TLR4 competence and mouse models of sublethal leptospirosis
Olifan Zewdie Abil, Suman Kundu, Leonardo Moura Midon, Maria Gomes-Solecki, Joseph Vinetz, Joseph Vinetz, Joseph Vinetz

TL;DR
This study compares mouse models of leptospirosis and finds that TLR4 function does not determine susceptibility to the disease.
Contribution
The study demonstrates that TLR4 competence is not sufficient to cause susceptibility to sublethal leptospirosis in mice.
Findings
TLR4 competent strains showed similar clinical and molecular signs of leptospirosis as hyporesponsive strains.
Serologic immune factors showed increased IgM and IgG3 with no significant inflammatory markers.
C3H/HeN and C57BL/6 are suitable models for sublethal leptospirosis.
Abstract
Mice are slowly being accepted as alternative models for investigation of leptospiral infection. The strain often used to analyze sublethal disease (C3H/HeJ) expresses a hyporesponsive tlr4 gene in its cells and thus the model is deemed immunocompromised. To help resolve this scientific concern we compared infection of mice expressing competent tlr4 (C3H/HeN, C57BL6) versus tlr4 hyporesponsive mice (C3H/HeJ) with Leptospira interrogans serovar Copenhageni strain Fiocruz L1-130 over a period of two weeks. We found that the two mouse strains with a functional tlr4 gene (C3H/HeN and C57BL/6) developed clinical and molecular signs of leptospirosis less pronounced but not significantly different than tlr4 hyporesponsive C3H/HeJ, as quantified by weight loss, survival curves, presence of Leptospira 16S rRNA in blood and urine and burden of viable spirochetes in kidney as compared to the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
