An integrative approach to alpha taxonomy in Erica L. (Ericaceae) with three new species from the Western Cape, South Africa

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
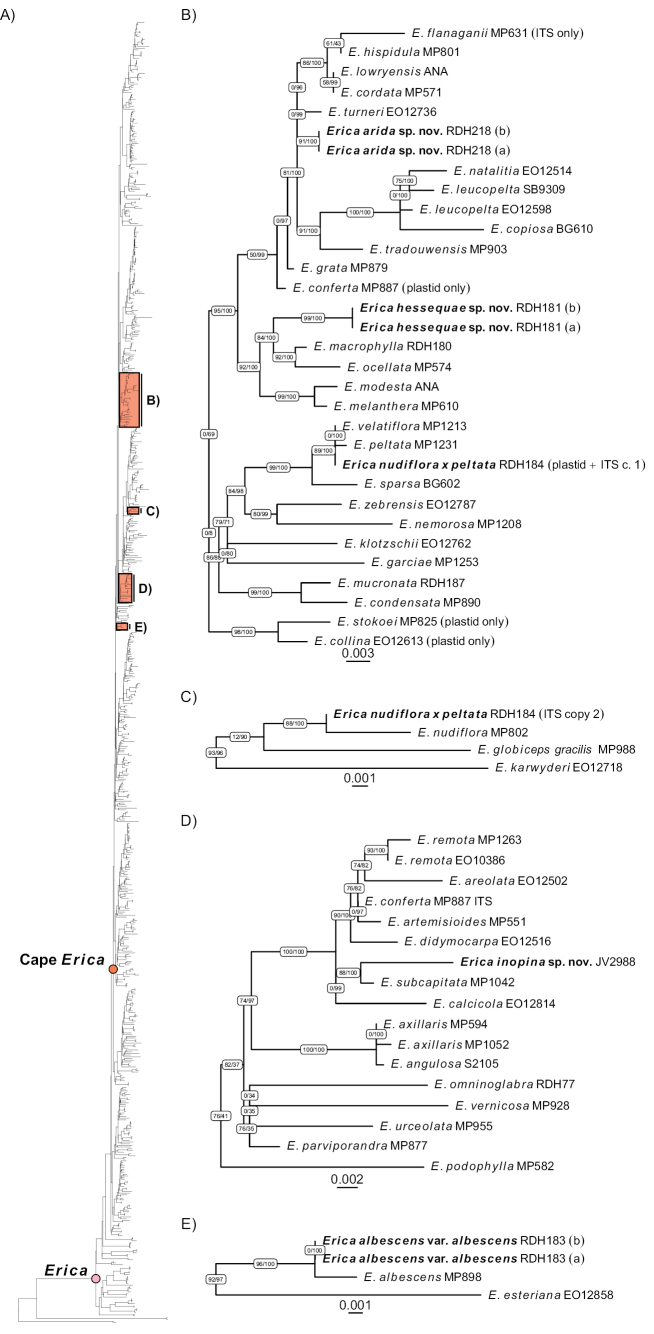 Figure 1
Figure 1 Figure 2
Figure 2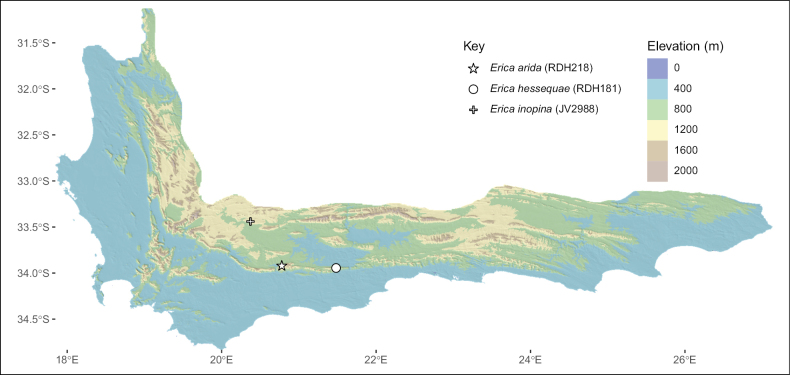 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6| Character | |||||
|---|---|---|---|---|---|
| Corolla Size (mm) | 1–5 | 1–5 OR 5–10 | 1–5 | 1–5 | 1–5 |
| Corolla Shape | Bell OR Cup OR Urn | Bell OR Cup | Bell OR Cup | Bell OR Cup | Urn OR Cup |
| Corolla Colour | White | White | Pink OR Red | White | Green OR Yellow OR Red |
| Sepal/Corolla Length Ratio | < ½ OR >½ | < ½ | < ½ OR > ½ | < ½ | < ½ |
| Hairy structures | Stem AND Leaf AND Pedicel AND Sepal AND Ovary | Leaf AND Pedicel AND Sepal AND Corolla AND Ovary | Stem AND Leaf AND Pedicel AND Sepal AND Ovary | Ovary | Stem |
| Non-Hairy structures | Corolla | - | Corolla | Stem AND Pedicel AND Corolla | Leaf AND Pedicel AND Sepal AND Corolla AND Ovary |
| Style | Exserted | - | Exserted | Exserted | Exserted |
| Anthers | - | Non exserted | Non exserted | Non exserted | - |
| Anther appendages | Present | Present | Absent | Absent | Absent |
| Stamens (no.) | 8 | 8 | 8 | 8 | 6 |
| Sepals (no.) | 4 | 4 | 4 | 4 | 4 |
| Corolla lobes (no.) | 4 | 4 | 4 | 4 | 4 |
| Leaves (n-nate) | 3 | 3 | 3 | 3 | 3 |
| Bracts and bracteoles (no.) | 3 | 3 | 3 | 3 | 3 |
| Fire survival | Reseeder | Reseeder | Reseeder | Reseeder | Reseeder |
| Flowering month | December OR January | January OR February | January OR February | December OR January OR February | April OR May OR June |
| Region | Langeberg | Langeberg | Agulhas | Langeberg | Karoo Mountains |
| Match function | |||||
| Region and flowering month excluded (no.) | (0) | ||||
| Region included and flowering month excluded (no.) | (0) | ||||
| All characters included (no.) | (0) | (0) | |||
| Probability function | |||||
| Region and flowering month excluded | |||||
| Region included and flowering month excluded | |||||
| All characters included | |||||
|
|
| ||
|---|---|---|---|
| Branches | pubescent | glabrous | pubescent |
| Leaves | 3-nate | 3-nate | 3-nate |
| 4–10 mm long | 2.5–5.0 mm long | 2.0–3.5 mm long | |
| lanceolate, acute | lanceolate, acute | lanceolate, acute | |
| glabrous to hispidulous | mostly glabrous | mostly glabrous | |
| Flowers | pseudo-axillary on reduced side branches | terminal on short side branches | terminal at the ends of branchlets |
| Pedicels | 2.5–5.5 mm long | 2.0–3.0 mm long | ± 1 mm long |
| puberulous | pubescent | puberulous | |
| Bracts and bracteoles | 3 | 3 | 0–3 |
| remote | subapproximate | variable, bracteoles usually subremote | |
| Ciliolate | |||
| Calyx | 1:4 to 1:2 to corolla | 1:2 to corolla | 1:2–1:1 to corolla |
| lobes lanceolate, acute | lobes lanceolate, acute | lobes ovate, subacute to obtuse | |
| mostly glabrous, | |||
| Corolla | subcampanulate- | cyathiform | broad cyathiform |
| cyathiform | pink to red | pink | |
| pink to red | glabrous | glabrous | |
| glabrous | 2.0–2.5 mm long | 1.5–2.5 mm long | |
| 2.5–5 mm long | |||
| Anthers | exserted | included | manifest to exserted |
| scabrous | scabrous | scabrid | |
| muticous | muticous | muticous | |
| pores ¼–1/3 of anther | pores ½ of anther | pores 2/3 of anther | |
| Ovary | glabrous | pubescent | pubescent |
| Style | exserted | exserted | exserted |
| simple | simple | peltate | |
| Nectaries | present | present | absent |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Diversity and Evolution · Plant and animal studies · Plant Parasitism and Resistance
Introduction
The megadiverse genus Erica L. (Ericaceae) forms an extraordinary focus of diversity in the Cape Floristic Region (CFR) of South Africa, where nearly 700 of its 851 species are found, almost all of which are regionally endemic (Elliott et al. 2024). This reflects the overall high levels of diversity and endemism in the CFR (Goldblatt and Manning 2000; Linder, 2003). Its sheer size and taxonomic complexity make Erica challenging to work with. It has been the subject of substantial taxonomic work, from the foundational treatment of the South African species by Guthrie and Bolus in Flora Capensis (1905) and the comprehensive revision of Dulfer (1964, 1965), to works focused on particular regions and groups (Oliver 2000; Oliver and Oliver 2002, 2005; Beentje 2006; Nelson 2011). However, these represent fragmented and in part outdated taxonomic accounts. Morphological variation in Erica is complex, and the task of distinguishing known and unknown diversity has become increasingly challenging. Undescribed taxa may be effectively hidden within species complexes that have only recently begun to be resolved with the aid of molecular studies (e.g. Pirie et al. 2017; Musker et al. 2024).
