Rubussemiplenus (Rosaceae), a new species with naturally occurring semi-double flowers from Hunan, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
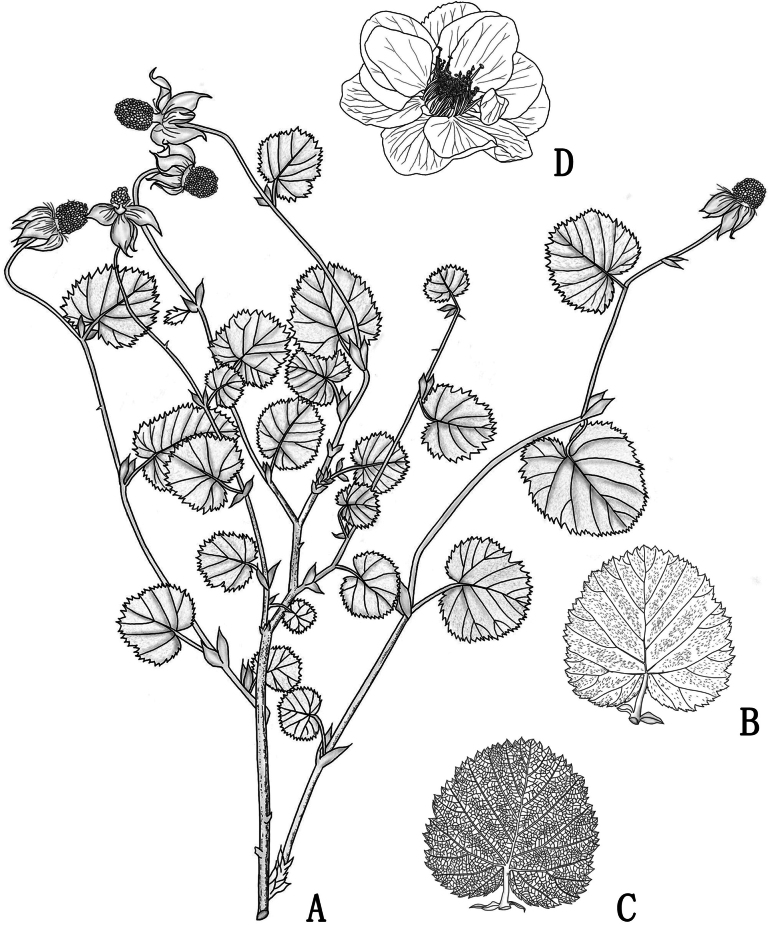 Figure 1
Figure 1 Figure 2
Figure 2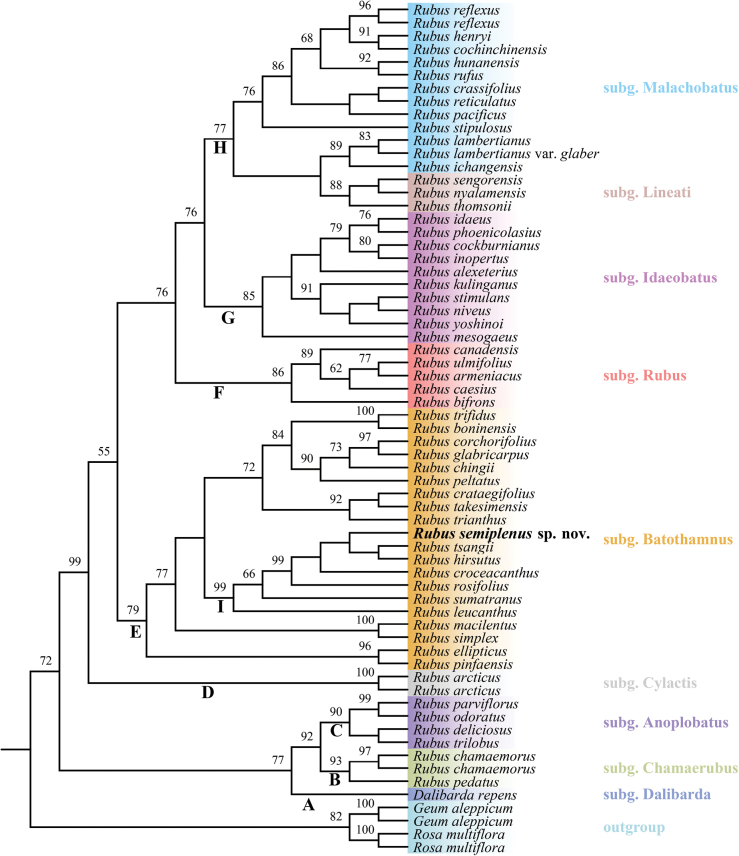 Figure 3
Figure 3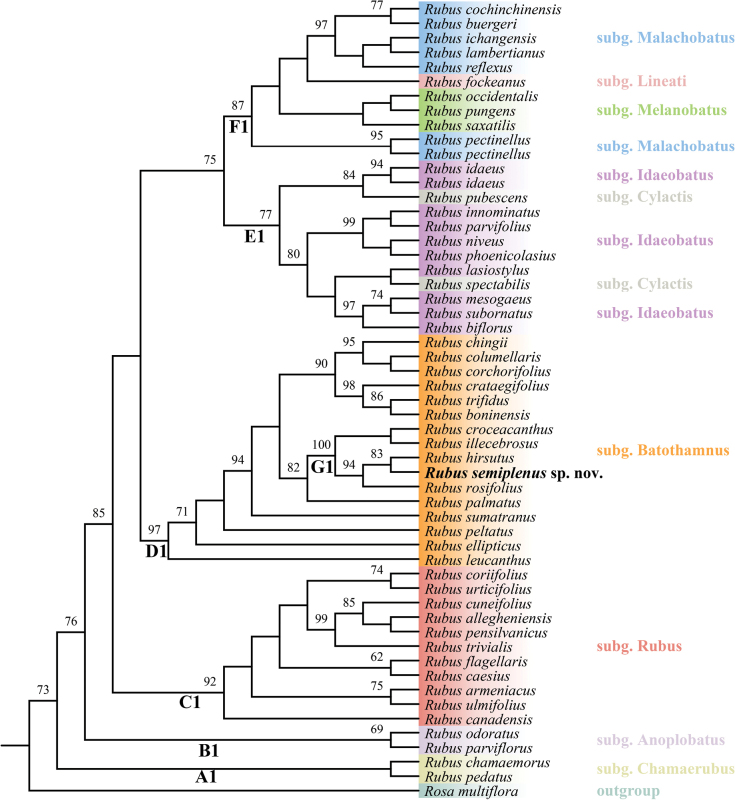 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Taxonomy and Phylogenetics · Horticultural and Viticultural Research · Botanical Studies and Applications
Introduction
Rubus L. is one of the largest genera in the Rosaceae family, belonging to the tribe Rubeae of subfamily Rosoideae (Yu and Lu 1985; Lu and Boufford 2003; Potter et al. 2007). This genus is found on all continents except Antarctica, with its primary distribution in East Asia and North America (Thompson 1995, 1997), and contains approximately 750–1000 species worldwide (Yu and Lu 1985; Jennings 1988). Rubus is one of the most taxonomically challenging genera of angiosperms (Wang et al. 2016; Okada et al. 2020; Gao et al. 2023; Huang et al. 2023). Species within this genus are typically shrubs, subshrubs, or perennial creeping herbs. Their leaves can be either compound or simple, and flowers are generally bisexual, although unisexual flowers occur rarely; fruits are fleshy aggregates of drupelets (Yu and Lu 1985; Kalkman 1993; Naruhashi 2001; Lu and Boufford 2003). Plants of Rubus have significant value in terms of food, medicine, ecology, and economy (Chen and Wang 2006; Foster et al. 2019).
The latest global revision of the genus Rubus was published nearly a century ago by Focke (1910, 1911, 1914), in which he divided the genus into twelve subgenera. In this classification, the majority of species were placed in R.subg.Rubus (= Eubatus), R.subg.Malachobatus (Focke) Focke, and R.subg.Idaeobatus (Focke) Focke. Focke’s subgeneric classification has been widely accepted and applied (Nakai 1918; Bailey 1941–1945; Naruhashi 2001). However, modern molecular systematics have not fully supported Focke’s classification (Alice and Campbell 1999; Yang and Pak 2006; Yang et al. 2012; Wang et al. 2016). More recently, Huang et al. (2023) proposed a new subgeneric classification system for Rubus, based on molecular phylogenetic analysis and morphological data, which divides the genus into 10 subgenera.