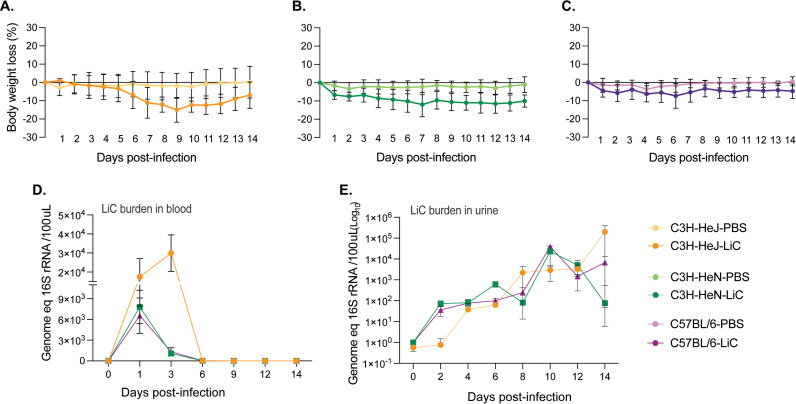 Fig 1
Fig 1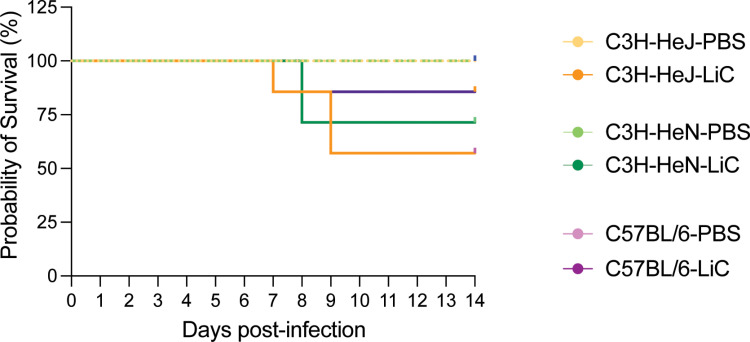 Fig 2
Fig 2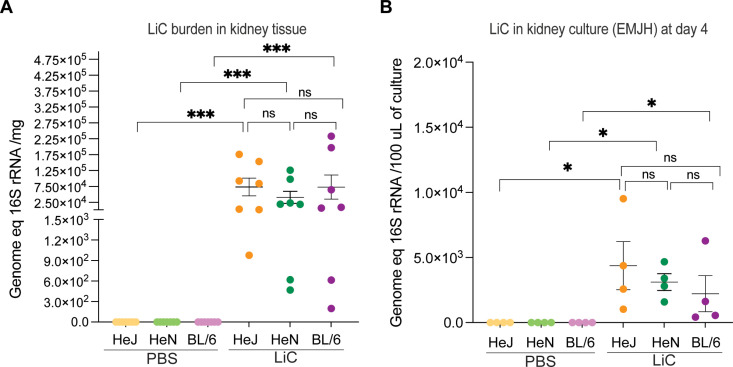 Fig 3
Fig 3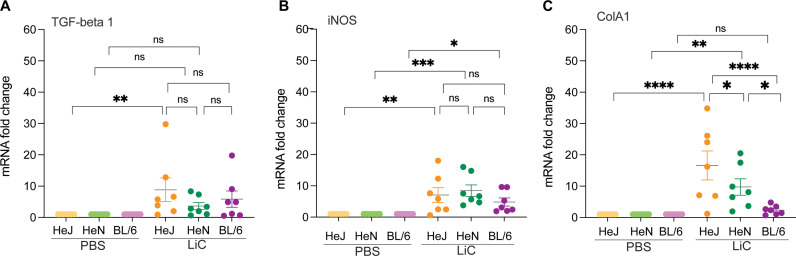 Fig 4
Fig 4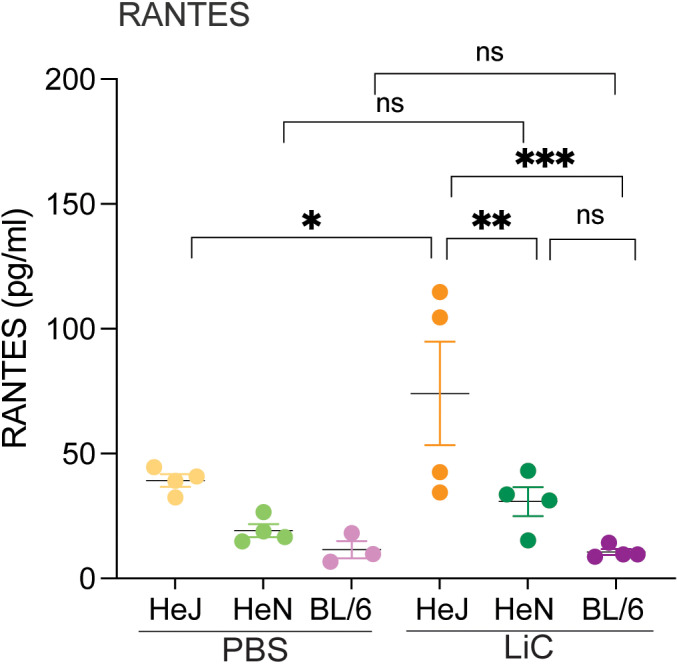 Fig 5
Fig 5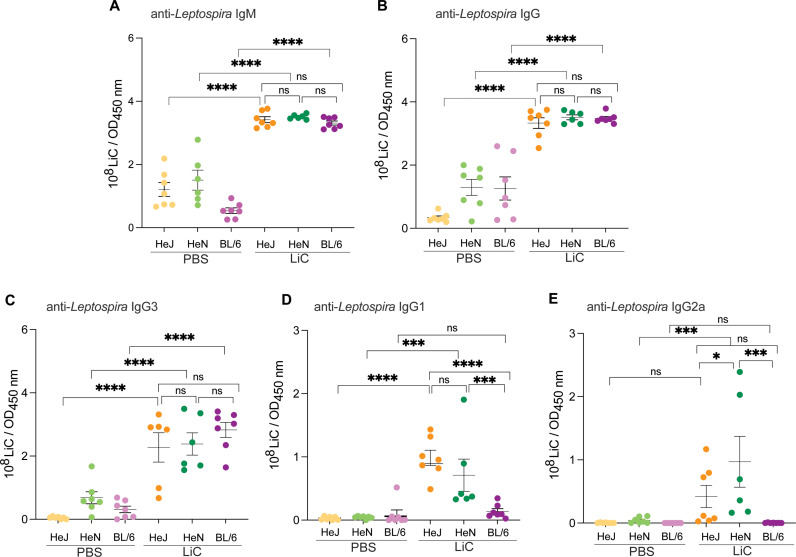 Fig 6
Fig 6- —http://dx.doi.org/10.13039/100000060National Institute of Allergy and Infectious Diseases
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLeptospirosis research and findings · Veterinary medicine and infectious diseases
Introduction
Leptospirosis, an often overlooked but resurging infectious disease caused by a spirochete, is a significant global health concern impacting millions of individuals worldwide. This disease carries a high mortality rate, resulting in approximately 65,000 deaths annually [1]. Furthermore, it poses a serious threat to animals of agricultural importance, leading to substantial economic losses, particularly in tropical and subtropical regions [2]. Although significant efforts have been made to formulate novel vaccination approaches that confer enduring immunity and safeguard against various serovars, our understanding of the specific immune factors contributing to host defense and disease progression remains limited [3].