Many species of Erica are infrequently encountered or collected because of their natural rarity, ongoing decline in species populations, and tendency to occur in remote and inaccessible mountainous areas. Many are also highly localised endemics, some restricted to areas as small as a few hundred m^2^, and many of these, such as E.burchelliana E.G.H.Oliv., E.chrysocodon Guthrie & Bolus, E.extrusa Compton, E.feminarum E.G.H.Oliv., E.gerhardii E.G.H.Oliv. & I.M.Oliv., E.heleogena T.M.Salter, E.hillburttii (E.G.H.Oliv.) E.G.H.Oliv., E.karwyderi E.G.H.Oliv., E.petrusiana E.G.H.Oliv. & I.M.Oliv., E.perplexa E.G.H.Oliv., E.remota (N.E.Br.) E.G.H.Oliv., and E.sociorum L.Bolus, are critically endangered (Raimondo et al. 2009). Their specialised niches and highly restricted geographical distributions place them at high risk of extinction. According to the International Union for Conservation of Nature (IUCN) Red List definitions, more than 150 species and subspecies can be considered threatened (Raimondo et al. 2009); however, most assessments fall outside the 10-year threshold specified by IUCN, they leave many as Data Deficient or unassessed, and they cannot address undescribed species’ diversity (Pirie et al. 2024). The latter is particularly critical as undescribed taxa are more likely to be threatened with extinction in the wild (Brown et al. 2023). Our research, and specimens collected and curated by the pre-eminent expert on the genus E.G.H. Oliver (Nelson et al. 2024) prior to the cessation of his active work, point to a significant remaining shortfall in the documentation of Erica alpha diversity. New species descriptions need to be prioritised as the first step towards justifying targeted conservation of these taxa.
Most of the taxonomic work on Cape Erica has been limited to the prolific output of few, largely successively working specialists, with the majority of recent work having been performed over much of the 20^th^ and into the 21^st^ century by the South African botanists E.G.H. and I.M. Oliver (Nelson et al. 2024). Sadly, they are no longer with us, and in the absence of a modern, consolidated revision it is hard even for experts to identify novelties amongst the enormous species diversity of a complex group like Erica. Fortunately, an overview of accepted taxa and variation in morphological characters is provided by ‘Genus Erica: an Identification Aid (the Erica ID aid; Oliver et al. 2024a) Version 4.00.02 (https://doi.org/10.5281/zenodo.10651955). The Erica ID aid offers a means by which to identify the most similar taxa to a given specimen, even when it may represent an undescribed taxon, making this tool an accessible means to aid species identification and guide alpha taxonomy.
A further resource for comparing known variation is the body of published DNA sequence data, which represents over 65% of the recognised species of Erica (Pirie et al. 2024). Despite limited resolution of phylogenetic relationships, particularly within the Cape clade, a researcher may be able to identify the closest relatives of putative new species or to discern them from morphologically similar distant relatives with the use of standard protocols and widely used DNA sequence regions such as the nuclear ribosomal Internal Transcribed Spacers (ITS) and plastid trnF-ndhJ intergenic spacer.
In this paper, we assess five examples of apparently undescribed variation in Erica. In each case, we used the Erica ID aid to generate lists of most similar known taxa. To test the relatedness of the putative new taxa, we collected and analysed new DNA sequence data, including data from new collections of similar and potentially related known taxa. We then assessed potential diagnostic differences between focal specimens and known taxa, concurrently implementing improvements to the Erica ID aid. Where the results support the recognition of new taxa, we describe them formally. Using this combined approach, we describe three new Erica species from the Eastern Langeberg and Karoo Mountain regions in the Western Cape, South Africa, presenting diagnoses and descriptions, as well as distributions, photos and threat status.
