiNational Institute of GeneticsNIG14791R-2Drosophila melanogaster: UAS-Rab27-RNAiBloomington Drosophila Stock CenterBDSC31887Drosophila melanogaster: UAS-ALiX-RNAiNational Institute of GeneticsNIG HMS00298Drosophila melanogaster: UAS-ALiX-RNAiVienna Drosophila Resource CenterVDRC32049Drosophila melanogaster: Gaq^221C^Bloomington Drosophila Stock CenterBDSC30744Drosophila melanogaster: UAS-Gaq-RNAiNational Institute of GeneticsNIG17759R-2Drosophila melanogaster: UAS-Gaq-RNAiNational Institute of GeneticsNIG HMJ30300Drosophila melanogaster: UAS-Gaq-RNAiBloomington Drosophila Stock CenterBDSC JF02390Drosophila melanogaster: UAS-Plc21C-RNAiNational Institute of GeneticsNIG4574R-2Drosophila melanogaster: UAS-Plc21C-RNAiNational Institute of GeneticsNIG4574R-3Drosophila melanogaster: norpA^7^Kyoto Stock Center (DGRC)DGRC108362Drosophila melanogaster: UAS-norpA-RNAiVienna Drosophila Resource CenterVDRC105676Drosophila melanogaster: UAS-norpA-RNAiBloomington Drosophila Stock CenterBDSC JF02390Drosophila melanogaster: UAS-PucMartín-Blanco et al.80N/ADrosophila melanogaster: wg^l−^^8^Bloomington Drosophila Stock CenterBDSC5351Drosophila melanogaster: UAS-yellow-RNAiNational Institute of GeneticsNIG3757R-1Drosophila melanogaster: UAS-Dronc^DN^Quinn et al.81N/ADrosophila melanogaster: UAS-Dronc-RNAiVienna Drosophila Resource CenterVDRC23033Drosophila melanogaster: GFP-Wingless^S239A^McGough et al.65N/ADrosophila melanogaster: UAS-VHH4-CD8-HAMcGough et al.65N/ADrosophila melanogaster: UAS-Eiger^+W^Igaki et al.82N/ADrosophila melanogaster: UAS-Eiger^KB^Moreno et al.83N/ADrosophila melanogaster: UAS-Hep^CA^Bloomington Drosophila Stock CenterBDSC 58781Drosophila melanogaster: wg^026578^Bloomington Drosophila Stock CenterBDSC 11205Software and algorithmsLeica LAS AF Lite softwareLeica microsystemshttps://leica-las-af-lite.software.informer.com/4.0/ExcelMicrosofthttps://www.microsoft.com/en-gb/microsoft-365/excelFiji softwareSchindelin et al.84RRID: SCR_002285RStudio (version 1.2.5033)RStudio, Inc.https://rstudio.comKaleidaGraphSynergy Softwarehttps://www.synergy.com/OtherLeica TCS SPE MicroscopeLeica Microsystemshttps://www.leica-microsystems.com/Leica STELLARIS 5 MicroscopeLeica Microsystemshttps://www.leica-microsystems.com/Leica M205C stereo microscopeLeica Microsystemshttps://www.leica-microsystems.com/ZEISS LSM 900 Microscopehttps://www.zeiss.com/microscopy/en/home.html?vaURL=www.zeiss.com/microscopyOlympus FV3000 MicroscopeOlympus Life Sciencehttps://www.olympus-lifescience.com/ja/
Experimental model and study participant details
Fly strains
Drosophila melanogaster fly stocks were cultured at 25°C on standard fly food. Larval experiments were conducted with individuals of both sexes, whereas experiments involving adult wings were performed using only females. All experiments were conducted in accordance with the guidelines of Nagoya University and were approved by the Institutional Animal Care and Use Committee (protocol number: 560).