China has a rich diversity of Rubus, particularly in the southwestern region (Lu 1983; Gu et al. 2000; Lu and Boufford 2003). The first comprehensive revision of the genus Rubus in China was conducted by Yu and Lu (1985) for the “Flora Reipublicae Popularis Sinicae”, in which 194 species were recognized and classified into 8 sections and 24 subsections. Later, Lu and Boufford (2003) reviewed the Chinese species in the “Flora of China”, recognizing 208 species in China, with 139 of them being endemic. In recent years, numerous new species have been continuously discovered and published in China (e.g. Wang and Wang 2019; Wang et al. 2019; Wang et al. 2022; Chen et al. 2024a, 2024b), highlighting the need for continued investigation and study of Rubus diversity in China.
During a recent botanical expedition in Nanyue Hengshan National Nature Reserve, Hunan Province, China, we discovered an unknown species of Rubus characterized by its herbaceous to dwarf subshrub habit, simple leaves and semi-double flowers. Intrigued by its unique characteristics, we undertook a thorough examination of its morphological traits and performed phylogenetic analyses. The results of these studies clearly supported that this plant is a previously undescribed species.
Materials and methods
Morphological analyses
Morphological investigations were conducted in accordance with standard protocols for plant taxonomic surveys and herbarium-based classification (Davis and Heywood 1963). Specimen dissection and morphological examinations were performed at the laboratory in Yunnan University. A comprehensive list of morphological features was analyzed, including leaf shape (length, width), petiole length, stipule length, flower structure (size, number of petals, stamen count), fruit dimensions, seed morphology, and other distinctive vegetative and reproductive structures. These features were studied under a stereomicroscope (Olympus SZX2, Tokyo, Japan), and measurements were made using a ruler or a micrometer. Digital images available at the JSTOR Global Plants (http://plants.jstor.org/) and the Chinese Virtual Herbarium (https://www.cvh.ac.cn/), as well as relevant collections housed at CDBI, KUN, PE, PYU and YUKU (acronyms according to Thiers 2022), were examined and compared with the new species. Pertinent taxonomic literature (e.g. Focke 1910, 1911, 1914; Yu and Lu 1985; Lu and Boufford 2003; Wang et al. 2016; Wang et al. 2019) was extensively consulted.
Phylogenetic study
To determine the phylogenetic position of the new species, ITS and two plastid genes (rbcL and matK) were used as molecular markers. Total genomic DNA was extracted from silica-gel dried leaves of this new species using the DNAsecure Plant Kit (TIANGEN, Beijing, China).
The ITS primers used were ITS4 and ITS5, as detailed by White et al. (1990), and the PCR protocols were conducted according to Wang et al. (2016). Bidirectional sequencing was performed using an ABI 3730xL DNA Analyzer (Applied Biosystems) at the Kunming Branch of Beijing Qingke Biotechnology Co., Ltd. (Yunnan, China).
The new species was sequenced using the genome skimming technique (Straub et al. 2012; Zeng et al. 2018). DNA library construction and paired-end sequencing were completed by Novogene Bioinformatics Technology Co., Ltd. (Tianjin, China), with sequencing performed on the Illumina NovaSeq 6000 platform, generating approximately 3 Gbp of data. The chloroplast genomes (plastomes) were assembled from the clean data using GetOrganelle (Jin et al. 2020), and annotated the plastomes using GeSeq (Plastid Genome Annotator) (Tillich et al. 2017), followed by manual verification in Geneious (Kearse et al. 2012). Finally, the matk and rbcL gene sequences were extracted separately from the assembled complete chloroplast genome (GenBank: PV590107) using Geneious.
A total of 65 ITS sequence datasets were obtained in this study, including a new ITS sequence of R.semiplenus, 60 Rubus sequences and four outgroup sequences obtained from GenBank (Appendix 1: Table A1). In addition, 54 rbcL and matK sequences were downloaded from GenBank (Appendix 1: Table A2). Sequences were aligned using MAFFT (Katoh and Standley 2013). The combined dataset of rbcL + matK was generated using the Connected Sequence plugin in PhyloSuite (Zhang et al. 2020). The sequences and accession numbers used in this study are detailed in Appendix 1.