Materials and methods
Field research and collection
Collections of potentially undescribed taxa were made in the field under research permits from Cape Nature (CN35-28-27705) and permission from private landowners. Herbarium vouchers were lodged at NBG with some duplicates at BG (acronyms after Thiers 2018), and fresh leaf samples were dried in silica gel for DNA extraction. Details of habitat and ecology were recorded in the field and, where possible, photographic records were taken of specimens in their habitat and of floral and vegetative parts. Limited surveys were performed in surrounding localities to assess the extent of distributions. Additional targeted surveys were conducted to locate further subpopulations. We used GeoCAT (https://geocat.iucnredlist.org) to calculate an Extent of Occurrence (EOO) and Area of Occupancy (AOO) for each taxon.
Morphological comparisons using the Erica ID aid
We performed character-based searches using the Erica ID aid. The key features a strict “Match” and a more flexible “Probability” algorithm for narrowing down species identifications. The “Match” algorithm provides a list of only those taxa with characteristics that are consistent with those selected, while considering intra-specific character variability. For example, under the character “Corolla Size”, when “1–5 mm” and “5–10 mm” are selected, all species which have corollas of size 1–5 mm and/or 5–10 mm are possible identifications. When we processed species using this feature, results were recorded for three scenarios: “region and flowering month excluded”; “region included and flowering month excluded”, and “all characters included”. This was done to further minimise errors as it accounted for variability in geographic range, which might have been underestimated in previous records of recognised species, and flowering times, which might similarly be longer than previously recognised or have changed over time.
The “Probability” algorithm provides a complete list of taxa ordered by probability, whereby rarer characteristics impart higher probabilities than more common ones and narrow variation imparts higher probabilities than wide. The most probable results can therefore include taxa that are excluded altogether when using the strict “Match” algorithm and strict matches may score as less probable if characters are coded to multiple states (Oliver et al. 2024b). Both sets of results can be reproduced with the same character inputs presented in the Results with the same (permanently archived) version of the Erica ID aid.
Identifying closely related taxa using phylogenetic inference
To identify close relatives of the putative new taxa and potentially exclude morphologically similar but distantly related species from considerations of species boundaries, we sequenced ITS and trnF-ndhJ, markers used in the most comprehensive phylogenetic analyses of Erica (Pirie et al. 2016, 2024) and reanalysed the available data. We rarely had access to multiple accessions of the same taxon; where only one individual was found, we sequenced two samples of the same individual that had been collected on separate occasions to confirm the result. DNA was isolated with the DNeasy Plant Mini Kit (Qiagen, Hilden, Germany) followed by PCR amplification with universal angiosperm primers (Taberlet et al. 1991; Shaw et al. 2007) as in Mugrabi de Kuppler et al. (2015).
In most cases, Sanger sequencing delivered unambiguously interpretable sequences. In one case (Erica sp. RDH184), the resulting ITS chromatograms showed clear double peaks of similar heights at 12 informative sites. Plastid data indicated a sister group relationship to the morphologically similar species E.peltata Andrews (see Results). At each polymorphic site in the ITS sequence we observed that one of the two overlapping peaks corresponded to the same base observed in E.peltata. Under the assumption that this pattern reflected inheritance from one of two distinct parental species, we recreated two separate ITS sequences, one with E.peltata base variants and the other with the unknown alternative variants.
New sequences were added manually to the alignments of ITS and of trnF-ndhJ sequences in Pirie et al. (2024) and analysed separately to assess supported phylogenetic conflict before (in the absence of such conflict) being combined in a supermatrix of the complete nrDNA and plastid sequence data. For Erica sp. RDH184, the one ITS sequence corresponding to E.peltata was combined with the plastid data whilst the other was included as a separate operational taxonomic unit in the combined analyses, as in the approach of Pirie et al. (2009).
We performed phylogenetic analyses using IQ-TREE v. 2.3.4, with automatic partition merging and model selection, followed by 3,000 iterations of tree searching (Chernomor et al. 2016; Kalyaanamoorthy et al. 2017; Minh et al. 2020). We evaluated branch support using 10,000 replicates of Ultrafast Bootstrap (Hoang et al. 2018) and SH-alrt (Guindon et al. 2010).
Morphological comparisons with herbarium records
Material in the BOL and NBG herbaria (Thiers 2018) was examined, both of morphologically similar and closely related taxa, as identified above, to find additional collections of each putative novel taxon. Potentially diagnostic characters for putative new taxa were compared to the morphologically similar and closely related species.
Results
Field research and collection
We made collections of 10 specimens representing five unidentified and/or potentially undescribed species and five additional known taxa not previously sequenced. Observations were recorded on the open platform iNaturalist (https://www.inaturalist.org). Collection and observation data, along with GenBank accession numbers, were uploaded to GBIF (Suppl. material 1; https://doi.org/10.15468/abecqd).
Morphological comparisons using the Erica ID aid
We found little overlap between taxon lists generated from the strict matching versus highest-scoring taxa probabilistic analyses (Tables 1, 2; Suppl. material 2). In some cases, there was no strict match at all; in others, taxa that are consistent with the coding of the putative novelties but more variable were recovered with lower probability. Adding distribution and phenological data to the analysis dramatically reduced options for similar taxa.
Identifying closely related taxa using phylogenetic inference
In most cases, phylogenetic analyses of the separate ITS and trnF-ndhJ sequence alignments yielded consistent results (except for the two ITS copies in Erica sp. RDH184) and thus the data from both partitions were combined into a single analysis (Fig. 1; Suppl. material 3). The following are the closest related taxa for the focal putative novel taxa based on ITS and plastid data:
Selected clades from the phylogenetic analysis of combined plastid and nuclear sequence data illustrating the relatedness of the focal specimens analysed here. The complete phylogeny is shown in A and the positions of the focal clades (B–D) are highlighted.