Leptospirosis research is challenging due to the inconsistent outcomes observed in leptospiral infections involving animals and humans [2], and due to an abundance of high and low virulence Leptospira serovars [4–7]. Hamsters and guinea pigs have been widely used as small-animal models of acute disease, as they recapitulate the manifestations of severe disease in humans [8,9]. Rats have been used as a model for studying the severity of human leptospirosis as they become chronically infected and shed Leptospira in urine many months after infection [10]. However, the usage of these animals in research is limited in some parts of the world where the disease is endemic due to stringent animal regulations [11] and the scarcity of accessible reagents for routine experiments.
The outcomes of experimental leptospiral infection have been analyzed using various mouse models, including studies on lethal, sublethal, and chronic leptospirosis [12–14]. For instance, C57BL/6 and BALB/c are more resilient to acute disease and have the potential to serve as models for persistent infection caused by Leptospira interrogans [13,14]. C3H/HeJ mice infected with L. interrogans develop disease that can be easily monitored through measurement of clinical scores. This strain produces valuable models of lethal [12,15,16] and sublethal disease [17,18], and has been used to study inflammatory signatures of infection [19], necroptosis [20], and immunity protection [21].Toll-like receptor (TLR) 4, which recognizes LPS, plays a central role in the control of leptospirosis [22,23]. One study described how leptospiral-LPS activates murine, but not human, TLR4 in cultured macrophages [24], and it is associated with resistance to infection [25]. C3H/HeJ mice have a single amino acid substitution (aa712, P to H) within the coding region of the tlr4 gene that makes this molecule hyporesponsive to the atypical Leptospira LPS [23,26]. Of note, humans, also believed to be susceptible to leptospirosis express a TLR4 molecule that does not sense the atypical Leptospira LPS [24]. Nevertheless, one consistent criticism regarding the use C3H/HeJ mice is that its hyporesponsive Toll-Like Receptor 4 (TLR4) [26] qualifies these mice as immunocompromised. To help resolve this valid scientific concern we did a study in which we compared infection with L. interrogans in mice expressing competent tlr4 (C57BL6, C3H/HeN) versus mice that are tlr4 hyporesponsive (C3H/HeJ). The goal of this study was to determine if the TLR4 competent strains can also be used as mouse models of sublethal leptospirosis.
Results
Weight loss, burden in blood, shedding in urine and survival after infection with L. interrogans serovar Copenhageni FioCruz.
Following experimental leptospiral infection (10^8^ LiC), mice were monitored over a 15-day period. In C3H/HeJ, steady weight loss was observed beginning on day 6, reaching the lowest mean on day 9 (-14.9%), at which point mice recovered gradually (Fig 1A). C3H/HeN mice exhibited weight loss from day 1 reaching the lowest mean on day 7 (-11.9%) which was maintained until day 14 (Fig 1B). C57BL/6 mice experienced lower loss of weight throughout the 14 days, reaching the lowest mean (-7.4%) on day 6 (Fig 1C). The weight loss curves of the three infected mouse strains were statistically significant compared to their respective controls (p < 0.0001).
Weight loss and bacterial burden in blood and urine after infection.Male C3H/HeJ, C3H/HeN and C57BL/6 mice (n = 7/group) were inoculated IP with 108 L. interrogans serovar Copenhageni strain FioCruz L1-130 (LiC) and with PBS as control. A-C) Body weight (% change) was recorded daily for 15 days post infection; D-E) Quantification of bacterial load in blood and urine by qPCR of the 16S rRNA gene. Data from two independent experiments is shown. For weight loss, statistics by Ordinary 2-Way ANOVA, p < 0.0001 for infected versus control C3H/HeJ, C3H/HeN and C57BL/6.