To detect dying cells in the wing pouch, the following strains were used: nub-Gal4; UAS-CD8-PARP-Venus (control); nub-Gal4; UAS-CD8-PARP-Venus, RpS3^Plac92^ (BL5627)/TM6B (RpS3/+ tester). Additional strains were used as follows: UAS-EGFP.CD63 (BL91390), UAS-CD63.mCherry (BL91389), UAS-Syt1-eGFP (BL6925), RpL19^K03704^ (DGRC102285), 20XUAS-IVS-GCaMP6m (BL42750), UAS-unc-13-RNAi (VDRC33609, NIG2999R-2), UAS-LexA-RNAi (BL67947), UAS-Rab3-RNAi (NIG7576R-3, BL31691), UAS-SNAP25-RNAi (HMS01367, BL27306), UAS-Syt1-RNAi (NIG3139R-1, BL31668), UAS-Rab27-RNAi (NIG14791R-2, BL31887), UAS-ALiX-RNAi (HMS00298, VDRC32049), UAS-Gαq-RNAi (NIG17759R-2, HMJ30300, JF02390), Gαq^221C^ (BL30744), UAS-Plc21C-RNAi (NIG4574R-2, 4574R-3), UAS-norpA-RNAi (VDRC105676, JF01585), norpA7 (DGRC108362)UAS-y-RNAi (NIG3757R-1), wg^l−8^ (BL5351), wg^026578^ (BL11205), UAS-Puc,80 UAS-Dronc^DN^(Sharad Kumar),81 UAS-Dronc-RNAi (VDRC23033), GFP-Wingless^S239A^ (JP. Vincent),65 UAS-VHH4-CD8-HA (JP. Vincent),65 UAS-Eiger^+W^,82 UAS-Eiger^KB^ (Konrad Basler),83 and UAS-Hep^CA^ (BL58781).
Method details
Histology
Wandering third-instar larvae were dissected in phosphate buffered saline (PBS) and fixed with 4%-Paraformaldehyde (PFA) for 5 min on ice, followed by 20 min at room temperature. The fixed larvae were washed 3 times with PBT (PBS +0.1% Triton X-100) for 20 min each, and were then blocked with PBTn (5% goat serum (Jackson ImmunoResearch, #005-000-121) + PBT) solution for 30 min. For immunostaining, they were incubated overnight at 4°C with primary antibodies in PBTn. After four 30-min washes with PBT, the samples were re-blocked in PBTn for 30 min, then incubated with secondary antibodies for 2 h at room temperature. Following another set of four 30-min washes with PBT, the samples were mounted using either SlowFade Diamond Antifade Mountant (Invitrogen, #S36972) or an alternative anti-fade mounting medium. The latter was composed of 70% Glycerol (Sigma, #G5516), 0.2% n-Propyl Gallate (KANTO CHEMICAL, #32465-31), and supplemented with DAPI (Sigma, #D9542). For extracellular immunostaining, larvae were dissected in ice-cold Schneider’s Drosophila Medium (Gibco, #21720024) and incubated with primary antibodies for 2 h on ice. The larvae were then rinsed with ice-cold Schneider’s Drosophila Medium followed by PBS and fixed with 4% PFA for 40 min at room temperature. After fixation, samples were washed with PBS. Subsequent processing was the same as in the preceding protocol. Images were taken with Leica SP5, Leica SP8, Leica SPE, Leica STELLARIS and ZEISS LSM900 confocal microscopes. Primary antibodies used are as follows; rat anti-GFP antibody (Nacalai Tesque, #04404-26, 1:1000) (Extracellular GFP, 1:300), rabbit anti-GFP antibody (Invitrogen, #A6455) (Extracellular GFP, 1:75), rabbit anti-cleaved PARP (Asp214) antibody (Cell Signaling Technology (CST), #9541, 1:200), rabbit anti-phospho-histone H3 (Ser10) antibody (CST, #9706, 1:100), mouse anti-β-galactosidase antibody (Sigma, #G8021, 1:500), rabbit anti-DsRed antibody (Takara Bio Clontech (CLN), #632496, 1:250) (Extracellular dsRed, 1:75), and rabbit anti-Cleaved Drosophila Dcp-1 (Asp216) antibody (CST, #9678, 1:100) for 3 days. Secondary antibodies used are as follows; Goat anti-rabbit Alexa 488 (1:250, Invitrogen #A11034), Goat anti-rabbit Alexa 546 (1:250, Invitrogen #A11035), Goat anti-rabbit Alexa 647 (1:250, Invitrogen #A21245), Goat anti-rat Alexa 488 (1:250, Invitrogen #A11006), Goat anti-chicken Alexa 488 (1:250, Invitrogen, #A11039), and Goat anti-mouse Alexa 647 (1:250, Invitrogen #A21237) antibodies.