For Erica sp. RDH184, the plastid phylogeny placed this sample in a well-supported clade (ultrafast bootstrap support [BS] = 100%; SH-alrt support [AS] = 90%) along with E.peltata, E.sparsa G.Sinclair, and E.velatiflora E.G.H.Oliver (Suppl. material 3: fig. S1). The two copies of ITS “phased” manually from the sequence chromatograms showed contrasting relationships in the nuclear phylogeny (Suppl. material 3: fig. S2). As expected, the copy that was phased based on its close match to E.peltata (Copy 1) was placed with good support (BS = 100%, AS = 97%) in a clade with E.peltata and E.velatiflora, while the other copy (Copy 2) was well supported (BS = 100% AS = 83%) as sister to E.nudiflora L.. Based on the concordance between the plastid and nuclear results, we concatenated Copy 1 with the plastid data for the combined nuclear and plastid analysis, while retaining Copy 2 separately. The combined analysis showed a pattern consistent with the plastid and nuclear phylogenies, with the “primary” RDH184 sequence grouping with E.peltata and E.velatiflora (BS = 100%, AS = 89%; Fig. 1B) and the “secondary” (alternative ITS-only) copy grouping with E.nudiflora (BS = 100%, AS = 88%; Fig. 1C).
For Erica sp. RDH181, the nuclear phylogeny suggested a well-supported (BS = 100%, AS = 91%) sister relationship with E.ocellata Guthrie & Bolus and E.macrophylla Klotzsch ex Benth. (Suppl. material 3: fig. S2). In contrast, the plastid phylogeny supported a sister relationship with E.grata Guthrie & Bolus (BS = 100%, AS = 87%), although the placement of this pair was poorly resolved (BS = 51%, AS = 0%) within a broader well-supported (BS = 99%, AS = 88%) clade including, among several others, E.macrophylla and E.ocellata (Suppl. material 3: fig. S1). The combined analysis agreed with the nuclear phylogeny, placing RDH181 in a clade with E.macrophylla and E.ocellata (BS = 100%, AS = 84%; Fig. 1B).
In all three analyses, Erica sp. RDH218 was included in a clade with, among others, E.grata, E.tradouwensis Compton, E.turneri E.G.H.Oliv., E.cordata Andrews, and E.hispidula L., although this grouping received variable support depending on the analysis (nuclear: BS = 100%, AS = 90%; plastid: BS = 51%, AS = 0%; combined: BS = 97%, AS = 0%; Fig. 1B; Suppl. material 3: figs S1, S2).
The placement of Erica sp. JV2988 was not resolved by the plastid data (Suppl. material 3: fig. S1), but the nuclear and combined analyses indicated a well-supported (nuclear: BS = 100%, AS = 89%; combined: BS = 100%, AS = 88%) close relationship to E.subcapitata (N.E.Br.) E.G.H.Oliv., the two forming a sister pair nested in a clade including E.calcicola (E.G.H.Oliv.) E.G.H.Oliv., E.areolata (N.E.Br.) E.G.H.Oliv., E.artemisioides (Klotzsch) E.G.H.Oliv., E.conferta Andrews, E.didymocarpa E.C.Nelson & E.G.H.Oliv., and E.remota (N.E.Br.) E.G.H.Oliv., which had strong support (nuclear: BS = 100%, AS = 97%; combined: BS = 100%, AS = 100%; Fig. 1D; Suppl. material 3: fig. S2).
Erica sp. RDH183 was placed as sister to E.albescens Klotzsch ex Benth. (nuclear: BS = 100%, AS = 87%; plastid: BS = 100%, AS = 93%; combined: BS = 100%, AS = 96%; Suppl. material 3: figs S1, S2, S3).
Discussion
In this work we focused on five apparently distinct Cape ericas potentially representing new species. In three cases we were able to compare the specimens to both the most similar species and to their closest relatives (insofar known) and show that they can be clearly distinguished and warrant describing as new (see taxonomic treatment below). In the two other cases, the approach allowed us to better explain the variation observed and we were able to rule out the necessity for describing new taxa.
In the case of Erica sp. RDH184, the molecular evidence indicated that a morphologically distinct individual represents a (naturally occurring) hybrid. The data represents two independent gene trees: that of the generally maternally inherited plastid and biparentally inherited nuclear ribosomal genes. The latter can be problematic for phylogenetic inference because they are present in numerous copies in the genome, including potentially paralogous copies (Baldwin et al. 1995). However, in Erica this rarely appears to present a problem (several exceptions are listed in Pirie et al. 2024), and in this case it was possible to discern two copies with apparently comparable copy numbers but distinct phylogenetic signals: one consistent with the plastid data, indicating a maternal inheritance from E.peltata (or potentially an unsampled close relative), the other more similar to E.nudiflora. During character-based searches with the Erica ID aid, E.peltata was included in the results for the “Match” algorithm and E.nudiflora appeared in all three searches with the “Probability” algorithm where region was included as a character. Both putative parent species are known from the area and there are clear morphological similarities (see Fig. 2, Table 3 for comparisons). Additionally, only one individual like Erica sp. RDH184 has been found despite exhaustive surveys in the area. The data therefore provide strong evidence that it is an F_1_ hybrid. This result contributes to our knowledge of reproductive barriers amongst Erica species and highlights how an integrative approach could avoid artificially inflating the numbers of true species with morphologically distinct hybrids.
Morphological comparison between Ericanudiflora (top and bottom left), E. sp. RDH184 (top and bottom middle) and E.peltata (top and bottom right).
For Erica sp. RDH183, the results pointed to a match with an existing species whilst identifying a need to update the character coding within the Erica ID aid. Phylogenetic analysis of this specimen showed a close relationship to a specimen (MP898) collected in the nearby area of Boosmansbos identified by E.G.H. Oliver as E.albescens. Morphologically, both specimens appear to correspond to E.albescensvaralbescens whilst deviating in flower shape coded for the species in the Erica ID aid, thereby excluding a strict match. However, using the “Probability” algorithm, E.albescens was recovered as the most likely species once the region was included as a character (see Table 2). The simplest conclusion from these results is to determine the specimen as E.albescensvaralbescens and to update the Erica ID aid accordingly.
In general, the results highlighted the importance for identification and species matching of factors such as geographic distribution and flowering time. These are clearly relevant for species boundaries in the context of the patterns of narrow endemism and shifts between pollinators within the Cape (Van der Niet et al. 2014; Musker et al. 2024). The differences between results based on strict and probabilistic matching are consistent with the different interpretations of character variation they imply. Otherwise similar taxa might be unambiguously distinguished with strict matching. As an example, Ericapodophylla is the second-most probable match for Erica sp. RDH181, yet it does not feature in the strict matching due to its hairy corolla (non-hairy in Erica sp. RDH181). For Erica sp. JV2988 there is no strict match at all, and probabilistic matching suggests comparison to E.atricha with the more common 8 stamens (as opposed to 6 stamens as in Erica sp. JV2988). The possibilities for unrecorded intraspecific variation and for simple error clearly support the use of both algorithms when attempting species identifications.