Quantitative PCR (qPCR) targeting the Leptospira 16S rRNA gene was performed to analyse the dissemination of Leptospira in blood and urine. Blood samples were collected on days 0, 1, 3, 6, 9, 12, and 14 post-inoculation to quantify the Leptospira burden. Peak Leptospira burdens in blood were observed on day 3 in C3H/HeJ mice (~3x10^4^), whereas the peak burden occurred on day 1 in both C3H/HeN and C57BL/6 mice, ~ 6-9x10^3^ (Fig 1D). Leptospira shedding in urine was also assessed using qPCR on alternate days. Shedding (Fig 1E) was detected at low levels (~10^2^) during the first week post-infection and gradually increased in the second week with peaks on day 10 for C3H/HeN and C57BL/6 (~10^4^), and on day 14 for C3H/HeJ (~10^6^).
Regarding survival, none of the 9–10 week old mouse strains injected with 10^8^LiC reached the LD50: 57.1% of C3H/HeJ, 71.4% of C3H/HeN and 85.7% C57BL/6 survived at 14 days post-infection in comparison with 100% survival in each of the control groups (Fig 2). Differences are not statistically significant.
Probability of survival.Male C3H/HeJ, C3H/HeN and C57BL/6 mice (n = 7/group) were inoculated IP with 108 L. interrogans serovar Copenhageni strain FioCruz L1-130 (LiC) and with PBS as control. The percentage of mice that met endpoint criteria for euthanasia before and at the term of two independent experiments is shown. Statistics by Log-rank (Mantel-Cox) test, p = 0.1088.
Burden and viability of L. interrogans serovar Copenhageni FioCruz in kidney tissue
To assess renal colonization by L. interrogans, kidney tissues from infected mice were collected on day 15 post-infection and analyzed for the presence of live Leptospira under a dark field microscope and for DNA quantification using qPCR. The bacterial burden was ~ 7.3 × 10⁴ Leptospira per mg of kidney tissue in C3H/HeJ, ~ 4.1 × 10⁴ in C3H/HeN, and ~5.4 × 10⁴ in C57BL/6 infected mice (Fig 3A). Differences between infected and control groups are significant, p < 0.001. Additionally, the viability of Leptospira was assessed by culturing kidney tissues in EMJH medium at 30°C for 4 days, visualizing live motile Leptospira under a dark field microscope and quantifying spirochetes using qPCR. On day 4 of culture, the average number of spirochetes per 100 μL of EMJH culture was ~ 4.3x10^3^ for C3H/HeJ, ~ 3.1 × 10³ for C3H/HeN, and ~2.2 × 10³ for C57BL/6 mice. Differences between infected and control groups are significant, p < 0.05 (Fig 3B).
Burden and viability of Leptospira in kidney.A) Kidney tissues were collected 2 weeks post-infection for qPCR analysis of the 16S rRNA gene; data from two independent experiments is shown; B) kidney was placed in culture to evaluate motility of L. interrogans under a dark field microscope on day 4, and qPCR was used to quantify Leptospira; data from one experiment is shown. Statistics by Mann-Whitney U test, *** p < 0.001 and * p < 0.05.
Inflammatory cytokines and signatures of fibrosis in kidney
To assess the upregulation of inflammatory cytokines critical for immune responses and fibrosis markers associated with kidney damage, kidney was collected from experimental mice on day 15 post-infection and processed for mRNA expression analysis of TNF-α, TGF- β1, IL-1β, IFN-γ, IL-6, IL-23, IL-17a, iNOS and ColA1 by RT-PCR using glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as the calibrator to normalize gene expression. Expression of TNF-α, IL-1β, IFN-γ, IL-6, IL-23, IL-17a were not different between infected and uninfected groups. The expression TGF- β1 mRNA, was upregulated in C3H/HeJ mice in comparison to uninfected control; however differences between the mouse strains were not significant (Fig 4A). In addition, mRNA of the fibrosis marker inducible Nitric Oxide Synthase (iNOS), was increased in C3H/HeJ, C3H/HeN and C57BL/6 in comparison to the respective controls; however differences between the mouse strains were not significant (Fig 4B). The mRNA expression of collagen A1 (ColA1) was elevated in both C3H/HeJ and C3H/HeN mice compared to the respective controls in contrast to C57BL/6, and differences between the mouse strains were significant (Fig 4C).
