Prevalence and characterization of Cryptosporidium in sika deer from Northern China
Yan Tang, Nian-Yu Xue, Yang Gao, Zhen-Qiu Gao, Hong-Di Zhuang, Guang-Rong Bao, Hong-Wei Cao, Jing Liu, Jian-Ming Li, Shuo Liu

TL;DR
This study finds that Cryptosporidium, a zoonotic parasite, infects sika deer in Northern China, with higher rates in young deer and potential for human transmission.
Contribution
The study provides updated epidemiological data and genetic characterization of Cryptosporidium in sika deer in Northern China.
Findings
Cryptosporidium infection rate in sika deer was 14.81% across four Chinese provinces.
Young sika deer had significantly higher infection rates compared to adults.
Cryptosporidium deer genotype and a zoonotic C. ubiquitum subtype were identified.
Abstract
Cryptosporidium spp. are important zoonotic parasites that can cause moderate to severe diarrhea in humans and animals. However, the epidemiological data of Cryptosporidium in sika deer in China need to be updated. In this study, a total of 466 fecal samples were collected from sika deer in Shandong, Jilin, Liaoning, and Heilongjiang provinces. Nested PCR was used to amplify the SSU rRNA gene to detect Cryptosporidium spp. The results showed that the overall infection rate of Cryptosporidium spp. was 14.81%, with no significant differences among regions (p = 0.05). The highest infection rate was found in Heilongjiang Province (23.60%) and the lowest in Jilin Province (10.71%). The infection rate in summer (23.61%) seemed higher than that in autumn (13.20%), but the difference was not statistically significant (p = 0.30). Notably, young sika deer showed a significantly higher infection…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
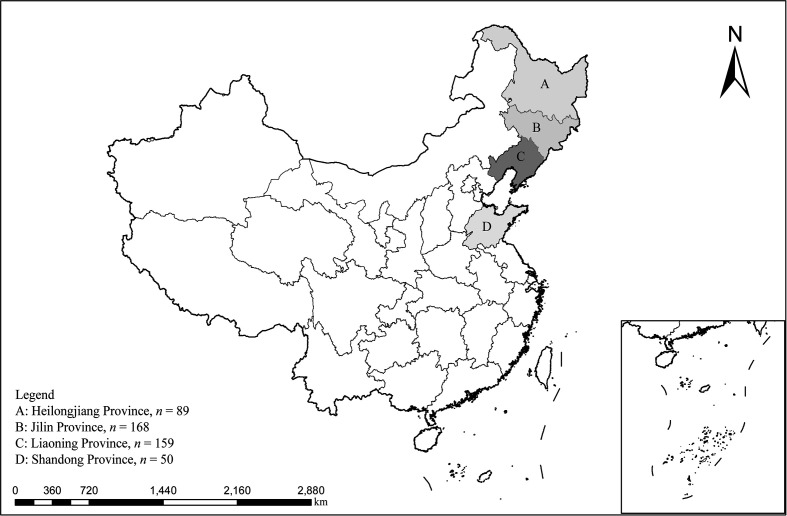 Figure 1
Figure 1 Figure 1
Figure 1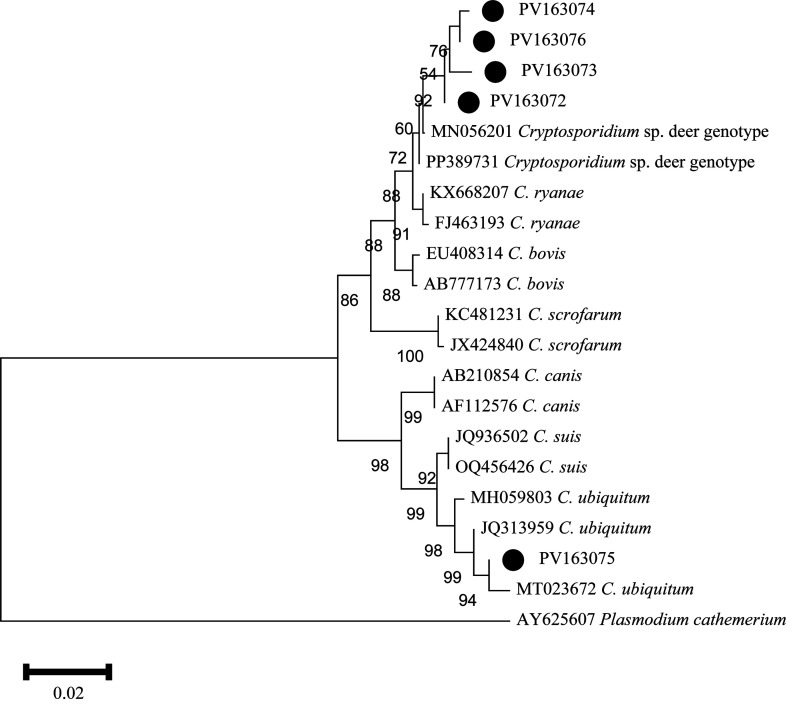 Figure 2
Figure 2 Figure 2
Figure 2- —Jilin Provincial Key Research and Development
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasitic Infections and Diagnostics · Amoebic Infections and Treatments · Dermatological diseases and infestations
Introduction
Cryptosporidium sp. is an important zoonotic parasite that can infect a wide range of vertebrate hosts [37, 43]. It is the second leading cause of diarrhea in children, following rotavirus [7]. Cryptosporidium oocysts excreted by infected hosts can contaminate water and food sources, resulting in infections in both humans and animals [14, 21]. Clinical manifestations of Cryptosporidium infections in humans and animals commonly include diarrhea, growth retardation, and weight loss [35].