Quantification of puncta number
The vesicle marker EGFP-CD63, CD63-mCherry, and Syt1-eGFP were expressed in the wing pouch under the control of nub-gal4 driver. In the region where massive cell death is observed in the M/+ wing disc, these markers, in addition to GFP-Wingless signals, were acquired at the confocal plane exhibiting the highest signal intensity. CD63-mCherry were also expressed within the mosaic clones of the eye-antenna disc. Puncta were manually counted using Fiji software84 and the data were analyzed with KaleidaGraph (Synergy Software).
Quantitative analysis of GFP-Wingless and CD63-mCherry colocalization
To analyze the overlap between GFP-Wingless-positive puncta and CD63-mcherry-positive vesicles, their colocalization was manually quantified in the regions exhibiting massive cell death. This quantification was conducted using the confocal plane that displayed the highest signal intensity. Images were manually thresholded to eliminate background signals, and the areas of overlap were calculated using Fiji software, followed by further analyses with KaleidaGraph.
Calcium time-lapse imaging and quantification
To monitor Ca^2+^ activity, wandering third-instar larvae were dissected and cultured in Schneider’s Drosophila medium supplemented with 2% FBS, 1% Insulin solution (Sigma-Aldrich, #I0516), 3% fly extract (VDRC, #VFE1.25), and 2% low-melting agarose (Sigma-Aldrich, #A4018)/PBS. Imaging was conducted at 10 s intervals for 20-min with a Leica STELLARIS confocal microscope.49^,^85 For quantitative analysis, dying cells exhibiting continuous calcium activity were manually counted using Fiji software and the data were analyzed with KaleidaGraph. To monitor dying cells, wing discs were incubated in the above medium with 1μM SYTOX (Invitrogen, #11348) for 10 min before imaging.
Detection of dying cells and quantification using CD8-PARP-Venus
To detect dying cells, the activated-caspase-3 indicator CD8-PARP-Venus was expressed in the pouch under the control of nub-gal4 driver. Cleaved PARP signals were acquired at the confocal plane where signal intensity was highest. The number of these signals in the wing pouch was manually measured using Fiji software and the data were analyzed with KaleidaGraph.
Quantification of pH3-positive area
For the analysis of pH3-positive cells, total area of pH3-positive cells and the size of the wing pouch area were automatically measured from confocal slices of z stack images compressed using maximum projection functions in Fiji software, as previously described.86 The acquired data were then analyzed with KaleidaGraph.
Measurement of adult wings
Left and right wings of female flies were rinsed with xylene and mounted in a Canada Balsam (Nacalai Tesque). Images of the wings were acquired using Leica M205C stereo microscope.
Quantification of clone size
For the analysis of clone size in the eye disc and wing disc, total clone area as a percentage of the disc area (%) was measured from XY confocal images using Fiji software, as previously described.86 The acquired data were then analyzed with KaleidaGraph.
FACS and mRNA-seq analysis
GFP+ wing pouch cells were isolated using a FACS Aria II cell sorter (BD Bioscience). Total RNA was then extracted for RNA sequencing analysis, following the method previously described.87 Briefly, third-instar larvae were dissected in ice-cold PBS. The wing disc cells were dissociated using Trypsin (10x TrypLE Select) (Gibco, #A1217701) at 37°C for 20 min. The enzymatic reaction was stopped with Schneider’s medium containing 5% FBS, and GFP-positive cells from wing pouches were sorted using a FACS Aria II cell sorter (BD Bioscience). RNA samples were extracted using NucleoSpin RNA XS Kit (TaKaRa, #740902). The quality of the extracted total RNA was assessed using an Agilent 2100 Bioanalyzer, and then Strand-specific Libraries for mRNA-seq were prepared using the NEBNext Ultra II Directional RNA Library Prep Kit for Illumina (E7760S). For RNA-seq, NextSeq 500/550 High output Kit v2.5 was utilized on a NextSeq 500 (Illumine), generating single-end reads at a length of 86 bases.