Inflammatory and fibrosis markers in kidney.Kidney tissues collected 2 weeks post-infection were evaluated for mRNA expression analysis of 9 inflammatory markers by RT-PCR normalized against GAPDH. We show TGF-beta 1 (A), iNOS (B) and ColA1 (C) as these markers produced differences between the three mouse strains in two independent experiments. Statistics by Two-way ANOVA followed by Tukey’s multiple comparisons test, * p < 0.05, ** p < 0.005, *** p < 0.0005, **** p < 0.0001.
Inflammatory markers circulating in serum
The concentration of chemokines CxCL1/KC/GRO-α, CxCL2/MIP-2, CCL5/RANTES and cytokines TNF-α, IL-1β, IFN-γ, IL-5, IL-6, IL-9, IL-10, IL-13, IL-17, IL-23 circulating in blood were measured in experimental animals on day 15 post-infection. Only one chemokine, CCL5/RANTES, was increased in infected C3H/HeJ in comparison to the respective uninfected control. None of the inflammatory markers tested were increased in serum from C3H/HeN or C57BL/6 two weeks post infection. Differences in CCL5/RANTES between C3H/HeJ and C3H/HeN, as well as C3H/HeJ and C57BL/6 were significant (Fig 5).
*Inflammatory markers circulating in serum.The concentration of 13 inflammatory markers was measured in serum from experimental mice collected 2 weeks post-infection. Shown is RANTES, the only chemokine detected. Data from one experiment is shown. Statistics by Two-way ANOVA followed by Tukey’s multiple comparisons test, *p < 0.05, ** p < 0.005, **p < 0.0005.
Serological antibody responses
Terminal blood samples were collected 15 days post-infection to evaluate the levels of antibody production to L. interrogans (Fig 6). As expected, the three strains of infected mice produced increased levels of anti-Leptospira-specific IgM (Fig 6A) and total IgG (Fig 6B) compared to the respective uninfected controls; no differences were observed between the mouse strains. IgG isotyping produced the following results on d15 post-infection: IgG3 mirrored the total IgG profile for the three mouse strains (Fig 6C) and IgG1/IgG2a were not detected in infected C57BL/6 compared to the uninfected control (Fig 6D and 6E). IgG1 was increased in infected C3H/HeJ and C3H/HeN compared to the respective controls but differences between the C3H mouse strains were not significant (Fig 6D). IgG2a was only increased in infected C3H/HeN compared to the uninfected control, and differences between this strain and C3H/HeJ as well as C57BL/6 were significant (Fig 6E).
Antibody responses to Leptospira. L. interrogans serovar Copenhageni FioCruz specific IgM, IgG, IgG3, IgG1 and IgG2a were measured in serum from experimental mice by ELISA on day 15 post-infection.Data from two independent experiments is shown. Statistics by Two-way ANOVA with uncorrected Fisher’s LSD test, * p < 0.05, *** p < 0.0005, **** p < 0.0001.
Discussion
We previously used the C3H/HeJ strain to develop a mouse model of experimental sublethal leptospirosis [17,27]. Since resistance to acute lethal infection with L. interrogans was associated with a functional tlr4 [25] and C3H/HeJ is hyporesponsive [26] to L. interrogans atypical LPS, there are valid concerns that this mouse model does not recapitulate a competent immune response to this spirochete. In this study, we did a side-by-side comparative analysis using three mouse strains (C3H/HeJ, C3H/HeN and C57BL/6). The two mouse strains with a functional tlr4 gene (C3H/HeN and C57BL/6) developed clinical and molecular signs of sublethal leptospirosis that were less pronounced but not significantly different than C3H/HeJ. Our data indicates that TLR4-competent C3H/HeN and C57BL/6 are appropriate mouse models of sublethal leptospirosis.