Phylogenetic analyses were performed using Maximum Likelihood (ML) methods. The gap sites were removed using trimAl (Capella-Gutiérrez et al. 2009). To determine the nucleotide substitution models for the dataset, we used ModelFinder (Kalyaanamoorthy et al. 2017) to evaluate multiple nucleotide substitution models based on Bayesian Information Criterion (BIC) and selected the best-fit model. ML analysis was then performed using IQ-tree (Nguyen et al. 2015) with 1000 bootstrap replicates. The resulting phylogenetic tree was visualized using iTOL (Letunic and Bork 2021).
Taxonomy
Rubus
semiplenus
Taxon classificationPlantaeRosalesRosaceae
Huan C. Wang, Ming Hong Li & T. T. Wang sp. nov.
64389E58-5BC8-5DF9-B7D5-F039D77ED660
urn:lsid:ipni.org:names:77362791-1
Type.
China • Hunan Province: Nanyue Hengshan National Nature Reserve, Hengyang City, 20 April 2022, M. H. Li et al. HY16383 (holotype: YUKU [YUKU02074893]!; isotypes: YUKU [YUKU02074894, YUKU02074895, YUKU02074896]!.)
Diagnosis.
Rubussemiplenus is most closely related to Rubushirsutus Thunb., but clearly distinguished morphologically from the latter by its habit herbs or dwarf subshrubs (vs. shrubs in R.hirsutus), stems villous hairs (vs. soft and glandular hairs), leaves simple (vs. imparipinnate, 3–5 foliolate), suborbicular (vs. ovate or broadly ovate), sepals ovate-orbicular (vs. ovate-lanceolate or triangular-lanceolate), petals dicyclic, semi-doubled flowers, with 10–12 (vs. petals 5).
Rubussemiplenus sp. nov. A habit B adaxial surface of leaf C abaxial surface of leaf D flower.
Description.
Herbs or dwarf subshrubs, 15–20 cm in height. Stem erect, terete, and branched. Branches and branchlets green, sparsely pilose and with minute prickles. Leaves simple. Stipules persistent, adnate to base of petiole, oblong-lanceolate, pilose, entire, 0.4–0.8 (–1.0) cm long, 0.1–0.3 cm wide, apex acuminate. Petiole 0.3–2.0 cm long, pilose. Leaf blade suborbicular, 0.7–2.5 cm long, 0.7–2.5 cm wide, both surfaces pilose, lateral veins 5 pairs, base cordate, margin doubly serrate, apex rounded. Inflorescences terminal, 2–3 flowers forming a corymb; bracts oblong-lanceolate, pilose, apex acuminate. Pedicel 3.0–4.5 cm long, densely pubescent. Flowers 2.0–3.5 cm in diameter; calyx light green, pubescent or tomentose abaxially, densely tomentose adaxially; sepals 5, ovate-orbicular, 0.5–1.0 (–1.2) cm long, 0.4–0.6 cm wide, entire, apex acuminate to caudate, spreading at anthesis, reflexed in fruit; petals semi-doubled, white, dicyclic, 10–12, ovate-orbicular or broadly ovate, longer than calyx, base shortly clawed; stamens straight, numerous, glabrous; filaments linear; pistils numerous, ca. 70–100, shorter than stamens; ovary villous at base. Aggregate fruit orange-red, subglobose, 0.5–0.7 cm in diameter, glabrous or sparsely pubescent.
Rubussemiplenus sp. nov. A habit B adaxial surface of leaf C abaxial surface of leaf D flower E calyx F infructescence G fruit.
Molecular phylogenetics.
The ITS, rbcL, and matK sequence lengths of Rubussemiplenus are 699 bp, 1428 bp, and 1512 bp, respectively, with GC contents of 54.1%, 43.0%, and 32.5%. Sequence alignment of the ITS dataset includes 65 sequences with 604 bp, among which 182 are parsimony-informative sites and 77 are singleton sites. The plastid (rbcL + matK) dataset consists of 2078 aligned positions, including 125 parsimony-informative sites and 158 singleton sites. According to the Bayesian Information Criterion (BIC), the best-fit models for the ITS and rbcL + matK datasets are TNe+R3 and K3Pu+F+R2, respectively.