RNA-seq data analysis
Each genotype was analyzed across three independent replicates. Each sequencing experiment generated more than 16 million raw reads. These reads were then quality-filtered using Trim_Galore! (v0.6.5) (https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/) with the default setting to remove low-quality reads and adaptor sequences. The filtered reads were mapped to the Drosophila melanogaster genome (Ensembl BDGP6.32), obtained from Illumina iGenomes using STAR.88 In the RpS3/+, nub-Gal4, UAS-Puc experiment, one sample exhibited only 65.69% of reads being uniquely mapped, which was lower compared to other experiments where more than 80% of reads were uniquely mapped in each sample. The number of reads mapped to each gene was quantified using RSEM (v1.3.3).89 Normalization was performed with calcNormFactors function in edgeR (v3.30.3).90 Significantly differentially expressed genes were identified with an FDR<0.05 using the glmQLFit and glmQLFTest function in edgeR (R version 4.0.1, limma 3.44.3). Furthermore, GO enrichment analysis was performed on the 362 genes regulated by JNK signaling, utilizing the online tool available at “Gene Ontology” (https://geneontology.org).
Quantification and statistical analysis
Statistical analysis was performed, and boxplot graphs were generated using KaleidaGraph version4.5 (Synergy Software). All experiments shown in the same graph were conducted at the same time. Thick line in the boxplot represents the median. Significance levels are indicated as follows: ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001, and n.s.; not significant. The Wilcoxon rank-sum test was used for non-parametric comparisons. Statistical details are included in the Figure legends, and all statistical data are summarized in Table S4.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cmejla R.Cmejlova J.Handrkova H.Petrak J.Pospisilova D.Ribosomal protein S 17 gene (RPS 17) is mutated in Diamond-Blackfan anemia Hum. Mutat.2820071178118210.1002/humu.2060817647292 · doi ↗ · pubmed ↗
- 2Doherty L.Sheen M.R.Vlachos A.Choesmel V.O'Donohue M.F.Clinton C.Schneider H.E.Sieff C.A.Newburger P.E.Ball S.E.Ribosomal protein genes RPS 10 and RPS 26 are commonly mutated in Diamond-Blackfan anemia Am. J. Hum. Genet.86201022222810.1016/j.ajhg.2009.12.01520116044 PMC 2820177 · doi ↗ · pubmed ↗
- 3Campagnoli M.F.Ramenghi U.Armiraglio M.Quarello P.Garelli E.Carando A.Avondo F.Pavesi E.Fribourg S.Gleizes P.E.RPS 19 mutations in patients with Diamond-Blackfan anemia Hum. Mutat.29200891192010.1002/humu.2075218412286 · doi ↗ · pubmed ↗
- 4Choesmel V.Fribourg S.Aguissa-TouréA.H.Pinaud N.Legrand P.Gazda H.T.Gleizes P.E.Mutation of ribosomal protein RPS 24 in Diamond-Blackfan anemia results in a ribosome biogenesis disorder Hum. Mol. Genet.1720081253126310.1093/hmg/ddn 01518230666 · doi ↗ · pubmed ↗
- 5Wang R.Yoshida K.Toki T.Sawada T.Uechi T.Okuno Y.Sato-Otsubo A.Kudo K.Kamimaki I.Kanezaki R.Loss of function mutations in RPL 27 and RPS 27 identified by whole-exome sequencing in Diamond-Blackfan anaemia Br. J. Haematol.168201585486410.1111/bjh.1322925424902 · doi ↗ · pubmed ↗
- 6Danilova N.Gazda H.T.Ribosomopathies: how a common root can cause a tree of pathologies Dis. Model. Mech.820151013102610.1242/dmm.02052926398160 PMC 4582105 · doi ↗ · pubmed ↗
- 7Lipton J.M.Ellis S.R.Diamond-Blackfan anemia: diagnosis, treatment, and molecular pathogenesis Hematol. Oncol. Clin. North Am.23200926128210.1016/j.hoc.2009.01.00419327583 PMC 2886591 · doi ↗ · pubmed ↗
- 8Bolze A.Mahlaoui N.Byun M.Turner B.Trede N.Ellis S.R.Abhyankar A.Itan Y.Patin E.Brebner S.Ribosomal protein SA haploinsufficiency in humans with isolated congenital asplenia Science (New York, N.Y.)340201397697810.1126/science.123486423579497 PMC 3677541 · doi ↗ · pubmed ↗