To date, more than 47 species and approximately 120 genotypes of Cryptosporidium have been identified using molecular methods and morphological data [13, 16, 41]. Approximately 17 species or genotypes of Cryptosporidium have been identified in deer. These include Cryptosporidium andersoni, C. bovis, C. canis, C. hominis, C. meleagridis, C. muris, C. parvum, C. ryanae, C. scrofarum, C. suis, C. ubiquitum, C. xiaoi, and several genotypes: Cryptosporidium caribou genotype, C. deer genotype, C. parvum genotype II, and C. suis-like genotype [10, 17, 19, 23, 26, 44].
The sika deer (Cervus nippon) is a small species within the Cervinae subfamily, valued for its significant ecological and economic importance. It is classified as a first-grade state-protected animal in China, with its primary habitat in Northeast China [28]. However, research on Cryptosporidium infections in sika deer remains limited. This study examined the prevalence and distribution of Cryptosporidium species and genotypes in sika deer from Northern China.
Materials and methods
Ethics approval
All procedures used in this study were approved by the Research Ethics Committee for the Care and Use of Laboratory Animals in Qingdao Agricultural University, China.
Collection of samples
A total of 466 fecal samples of sika deer were collected using random sampling from May to October 2024. Specifically, Shandong (n = 50), Jilin (n = 168), Liaoning (n = 159), and Heilongjiang (n = 89) (Fig. 1). Specific sampling methods used were as follows: immediately after defecation, fresh fecal samples were collected using sterile polyethylene gloves. To avoid contamination from the environment, only the portions of feces that had not been in contact with the ground were carefully collected. Sample source information was recorded during sampling, sika deer younger than 1.5 years old were classified as young, and those older than 1.5 years old were classified as adult. Samples were stored in frozen sampling boxes and transported to the laboratory, where they were stored at −80 °C until DNA extraction.
Figure 1A map of the People’s Republic of China showing the sampling regions marked with letters.
Extraction of DNA and PCR amplification
Each sample was weighed to 0.5 g, and 0.2 g of glass beads were added for preprocessing. DNA extraction was then performed according to the manufacturer’s instructions. The primers used for nested PCR to amplify SSU rRNA genes for the identification of Cryptosporidium sp. were as previously described (Table 1) [32]. The first round PCR reaction system was 30 μL, containing 15 μL of 2× Mix Master Taq, 1 μL each of forward and reverse primers, 2 μL of DNA template, and 11 μL of deionized water. The reaction program was as follows: after predenaturation for 5 min at 94 °C, 35 cycles were performed, each consisting of 30 s at 94 °C, 30 s at 58 °C, 30 s at 72 °C, and a final extension for 10 min at 72 °C. The second round of PCR was performed using the same reaction system and under the same reaction procedure. The electrophoresis of 6 μL PCR products was performed on 1.0% agarose gel and observed using a UV light.