The phylogenetic trees constructed using ITS and plastid (rbcL + matK) sequences are largely congruent in topology. In the ITS phylogenetic tree (Fig. 3), the genus Rubus was clearly resolved into eight clades, which largely correspond to the subgenera delineated by Huang et al. (2023). Rubussemiplenus nests within clade E, representing subgenus Batothamnus (Focke) E. H. L. Krause, and belongs to a well-supported subclade I (BS = 99), which includes R.tsangii Merr., R.hirsutus Thunb., R.croceacanthus H. Lév., R.rosifolius Sm., R.sumatranus Miq and R.leucanthus Hance. Notably, except for R.leucanthus, the remaining five species in this subclade belong to the Sect. Rosaefolii as defined by Focke (1910–1914). Similar to the ITS phylogenetic tree results, in the rbcL + matK phylogenetic tree (Fig. 4), R.semiplenus falls within clade D1, also representing the subgenus Batothamnus, and clusters with R.hirsutus, R.rosifolius, R.illecebrosus Focke, R.croceacanthus to form subclade G1 with full support (BS = 100). All species in subclade G1 belong to Focke’s Sect. Rosaefolii. Additionally, R.semiplenus is resolved as sister to R.hirsutus with moderate support (BS = 83).
Maximum likelihood phylogenetic tree of Rubus based on ITS sequences, illustrating the phylogenetic placement of R.semiplenus. Bootstrap values are shown above branches. R.semiplenus is highlighted in bold face. Subgeneric names are listed on the right, with different subgenera represented by different colors. Subgeneric names and classifications follow the taxonomy proposed by Huang et al. (2023).
Maximum Likelihood tree of Rubus based on rbcL+matK gene sequences illustrating the phylogenetic placement of R.semiplenus. Bootstrap values are shown above branches. R.semiplenus is highlighted in bold face. Subgeneric names are listed on the right, with different subgenera represented by different colors. Subgeneric names and classifications follow the taxonomy proposed by Huang et al. (2023).
Phenology.
Flowering from March to April, Fruiting from May to June.
Etymology.
The specific epithet “semiplenus” is derived from the Greek words “semi” (meaning half) and “plenus” (meaning double), which refer to this new species with semi-double flowers.
Habitat and distribution.
Rubussemiplenus is known only from the type locality in Nanyue Hengshan National Nature Reserve, Hengyang City, Hunan Province, China. This nature reserve belongs to a subtropical monsoon humid climate and includes many Chinese National Key Protected Plants, such as Ginkgobiloba L., Taxuswallichianavar.mairei (Lemée & H. Lév.) L. K. Fu & Nan Li, and Bretschneiderasinensis Hemsl. The new species mainly grows under the shrubs at an altitude of 800–900 meters.
Discussion
The trait of doubled or semi-doubled flowers is highly valued in horticulture for its aesthetic appeal and ornamental potential, yet it remains exceptionally rare in wild plants. Within the Rosaceae, this trait is commonly observed in many cultivated species across several genera, including Prunus L. and Rosa L. (Yang et al. 2024). However, it is exceedingly rare in the genus Rubus. Prior to this study, only a few species, such as Rubusrosifoliusvar.coronarius (Sims) Focke, along with a limited number of cultivated varieties, were known to exhibit this characteristic (Mao et al. 2012). Given its rarity, the discovery of Rubussemiplenus is of significant botanical and horticultural value.
Compared to the typical shrub habit, species with subshrub or herbaceous habits are relatively rare in genus Rubus (Yu and Lu 1985). However, the emergence of subshrub habits may be the result of convergent evolution, as these species often appear in different evolutionary lineages. In China, about 40 species of Rubus have been recorded as subshrubs or herbaceous types (Yu and Lu 1985; Lu and Boufford 2003). According to the latest subgeneric classification by Huang et al. (2023), most of the subshrub or herbaceous species belong to the Subg. Lineati (Focke) T.R.Huang & X.Y.Zhu, such as R.fockeanus Kurz and R.metoensis T. T. Yu & L. T. Lu. Species within this subgenus are mostly adapted to high altitude or high latitude habitats. Morphologically, the new species should be classified into Subg. Batothamnus (Focke) E.H.L.Krause (Huang et al. 2023). Within Subg. Batothamnus, there are also some subshrub-type Rubus species, such as R.delavayi Franch., R.impressinervus F. P. Metcalf and R.simplex Focke. Nevertheless these taxa belong to different sections and have significant taxonomic differences from the new species. Molecular analyses demonstrate that although the new species clusters within the same major clade as R.leucanthus Hance and R.simplex Focke (Figs 3, 4; Clade E, Clade D1), marked discontinuities in both phylogenetic relationships and morphological characteristics are observed.