The new samples of known taxa and of newly discovered species bring the total representation of species diversity in the phylogenetic tree of Erica to around 66% of 854 species, as compared to 65% of 851 species in Pirie et al. (2024). Further increases in sampling will improve the power of phylogenetic comparisons in discovering and assessing putative species diversity in Erica. Testing results will provide further improvement, particularly when they are derived from single samples of specific taxa or if they seem unexpected from the morphologies and distributions of apparently closely related taxa (such as that of the ITS sequence of E.flanaganii Bolus MP631). Phylogenetic analysis of Erica taxa is mostly based on only a few Sanger-sequenced markers. As in the cases illustrated here, these markers can still be informative despite their limited variability and representation of genomes; however, they will not always deliver a conclusive result. High-throughput sequencing and phylogenomic approaches (Musker et al. 2024, 2025) may be needed to inform taxonomic decisions, particularly in species complexes.
Subgeneric classification is another area in which further research is needed. The classification used by Guthrie and Bolus (1905) to group similar species into sections has been maintained in subsequent work and adopted on iNaturalist. However, the sections are not monophyletic, often grouping distantly related taxa (Pirie et al. 2011, 2016). They cannot be assumed to have predictive value for combinations of traits in either known or undescribed taxa. This has made the attempt to place new species within the existing classification challenging and, arguably, arbitrary. Thus, the subgeneric placement of new species and the current classification should not be over-interpreted. Nonetheless, our hope is that the attempt will be useful as a bridge towards a more robust classification of Erica.
Taxonomic treatment
This section provides a taxonomic treatment for three new Erica taxa from the Western Cape, South Africa. As far as possible, taxonomic treatments were standardised in accordance with the methodology of E.G.H. Oliver, with minor morphological details maintained as subtle but key distinguishing characters in this large and complex genus. The treatment for E.inopina is reduced to follow Oliver’s (2000) methodology for species with indehiscent and partially dehiscent fruit, which includes less detail. Diagnoses are included above the descriptions as in the recommendations of Hassemer et al. (2020). We distinguish the new taxa from those that present a strict match according to the Erica ID aid without reiterating characters in Table 1; in one case without a strict match we report the characters that differ from the closest known relative. Further discussion of similar taxa, including close relatives, identification, and more variable characters (such as habit) are included under subsequent notes. Specimen localities are based on the quarter-degree grid cell system employed by the South African National Mapping Agency, National Geo-Spatial Information (2017), e.g. “3321CD”, in addition to precise coordinates. We also provide summary descriptions (Suppl. material 4) and geographic distributions (Fig. 3).
Map of the Cape Floristic Region, showing the distributions of the new taxa described in this study.
Erica
arida
Taxon classificationPlantaeEricalesEricaceae
R.D.Hoekstra sp. nov.
9651A820-52D4-5567-98C1-372F5A75DC57
urn:lsid:ipni.org:names:77362792-1
Link.
WFO: https://list.worldfloraonline.org/wfo-1000079209.
Type.
South Africa • Western Cape, 3320DD (Warmwaterberg): Langeberg Range, Barrydale outskirts, on steep middle south-facing slopes north of the Doringrivier Catchment Area, 709 m, 33.922°S, 20.779°E, 14 February 2024, R.D. Hoekstra 218 (NBG, holotype).
Diagnosis.
Ericaarida matches characters exhibited in E.tenuicaulis, from which it differs in leaf shape (saddle-shaped, as opposed to broadly linear in E.tenuicaulis), its straight style (curved in E.tenuicaulis) and staminal filaments approximately 3 times as long as the anthers (as long as the anthers in E.tenuicaulis).
Description.
Rounded to semi-spreading, resprouting shrublet up to 30 cm tall, rootstock thick. Branches twiggy, erect, glabrous; secondary branches erect or ascending, sparsely stipitate-glandular. Leaves 3-nate, erect to spreading, ovate to subcordate, saddle-shaped, blades 2.5–4.0 × 1.0–1.5 mm, acute; adaxially convex, glabrous, margins revolute, rarely sparsely ciliolate; abaxially deeply sulcate, densely hispidulous within sulcus with simple, eglandular hairs; petiole ± 1 mm long, occasionally loosely decurrent, margins ciliolate. Inflorescence of 3-nate flowers at ends of short side branches; pedicel 3.0–4.0 mm long, pale green to pink, viscid, sessile- and stipitate-glandular; bracteoles 2, opposite, median to submedian, oblong to shortly lanceolate, ± 0.75 mm long, subacute, sessile-glandular, glabrous but margins sparsely ciliolate at base, green and slightly sulcate towards abaxial apex; bract partially recaulescent, submedian, lanceolate, acute, sessile-glandular, margin sparsely ciliolate towards base. Calyx 4-lobed, sepals connate at base, lanceolate, 1.25–1.5 mm long, acute; abaxially glabrous, apex sulcate, the sulcus shortly villous; adaxially glabrous; margins sessile-glandular. Corolla campanulate to shortly urceolate with slightly constricted throat, 3.0–4.0 × 3.0–4.0 mm, white, glabrous, dry; lobes erect to spreading, ± 0.5 mm long, acute, minutely serrated. Stamens 8, free, included, filaments flat, ± 2.5 mm long, glabrous, white turning golden reddish distally; anthers ± 0.8 mm long, obcuneate, dorsifixed at base, bipartite, thecae erect, free, muticous, dark brown; pore ± 0.3 mm long, oval. Ovary 4-locular, shortly oblong, ± 0.8 mm long, slightly emarginate, red to purplish, hispid with long, simple, eglandular hairs; ovules ± 20 per locule; placenta apical; nectaries basal, green; style ± 3.5 mm long, glabrous, pale pink turning dark red or black towards stigma, exserted; stigma dark red to black, subcapitate. Fruit and seeds not seen. Flowering time: December to February.
Ericaarida R.D.Hoekstra A incomplete inflorescences B glandular pedicel, bracteoles and calyx C dissected flower showing parts of the androecium and gynoecium D whole plant in situ. Voucher R.D. Hoekstra 218 (NBG).
Distribution and habitat.