We found that weight loss differences between strains are more evident in C3H mice throughout the 15-day infection period (Fig 1). For a choice of animal models of disease, differences in weight loss in the C3H background produced clinical scores that may be easier to reproduce consistently. qPCR analysis of Leptospira 16S rRNA in blood, urine and kidney tissue showed no differences in the dynamics of L. interrogans dissemination between the three mouse strains, except for the load of Leptospira in blood which was higher in 9–10 week old C3H/HeJ (Figs 1 and 3). These data reproduce previous studies done using age/IP-dose matched C3H/HeJ [18] and C57BL/6 [28] mice.
There are numerous links between competent recognition of Leptospiral LPS by TLR4 and resistance to infection in mice [13,22–25]. Our data shows that susceptibility to sublethal infection can be observed in 9–10 week old C3H/HeN and C57BL/6 mice, which express competent TLR4 (Figs 1 and 2). Nevertheless, our data also shows that C3H-HeJ, expressing a hyporesponsive tlr4, consistently produced larger metrics of disease progression. Of note, humans express a tlr4 molecule that does not sense the atypical Leptospira LPS [24]. One could therefore speculate that the C3H/HeJ mouse may recapitulate the human immune response to pathogenic Leptospira more closely than tlr4 competent mice. In another study we found that inoculation of potentially susceptible TLR4/MD-2 humanized transgenic mice with L. interrogans did not produce different measurements of disease progression or Leptospira dissemination than wildtype mice [28].This may have been related to the humanized transgenic mouse background being C57BL/6. Overall, the data suggests that although TLR4 plays a role in susceptibility to Leptospira infection, it is not the main determinant as other receptors in mammalian cells (ex. TLR2) likely compensate for deficient receptor binding.
We analysed mRNA expression of 9 inflammatory markers (TGF-β1, TNF-α, IL-1β, IFN-γ, IL-6, IL-23, IL-17a, iNOS and ColA1) in kidney of experimental mice at 15 days post infection (Fig 4). We found that C3H/HeJ had increased TGF-beta 1, iNOS and ColA1, C3H/HeN had increased iNOS and ColA1, and C57BL/6 has iNOS increased but not ColA1. When we compare the C3H/HeJ data with a previous study using age/IP-dose matched mice studies [18], normalized for markers tested in both studies, we find the iNOS and ColA1 data in agreement, but not TNF-α and IFN-γ. Similarly, when we compare the C57BL/6 data with a previous study using age/IP-dose matched mice [28] we find discrepancies in iNOS, ColA1 and TNF-α. These differences suggest that although expression markers of inflammation in kidney may be statistically different in the reported studies, it still raises a question of biological relevance and should be interpreted with caution.
Regarding inflammatory markers circulating in serum we tested for markers engaged in innate (CxCL1/KC/GRO-α, CxCL2/MIP-2, CCL5/RANTES, TNF-α, IL-1β) and adaptive Th1, Th2 and Th17 responses (IFN-γ, IL-5, IL-6, IL-9, IL-10, IL-13, IL-17, IL-23) and found that only CCL5/RANTES was increased in C3H/HeJ at day 15 post infection (Fig 5). CCL5/RANTES is a chemokine important for the recruitment of T cells, neutrophils and NK cells. In our previous studies using age/IP-dose matched C3H/HeJ, we found that CCL5/RANTES was increased in serum at 24h and 72h post sublethal infection [19]. Furthermore, CCL5/RANTES was found increased in kidney, spleen and blood of C57BL/6J mice at 72h post-infection with 108 L. interrogans serovar Manilae in a model of lethal leptospirosis [29]. Now looking at the human leptospirosis landscape on CCL5/RANTES, low levels of serologic CCL5/RANTES were found in patients with fatal leptospirosis [30] and CCL5/RANTES was the most increased chemokine found in serum of patients diagnosed with non-lethal leptospirosis associated with L. interrogans serovars Copenhageni and Icteroheamorrhagiae [31]. Overall, the data from Mus and Homo strongly suggests that CCL5/RANTES plays an important role in the immune response to L. interrogans infection. We will further characterize this chemokine in future analysis of inflammatory markers of lethal and sublethal leptospirosis.