Table 1. Primer sequences targeting the SSU rRNA gene of Cryptosporidium sp.Primer sequences (5′–3′)Target fragment size (bp)Annealing temperature (°C)F1: GACATATCATTCAAGTTTCTGACC76358R1: CTGAAGGAGTAAGGAACAACCF2: CCTATCAGCTTTAGACGGTAGG58758R2: TCTAAGAATTTCACCTCTGACTGgp60F1: TTTACCCACACCATCTGTAGCGTCG104458gp60R1: ACGGACGGAATGATGTATCTGAgp60F2: ATAGGTGATAATTAGTCAGTCTTTAAT94855gp60R2: TCCAAAAGCGGCTGAGTCAGCATC
Subtype identification
In order to subtype C. ubiquitum, a nested PCR was used to amplify a 948 bp fragment of the 60 kDa glycoprotein (gp60) gene (Table 1) [25]. The reaction system for both rounds of PCR was 50 μL, which include: 25 μL of 2× Mix Master Taq, 1 μL of forward primer, 1 μL of reverse primer, 19 μL of deionized water, and 4 μL DNA template. The PCR amplification procedure was as follow: pre-denaturation for 5 min at 94 °C; then 35 cycles, each cycle consisting of denaturation for 45 s at 94 °C, annealing for 45 s at 58 °C (for primary PCR) or 55 °C (for secondary PCR), and extension for 1 min at 72 °C; finally, extension for 7 min at 72 °C.
Sequencing and phylogenetic analysis
The positive secondary PCR products were delivered to Qingdao Weilai Biotechnology Co., Ltd, China for Sanger sequencing. Sequences obtained were compared with the GenBank sequence to determine Cryptosporidium species. The neighbor-joining (NJ) method was used to build a phylogenetic tree and assess the relationship between obtained sequences and sequences of other Cryptosporidium species/genotypes, with 1,000 bootstrapping replicates to confirm data reliability. The representative gene sequences and the gp60 gene sequence were submitted to the GenBank database under the accession numbers PV163072–PV163076 and PV642472, respectively.
Statistical analysis
A chi-squared test in SPSS software (v.27.0, IBM Corp., Armonk, NY, USA) was used to compare the prevalence of Cryptosporidium in different regions. A value of p < 0.05 was considered statistically significant. Additionally, odds ratios (OR) and 95% confidence intervals (95% CI) were calculated using SPSS software to examine the strength of the association between Cryptosporidium infection and location of sika deer.
Results
Prevalence of Cryptosporidium sp.
In this study, 466 fecal samples of sika deer were tested, and 69 were positive for Cryptosporidium, with an overall infection rate of 14.81%. The infection rates in different provinces ranged from 10.71% to 23.60%, and there was no significant difference (χ^2^ = 7.82, df = 3, p = 0.05). The infection rate was highest in Heilongjiang province (23.6%, 95% CI 15.29–33.03), followed by Shandong province (18.00%, 95% CI 8.41–30.02) and the lowest in Jilin province (10.71%, 95% CI 6.44–15.88). The seasonal analysis showed that there was no significant difference between seasons (χ^2^ = 4.70, df = 1, p = 0.30), and the prevalence of Cryptosporidium in summer (23.61%, 95% CI 14.44–34.18) was higher than that in autumn (13.20%, 95% CI 10.02–16.73). Among different age groups, the prevalence of sika deer was significantly different (χ^2^ = 19.11, df = 1, p < 0.0001), and the infection rate of young sika deer (28.21%, 95% CI 20.38–36.74) was significantly higher than that of adult sika deer (10.32%, 95% CI 7.33–13.74) (Table 2).
Table 2. Prevalence and distribution of Cryptosporidium sp. in sika deer.FactorsCategoryNo. testedNo. positive% (95% CI)HeterogeneityOR(95% CI)χ^2^/df/I^2^(%)/PRegionJilin1681810.71 (6.44–15.88)7.82/3/61.6/0.05–Heilongjiang892123.60 (15.29–33.03)Liaoning1592113.21 (8.35–18.95)Shandong50918.00 (8.41–30.02)SeasonAutumn3945213.20 (10.02–16.73)4.70/1/78.7/0.30–Summer721723.61 (14.44–34.18)AgeAdult3493610.32 (7.33–13.74)19.11/1/94.8/< 0.0001ReferenceYoung1173328.21 (20.38–36.74)3.42 (2.01–5.80)Total–4666914.81 (11.71–18.36)––
Cryptosporidium species/genotypes and subtypes
In this study, based on SSU rRNA analysis, 69 Cryptosporidium sp.-positive samples were identified, including C. deer genotype (n = 68) and C. ubiquitum (n = 1). Additionally, the C. ubiquitum isolate was further subtyped using the gp60 gene and identified as subtype XIIa.