Ericaarida is known only from the mountains north of the Doringrivier Catchment Area east of Barrydale in the Western Cape. It has been recorded from a ridgeline at an altitude of 1190 m in Northern Langeberg Sandstone Fynbos (Mucina and Rutherford 2006), growing in association with Ericabarrydalensis, as well as on the middle south-facing slopes as low as 516 m growing in association with Phylicamairei, Ericavestita and Cliffortiapulchella. It has not been recorded from the north-facing slopes.
Threat status.
Ericaarida is only known from a few records on iNaturalist and our single collection from the Doringrivier Catchment Area. The three known subpopulations appear to be small and scattered within the catchment area, and our survey at the type locality revealed only 17 plants. Ericaarida was found to have an AOO of 12.00 km^2^ and an EOO of 12.00 km^2.^ Additionally, the marshland in the catchment area is heavily infested with invasive Hakeasericea with a significant encroachment up to the middle south-facing slopes where E.arida occurs. This poses a major threat to the two subpopulations on the middle slopes. The habitat of E.arida is remote and difficult to access and only limited botanical surveys have been performed around the catchment area. Therefore, there are not enough data to accurately estimate the population size of this species. However, the wide range in altitude between the known subpopulations and results from our survey suggest that additional small subpopulations may exist within this catchment area. Since all observed subpopulations have consisted of only a few plants we estimate a minimum total population of 100 plants. Based on this estimate, an EOO of 12.00 km^2^, ongoing decline in quality of habitat as a result of invasive alien plant species, and its restriction to a single known location, we recommend an IUCN (2012) category of Critically Endangered (CR) under criterion B1ab(iii) for E.arida.
Pollination syndrome.
Ericaarida has well-developed nectaries and small, open cup-shaped flowers, suggesting entomophily. The lack of a peltate stigma and the fact that pollen is not released in a cloud after in situ manipulation make anemophily highly unlikely, and the size and shape of the corolla make ornithophily unlikely.
Etymology.
Ericaarida is named for the seasonal aridity seen during its flowering months in the part of the Klein Karoo where it occurs; from the Latin, arida, for dry or arid.
Subgeneric classification.
The closest relatives of E.arida are classified in at least five different (non-monophyletic) sections, but most are in either Ceramia (glabrous, pitcher-shaped corollas and terminal inflorescences: consistent with E.arida) or Arsace (large peltate stigmas: inconsistent). Placement of the new species in Ceramia may be a useful provisional classification.
Notes on morphology and phylogeny.
The single strict match for E.arida with the Erica ID aid, E.tenuicaulis, diagnosed above, has yet to be analysed phylogenetically. Phylogenetic data place E.arida in a clade containing E.cordata, E.lowryensis, E.flanaganii (ITS only), E.hispidula, E.turneri, E.leucopelta, E.natalitia, E.copiosa, E.tradouwensis and E.grata, with which it is unlikely to be confused as it differs from all of these by its glabrous (as opposed to hairy) leaves. It further differs from E.cordata, E.lowryensis and E.tenuicaulis in leaf shape (saddle-shaped, as opposed to cordate in E.cordata, and lanceolate in E.lowryensis); from E.turneri and E.copiosa by its muticous (as opposed to awned) anthers; from E.turneri by its closed-backed (versus distinctly broad, open-backed) leaves, and glabrous (versus hairy) corolla, and from E.copiosa, E.tradouwensis and E.grata by its glabrous (as opposed to hairy) sepals. Ericahispidula, E.leucopelta and E.natalitia are clearly distinct wind-pollinated plants with peltate stigmas and absent nectaries, in contrast to E.arida which has a subcapitate stigma and conspicuous nectaries.
The placement of Ericaflanaganii is unexpected: it is geographically distant, from the Drakensberg Mountains more than 500 km from the eastern edge of the CFR, as well as morphologically dissimilar, possessing a hairy corolla, lanceolate imbricate leaves, a fully recaulescent bract and anther appendages. The result (from data presented by Pirie et al. 2011; 2024) is only shown by ITS and contradicted by plastid results. Additional data from different samples are needed.
Erica
hessequae
Taxon classificationPlantaeEricalesEricaceae
R.D.Hoekstra sp. nov.
345EC13E-C5A9-57F3-B393-93AA265D260F
urn:lsid:ipni.org:names:77362793-1
Link.
WFO: https://list.worldfloraonline.org/wfo-1000079210.
Type.
South Africa • Western Cape, 3321CD (Sandkraal): Langeberg Range, Romanskraal north of Albertinia, north-facing slopes west of Skoorsteen Peak, 1165 m, 33.945°S, 21.482°E, 23 December 2023, R.D. Hoekstra 181 (NBG, holotype; BG, isotype [2088580]).
Diagnosis.
Ericahessequae matches characteristics exhibited in E.hispidula, E.oakesiorum, E.rivularis, E.tegetiformis, E.umbratica and E.woodii. It can be distinguished from all these taxa by its ovate leaves (linear, linear-lanceolate or lanceolate in all). It is further distinguished by its glandular pedicel, sepals, bract and bracteoles (all eglandular in E.magistrati, E.oakesiorum and E.tegetiformis); by its subcapitate stigma (peltate in E.hispidula; simple in E.rivularis and E.tegetiformis), and by its densely lanate ovary (shortly hispid in E.hispidula, E.oakesiorum, E.rivularis, E.woodii and E.umbratica).
Description.