Phylogenetic analyses based on ITS and rbcL + matK sequences consistently indicate that the new species belongs to Rubussubg.Batothamnus (Focke) E.H.L. Krause. Moreover, R.semiplenus should be classified within Focke’s Sect. Rosaefolii. This result is not only supported by molecular results, but also aligns with its morphological characteristics, which are typical of species in this section, such as numerous carpels and white flowers. However, all previously known species in this section are shrubs with compound leaves. In contrast, R.semiplenus is unique, being the only species in this section with simple leaves and semi-double flowers, further distinguishing it from other members of the group.
Molecular analyses indicate that the new species is most closely related to R.hirsutus. However, it can be readily distinguished from R.hirsutus by several characteristics. The new species is a herb or dwarf subshrub, whereas R.hirsutus is a shrub. Its stems are villous, compared to the soft and glandular hairs of R.hirsutus. The leaves are simple, whereas R.hirsutus has imparipinnate leaves with 3–5 leaflets. The leaf blades are suborbicular, whereas those of R.hirsutus are ovate to broadly ovate. The petals are arranged in two cycles, with 10–12 petals, in contrast to the five petals of R.hirsutus. Additionally, the fruits are glabrous or sparsely pubescent, whereas those of R.hirsutus are entirely glabrous.
Additional specimens examined
Rubushirsutus. CHINA. Jiangxi: Fengxin County, 11 May 1963, Liu SL et al. 1169 (IBSC-0324075). Jiangsu: Nanjing City, June 1955, Guan KJ 147 (HENU-0180581). Anhui: Taiping County, at 700 m, June 1959, Herbarium of the Institute of Botany 2056 (PE-00250149). Fujian: Wuyishan City, Mount Wuyi, at 170 m, 4 April 1955, Wang MJ et al. 3003 (NAS-00365571). Henan: Xinyang City, kikungshan, at 700 m, 12 May 1998, Yang H 97053153 (HENU-1016206). Zhejiang: Hangzhou City, Changhua Town, at 650 m, 9 May 1957, He XY 33648 (IBSC-0324088).
Supplementary Material
XML Treatment for Rubus semiplenus
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alice LA Campbell CS (1999) Phylogeny of Rubus (Rosaceae) based on nuclear ribosomal DNA internal transcribed spacer region sequences.American Journal of Botany 86(1): 81–97. 10.2307/265695721680348 · doi ↗ · pubmed ↗
- 2Bailey LH (1941–1945) Species Batorum: The genus Rubus in North America I–X.Gentes Herbarum 5: 1–918.
- 3Capella-Gutiérrez S Silla-Martínez JM Gabaldón T (2009) trim Al: A tool for automated alignment trimming in large-scale phylogenetic analyses.Bioinformatics (Oxford, England)25: 1972–1973. 10.1093/bioinformatics/btp 34819505945 PMC 2712344 · doi ↗ · pubmed ↗
- 4Chen X Wang SY (2006) The resources research and to exploit using of Chinese bramble (Rubus L.). Southwest China Journal of Agricultural Sciences 19(Z 1): 351–355.
- 5Chen M Lin GC Wang T Zhuang YX Yao YX Yang CZ Qin Y Lin YX An C (2024 a) Rubustingzhouensis (Rosaceae), a new species from Fujian, China.Phyto Keys 249: 251–267. 10.3897/phytokeys.249.13895139659964 PMC 11629084 · doi ↗ · pubmed ↗
- 6Chen PH Huang JY Chung AC (2024 b) Rubuspuyumaensis, a new species of Rosaceae from Taiwan. Biodiversity Data Journal 12: e 115431. 10.3897/BDJ.12.e 115431 PMC 1083571138314125 · doi ↗ · pubmed ↗
- 7Davis PH Heywood VH (1963) Principles of Angiosperm Taxonomy.Oliver and Boyd, Edinburgh, 558 pp.
- 8Focke WO (1910) Species Ruborum. Monographiae generis Rubi prodromus I. Bibliotheca Botanica 17 (Heft 72): 1–120. 10.5962/bhl.title.15533 · doi ↗