In this study, after 99% clustering of 69 Cryptosporidium sp. sequences, five representative sequences were obtained. These five representative sequences together with 16 Cryptosporidium sp. sequences from GenBank and one exogenous sequence were used to construct a phylogenetic tree. The results showed that PV163072, PV163073, PV163074, and PV163076 clustered in the same branch with the reference sequence MN056201. However, there were subtle differences between these representative sequences and the reference sequence MN056201. Specifically, PV163072 has a single nucleotide polymorphism (SNP), PV163073 has two nucleotide deletions, PV163074 had one SNP, one nucleotide insertion, and three nucleotide deletions, and PV163076 had one nucleotide deletion and one insertion. The representative sequence PV163075 and the reference sequence JQ313959 clustered in the same branch and had 100% similarity (Fig. 2).
Figure 2. Phylogenetic relationships between the sequences in this study and the reference sequences of Cryptosporidium in GenBank and an outgroup, using a neighbor-joining (NJ) method. The genetic distance was calculated based on the Kimura 2-parameter model. Representative nucleotide sequences identified in this study are marked by black dots. The numbers on the branches represent the bootstrap percentage values for 1,000 replicates and are shown in the tree with values greater than 50%.
Discussion
In this study, the overall infection rate of Cryptosporidium sp. in sika deer was 14.81%, which was higher than the global infection rate of deer. For example, the infection rate was 7.5% in sika deer in Japan, 1.5% in red deer in Spain, and 1.42% in Cervidae in Brazil [10, 31, 36]. The infection rate in this study was also higher compared with that in other regions of China, such as the alpine musk deer in Gansu Province (3.9%), Père David’s deer in Jiangsu Province (1.46%), and sika deer in Inner Mongolia, Jilin, Heilongjiang, and Liaoning provinces (13.57%) as a whole [18, 26, 38]. The different infection rates may be due to differences in regional climate, species susceptibility to disease, and sampling time. In addition, in this study, the infection rate of each region ranged from 10.71% to 23.60%, with the highest infection rate in Heilongjiang Province (23.60%), followed by Shandong Province (18.00%), and the lowest infection rate in Jilin Province (10.71%). The sample size may be a factor affecting the difference in infection rates. It is worth noting that the infection rate of young animals (28.21%) in this study was significantly higher than that of adult animals (10.32%), a result consistent with the study of Zhao et al., which may be related to underdeveloped immune function [48].
Phylogenetic analysis revealed close phylogenetic relationships among C. deer genotype, C. ryanae, C. bovis, C. scrofarum, C. canis, C. suis, and C. ubiquitum. Based on epidemiological data, C. deer genotype has only been reported in Cervidae, such as German red deer, Chinese sika deer, and American white-tailed deer, highlighting its host specificity for Cervidae [19, 34, 40]. In contrast, C. ryanae, C. bovis, C. scrofarum, C. canis, and C. suis are found globally in a wide range of mammals, such as Turkish buffalo, Ethiopian calf, Japanese sika deer, Chinese pig, Latvian cattle, Korean dog, etc. [3, 11, 20, 33, 46]. Notably, C. ubiquitum was first identified in white-tailed deer in New York, USA, where it was named C. cervine genotype, and subsequently in goats in Hainan and wild rodents on the Qinghai-Tibet Plateau [30, 39, 47]. Additionally, Ong et al. first detected the presence of C. ubiquitum in humans in 2002. Since then, reports of human infections in this species have gradually increased, with case reports from Sweden, New York, and Spain, etc. [1, 4, 9, 29]. These results indicate that C. ubiquitum is transmissible across hosts and is zoonotic.
In this study, C. deer genotype (98.55%, 68/69) was the dominant species in sika deer, with one case (1.45%) of C. ubiquitum. This finding is consistent with the findings of Tao et al. [38]. Meanwhile, subtype analysis revealed that the *C. ubiquitum-*positive sample in this study belonged to the XIIa subtype, which is consistent with previous findings in sika deer from northeastern China [48]. This subtype has also been detected in Tibetan sheep from Gansu, camels from Xinjiang, chinchillas from Guangdong, and goats from Inner Mongolia [5, 8, 24, 42]. Globally, it has been reported in sheep from Spain and Australia, as well as goats from Algeria [2, 12, 45]. Notably, this subtype has also been identified in humans, including cases from Ethiopia and the United Kingdom [6, 22]. These findings suggest that the XIIa subtype has cross-species transmission potential and zoonotic capability, indicating that sika deer may serve as a potential reservoir for human Cryptosporidium infection.