Semi-spreading shrublet up to 80 cm tall. Branches twiggy, glabrous to puberulous; secondary branches ascending, hispid with a mix of simple, eglandular hairs and stipitate glands. Leaves 3-nate, erect, ovate, blades 4.0–5.0 × 1.5–2.5 mm, acute, open-backed; adaxially slightly convex, hispidulous when young with simple, eglandular hairs mixed with stipitate and sessile glands, glabrescent, margins thickened, revolute; abaxially deeply sulcate, pale, densely hispidulous with simple, eglandular hairs; petiole 0.8–1.2 mm long, hispidulous and glandular as for adaxial leaf surface, not decurrent. Inflorescence of 3-nate flowers, terminal on secondary and side branches; pedicel 4.0–5.5 mm long, pale creamy green turning red-purplish with exposure to sun, stipitate-glandular, viscid; bracteoles 2, median, lanceolate, ± 1.5 × 0.5 mm, acute, creamy white to dark pink, leathery, margins densely sessile-glandular, abaxially sparsely hispid towards apex and along basal margin, adaxially glabrous; bract partially recaulescent, sub-median, ovate to lanceolate, ± 0.5 × 1.5 mm long, acute, creamy white to dark pink, leathery, margins densely sessile-glandular, abaxially sparsely hispid towards apex and along basal margin, adaxially glabrous. Calyx 4-lobed; sepals adpressed to corolla, ovate, 2.0–2.5 × 1.0–1.5 mm, creamy white/yellow with green tips, leathery; margins densely sessile-glandular, revolute towards apex; abaxially sulcate, with occasional stalked glands and sparsely hispidulous with simple, eglandular hairs within sulcus and at base, otherwise glabrous, viscid; adaxially glabrous, midrib slightly raised. Corolla 4-lobed, cyathiform to shortly urceolate, throat constricted, 3.5–4.5 × 3.0–4.0 mm, creamy white, glabrous, viscid, occasionally with stalked glands adjacent to sepals; lobes recurved, 0.75–1.25 mm long, obtuse, margins smooth. Stamens 8, free, manifest to exserted, filaments flat, 2.5–3.0 mm long, glabrous, white turning dark reddish pink towards apex, apically kyphotic; anthers cuneate, 0.7–0.8 mm long, brown, dorsifixed at base, bipartite, thecae erect, ventral surface golden brown and shortly scabrous; awns ± 0.4 mm long, thin, laterally fixed to apex of filaments, simple or rarely with one or two basal barbs, reddish brown; pores round, ± 0.4 mm long. Ovary 4-locular, turbinate, ± 1.0 mm long, dark purple, densely lanate with simple, white, eglandular hairs; ovules 15–20 per locule; placenta apical; nectaries basal, green to black; style ± 3.5 mm long, glabrous, pale pink above but white towards base, exserted; stigma dark pink to purplish, subcapitate. Fruit and seeds not seen. Flowering time: December to January.
Ericahessequae R.D.Hoekstra A multiple inflorescences B single inflorescence C calyx and pedicel D dissected flower showing androecium E dissected flower showing ovary F whole plant in situ. Voucher R.D. Hoekstra 181 (NBG).
Distribution and habitat.
Ericahessequae is only known from two localities, some 4 km apart, on the mountains surrounding Romanskraal north of Albertinia—one along the ridgeline leading up to a plateau mountain west of Skoorsteen Peak, the other on Langeberg Peak. It appears to be confined to high-elevation ridgelines where it grows amongst craggy rocks in North and South Langeberg Sandstone Fynbos (Mucina and Rutherford 2006). It has only been observed at altitudes above 1100 m.
Threat status.
Ericahessequae appears to be a naturally rare, range-restricted species. There are only two records of the species, one plant having been observed on Langeberg Peak and two west of Skoorsteen Peak. Limited surveys performed in the surrounding area have failed to reveal additional localities. While alien vegetation is present in the region, favouring middle, south-facing slopes, neither location is yet threatened by invasive species. The plants examined at both localities demonstrated a reseeding regeneration strategy, which may represent a vulnerability to too-frequent fires. Ericahessequae was found to have an AOO of 8.00 km^2^ and an EOO of 8.00 km^2^. Unfortunately, its habitat is remote and not easily accessible, making estimation of the population size difficult in the absence of more systematic surveys. Until more data can be collected, current conservative estimates put the total population size at fewer than 50 mature individuals. Based on this estimate, an IUCN (2012) category Critically Endangered (CR) under criterion D is recommended.
Pollination syndrome.
The small, open cup-shaped flowers of Ericahessequae have well-developed nectaries and anther appendages, suggesting entomophily. The corolla is remarkably viscid and thus likely to trap crawling and flying insects that would need to land on the surface to effectuate pollination. Therefore, small and medium-sized flying insects that avoid trapping by landing on the recurved adaxial lobes or by entering the corolla through the mouth are the most likely pollinators of this species, although no clear interaction with pollinators has been observed. The lack of a peltate stigma and the fact that pollen is not released in a cloud after manipulation make anemophily highly unlikely.
Etymology.
Ericahessequae is named for the Hessequa clan which occupied most of the modern-day Hessequa municipal area encompassing the Eastern Langeberg Range between the Tradouw Pass and Gourits River.
Subgeneric classification.
Ericahessequae could be placed in section Ceramia (Guthrie & Bolus, 1905) by its glabrous, pitcher-shaped corolla and terminal inflorescence.
Notes on morphology and phylogeny.
Phylogenetic data suggest that E.macrophylla and E.ocellata (sect. Ceramia) are closely related to E.hessequae. While there are morphological similarities among these three Langeberg-endemic taxa, the former two are readily distinguishable by their muticous anthers. In addition, E.macrophylla has a more elongate, urceolate or tubular corolla with a more constricted throat as compared to the stouter bell- to cup-shaped corolla of E.hessequae. Ericaocellata is distinct in having a capitate, 6- to 10-flowered inflorescence (terminal and 3-nate in E.hessequae) with significantly shorter pedicels and sepals as compared to E.hessequae.
Beyond character matching and phylogenetic relatedness, Ericahessequae bears an overall morphological similarity to E.grata and E.cordata, two other species found in the Langeberg. Ericahessequae and E.grata are restricted to the Langeberg Range whilst E.cordata has a wider distribution. Ericahessequae can be distinguished from E.grata by its viscid (as opposed to dry), creamy white (as opposed to light- to red-pink) corolla with a less pronounced constriction at the throat, and by its more densely hairy, lanate (as opposed to hispid) ovary; and from E.cordata by its awned (as opposed to muticous) anthers and subcapitate (as opposed to capitate to peltate) stigma. The leaves in E.grata are usually narrower and more lanceolate with less markedly revolute margins than those of E.hessequae, and it is usually a laxer, upright shrublet preferring wet, south-facing lower to middle slopes. Ericacordata is also mostly upright in habit with softer, shorter side branches. In contrast, Ericahessequae has hard ligneous stems and main branches, a semi-spreading, rounded habit, and preference for high-altitude craggy peaks.
Additional specimens examined.
Western Cape, 3321CD (Sandkraal): Langeberg Mtns, summit of Unnamed Peak (Trig Beacon 45) east of the Langkloof, 1506 m, 33°57.037'S, 21°26.257'E, 22 February 2003, R.C. Turner 701 (NBG).
Erica
inopina
Taxon classificationPlantaeEricalesEricaceae
J.H.J.Vlok sp. nov.
DCE98308-BB4A-53A6-B260-3C8C2234A4D9
urn:lsid:ipni.org:names:77362794-1
Link.
WFO: https://list.worldfloraonline.org/wfo-1000079208.
Type.
South Africa • Western Cape, 3320BB (Laingsburg): Laingsburg district, about 20 km south-east of Touws River, on Lettaskraal Farm, Brandhoek section, 840 m, -33.440°S, 20.373°E, 13 April 2021, J.H.J. Vlok 2988 (NBG, holotype; BG, isotype [2088576]).
Diagnosis.
Ericainopina is closely related and morphologically similar to E.subcapitata. It can be distinguished by its glabrous (as opposed to hairy) sepals, the presence of 3 (as opposed to 0) bracts on the pedicel, and its smooth (as opposed to verrucose) 3-locular (as opposed to 1- or 2- locular) ovary.
Description.
Erect, densely branched, reseeding shrub to 50 cm tall. Branches erect, main branches with many secondary and tertiary flowering branches, pubescent when young with simple spreading hairs. Leaves 3-nate, erect, imbricate, clasping stems, narrowly oblong, blades 2.0–2.5 × 0.6–0.7 mm, apex acute, sulcate, glabrous; petiole 0.3–0.5 mm long, not decurrent. Inflorescence of 1- to 3-nate flowers, mostly axillary in upper leaves, sometimes terminal at the tips of small branches; pedicel ± 0.3 mm long, green, glabrous; bracteoles 2, adnate to calyx, lanceolate, 0.5 × 0.4 mm, with ciliate margins; bract adnate to calyx, lanceolate, 1.5–1.7 × ± 0.4 mm, glabrous with ciliate margins. Calyx equally 4-lobed; sepals adpressed to corolla, lanceolate, 0.7 × 0.4 mm, glabrous, green, margins entire. Corolla equally 4-lobed, urceolate, 1.2–1.7 × ± 1.7 mm, glabrous, yellow green, fading reddish, lobes half to three-quarters the length of corolla, apex obtuse, margins irregular. Stamens 6, connate, partially exserted; anthers golden brown, almost sessile, muticous; pore tear-shaped. Ovary 3-locular, often uniseriate septa, glabrous, smooth; 2 ovules per locule; style 2.0–2.5 mm long, glabrous, stigma exserted, ± 0.3 mm diam., peltate. Fruit and seeds not seen. Flowering time: March to May.
Ericainopina J.H.J.Vlok A flowering branch B single inflorescence. Voucher J.H.J. Vlok 2988 (NBG).
Distribution and habitat.
This species is currently only known from the type locality, where it is locally abundant in deep sandy soil on the ecotone between Arid Fynbos and Renosterveld at the base of a south-facing slope.
Threat status.
Ericainopina is restricted to a single locality in the Laingsburg district, where fewer than 500 plants were observed with no immediate threats and no significant population decline expected. Based on this data, an IUCN (2012) category Vulnerable under criterion D is recommended.
Pollination syndrome.
The exserted style and peltate stigma, small, cup-shaped corolla and the absence of nectaries suggest anemophily.
Etymology.
The specific name refers to the surprise to find an Erica of this group in an arid inland area.
Subgeneric classification.
Ericainopina falls within the Coccospermad group (section Ventiflora) of Erica species that have a fully recaulescent bract and bracteoles (Oliver 2000).
Notes on phylogeny and morphology.
The phylogenetic placement of E.inopina suggests E.subcapitata as its closest relative. These species are also morphologically similar but differ in several features clarified in the diagnosis above. Strict matching with the Erica ID aid yielded no results for E.inopina, irrespective of region and flowering times, and E.subcapitata featured low (32^nd^) using the “probability” algorithm. This highlights the need to integrate multiple lines of evidence for alpha taxonomy in Erica.
Beyond the formal results, E.inopina is also morphologically similar to E.didymocarpa and E.parviporandra, from which it can be distinguished by its glabrous, 3-locular ovary (2- locular in E.didymocarpa; hairy, 1-locular in E.parviporandra) glabrous sepals (hairy in E.didymocarpa), 6 exserted anthers (8, mostly included anthers in E.didymocarpa) and fully recaulescent bract and bracteoles (E.didymocarpa is ebracteolate).
Supplementary Material
XML Treatment for Erica arida
XML Treatment for Erica hessequae
XML Treatment for Erica inopina
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baldwin BG Sanderson MJ Porter JM Wojciechowski MF Campbell S Donoghue MJ Campbell CS Donoghue MJ (1995) The ITS region of nuclear ribosomal DNA: a valuable source of evidence on angiosperm phylogeny.Annals of the Missouri Botanical Garden 82: 247–277. 10.2307/2399880 · doi ↗
- 2Beentje HJ (2006) Ericaceae. In: Beentje HJ Ghazanfar SA (Eds) Flora of Tropical East Africa.Royal Botanic Gardens, Kew, 1–29.
- 3Brown MJM Bachman SP Nic Lughadha E (2023) Three in four undescribed plant species are threatened with extinction.The New Phytologist 240: 1340–1344. 10.1111/nph.1921437583098 · doi ↗ · pubmed ↗
- 4Chernomor O Von Haeseler A Minh BQ (2016) Terrace aware data structure for phylogenomic inference from supermatrices.Systematic Biology 65: 997–1008. 10.1093/sysbio/syw 03727121966 PMC 5066062 · doi ↗ · pubmed ↗
- 5Dulfer H (1964) Revision der südafrikanischen Arten der Gattung Erica L. 1. Teil.Annalen des Naturhistorischen Museums in Wien 67: 79–147.
- 6Dulfer H (1965) Revision der südafrikanischen Arten der Gattung Erica L. 2. Teil (Fortsetzung).Annalen des Naturhistorischen Museums in Wien 68: 25–177.
- 7Elliott AC Bester SP Klopper RR Nelson EC Pirie MD (2024) Curating an online checklist for Erica L. (Ericaceae): Contributing to and supporting global conservation through the World Flora Online.Phyto Keys 243: 121–135. 10.3897/phytokeys.243.12155538947554 PMC 11214009 · doi ↗ · pubmed ↗
- 8Goldblatt P Manning J (2000) Cape plants: a conspectus of the Cape flora of South Africa. Strelitzia 9.National Botanical Institute, Pretoria and Missouri Botanical Garden, St Louis, 743 pp.