However, this study still has some limitations, and the sample size in individual areas is small, which may affect an accurate representation of prevalence. Future studies should increase the sample size and use year-round surveillance to fully describe the prevalence of Cryptosporidium in sika deer. Notably, previous studies have shown that C. ubiquitum can be detected in surface water sources, and in particular, there were reports of C. ubiquitum contaminating water sources in the present study area [15, 27]. Therefore, future studies should focus on investigating water sources around sika deer farms to prevent waterborne outbreaks of the disease. In addition, testing of breeding personnel is particularly important to prevent the spread of pathogens to other populations or human communities.
Conclusion
In conclusion, the results of this study indicate that the overall prevalence of Cryptosporidium in sika deer from northern China is 14.81%, suggesting that this parasite is widespread in the local sika deer population. The C. deer genotype was identified as the dominant species in sika deer, and the zoonotic C. ubiquitum subtype (XIIa) was also identified, suggesting that sika deer may play a role in the transmission of Cryptosporidium sp. infection in humans. This study highlights the importance of environmental investigation and personnel screening to provide a basis for risk aversion for public health safety.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alderisio KA, Mergen K, Moessner H, Madison-Antenucci S. 2023. Identification and evaluation of Cryptosporidium species from New York city cases of cryptosporidiosis (2015 to 2018): a watershed perspective. Microbiology Spectrum 11(1), e 0392122.36688660 10.1128/spectrum.03921-22PMC 9927359 · doi ↗ · pubmed ↗
- 2Baroudi D, Hakem A, Adamu H, Amer S, Khelef D, Adjou K, Dahmani H, Chen X, Roellig D, Feng Y, Xiao L. 2018. Zoonotic Cryptosporidium species and subtypes in lambs and goat kids in Algeria. Parasites & Vectors 11(1), 582.30400983 10.1186/s 13071-018-3172-2PMC 6219180 · doi ↗ · pubmed ↗
- 3Bergholm J, Tessema TS, Blomström AL, Berg M. 2024. Detection and molecular characterization of major enteric pathogens in calves in central Ethiopia. BMC Veterinary Research, 20(1), 389.39227796 10.1186/s 12917-024-04258-7PMC 11373192 · doi ↗ · pubmed ↗
- 4Bujila I, Troell K, Ögren J, Hansen A, Killander G, Agudelo L, Lebbad M, Beser J. 2024. Cryptosporidium species and subtypes identified in human domestic cases through the national microbiological surveillance programme in Sweden from 2018 to 2022. BMC Infectious Diseases, 24(1), 146.38291399 10.1186/s 12879-024-09049-x PMC 10826111 · doi ↗ · pubmed ↗
- 5Cao Y, Cui Z, Zhou Q, Jing B, Xu C, Wang T, Qi M, Zhang L. 2020. Genetic diversity of Cryptosporidium in bactrian camels (Camelus bactrianus) in Xinjiang, Northwestern China. Pathogens, 9(11), 946.33202835 10.3390/pathogens 9110946 PMC 7697964 · doi ↗ · pubmed ↗
- 6Chalmers RM, Elwin K, Thomas AL, Guy EC, Mason B. 2009. Long-term Cryptosporidium typing reveals the aetiology and species-specific epidemiology of human cryptosporidiosis in England and Wales, 2000 to 2003. Eurosurveillance, 14(2), 19086.19161717 10.2807/ese.14.02.19086-en · doi ↗ · pubmed ↗
- 7Checkley W, White AC, Jr., Jaganath D, Arrowood MJ, Chalmers RM, Chen XM, Fayer R, Griffiths JK, Guerrant RL, Hedstrom L, Huston CD, Kotloff KL, Kang G, Mead JR, Miller M, Petri WA Jr., Priest JW, Roos DS, Striepen B, Thompson RC, Ward HD, Van Voorhis WA, Xiao L, Zhu G, Houpt ER. 2015. A review of the global burden, novel diagnostics, therapeutics, and vaccine targets for Cryptosporidium. Lancet Infectious Diseases, 15(1), 85–94.25278220 10.1016/S 1473-3099(14)70772-8PMC 4401121 · doi ↗ · pubmed ↗
- 8Chen J, Wang W, Lin Y, Sun L, Li N, Guo Y, Kvac M, Ryan U, Feng Y, Xiao L. Genetic characterizations of Cryptosporidium spp. from pet rodents indicate high zoonotic potential of pathogens from chinchillas. One Health, 13, 100269.10.1016/j.onehlt.2021.100269 PMC 817041834113708 · doi ↗ · pubmed ↗