While innate immune responses are the main immunological pathway for eliminating Leptospira, the humoral immune response plays a vital role in effectively eradicating the bacteria and expelling it from the host [32]. Our present observations (Fig 6) revealed that LiC infection led to an increase of Leptospira-specific IgM, IgG antibodies in the blood of the three mouse strains, which was expected [17,25,33,34]. IgG isotyping produced some interesting results for IgG3 and IgG1. IgG3, which is T-cell independent, was significantly increased in the three mouse strains and explains high total IgG for C57BL/6 in our study. Two weeks after infection, IgG1 was extremely low in C57BL/6, as observed by others [34], and significantly higher in C3H/HeJ and C3H/HeN. The production of IgG1 antibody in C3H mice suggests a Th2-biased engagement of the immune response at two weeks post infection, as shown in our previous studies in C3H/HeJ [17].
In conclusion, our data shows that C3H/HeN and C57BL/6 mice, both TLR4 competent strains, can be used to recapitulate sublethal leptospirosis as they produce unambiguous differences in clinical and molecular measurements of disease progression, Leptospira dissemination to tissues, colonization of kidney by live spirochetes and shedding in urine. Thus, competent recognition of L. interrogans serovar Copenhageni FioCruz factors by murine TLR4 does not determine susceptibility to disease. Each mouse strain has characteristics that can be leveraged in pursuit of knowledge on immunity and host response to pathogenic Leptospira species.
Materials and methods
Ethics Statement
All experiments with animals were performed in compliance with the University of Tennessee Health Science Center (UTHSC) Institutional Animal Care and Use Committee (IACUC), Protocol no. 22–0362.
Animals
Age matched 9–10 weeks, male C3H/HeJ, C3H/HeN, C57BL/6 mice were used. C3H/HeJ animals were purchased from The Jackson Laboratory (Bar Harbor); C3H/HeN and C57BL/6 were purchased from Charles River (Wilmington, MA) and maintained in specific pathogen-free at the Laboratory Animal Care Unit of the University of Tennessee Health Science Centre, with unrestricted access to food and water. Male mice were used because they are more susceptible to leptospirosis. Two independent experiments were done: the first was comprised of 6 groups of mice with 3 animals per group; the second, was comprised of 6 groups of mice with 4 animals per group; data from both experiments were combined for analysis.
Bacterial strains and culture
L. interrogans serovar Copenhageni strain Fiocruz L1-130 (henceforth LiC), frozen in -80°C, was passaged in hamster. Their kidneys were harvested and cultivated in 4mL of Hornsby-Alt-Nally (HAN) media [35] with 100 μg/mL 5-fluorouracil (MP Chemicals, CA) at 29°C for better growth. Passage 2 was done in Ellinghausen-McCullough-Johnson-Harris (EMJH) medium supplemented with Difco Leptospira enrichment EMJH (Becton, MD) at 28–30°C. EMJH culture passage 2 was allowed to reach the log phase of growth, pelleted by centrifugation at 3,000 × g for 5 min, and washed and resuspended in sterile 1 × phosphate-buffered saline (PBS) (Thermo Fisher Scientific). Next, the cells were used to infect the animals after counted under a dark-field microscope (Zeiss USA, Hawthorne, NY) using a Petroff- Hausser chamber as previously described [27]. We found that recovering -80C frozen Leptospira in HAN media before passage in EMJH allowed for faster recovery of the culture.
Animal infection and collection of specimens
Mice were inoculated with 1 × 10^8^ LiC in 200 μL of endotoxin-free 1 × PBS intraperitoneally (i.p.). Control mice were inoculated with the same volume of endotoxin-free 1 × PBS. Survival and body weight loss were monitored for 15 consecutive days post-infection. Mice were euthanized 15 days post-infection or when they reached the humane endpoint criteria (20% body weight loss). Blood, urine, and kidney were collected for further analysis. Immediately after the mice were restrained, the bladder was massaged, and Eppendorf tubes were used to collect the excreted urine.
Bacterial quantification
The NucleoSpin tissue kit (Clontech, Mountain View, CA) was used to purify genomic DNA from kidney, blood and urine following the manufacturer’s protocol, and the purified DNA was then stored at −20°C for further analysis. To quantify Leptospira, quantitative polymerase chain reaction (qPCR) was performed using Leptospira 16S rRNA primers (Forward: CCCGCGTCCGATTAG and Reverse: TCCATTGTGGCCGAACAC) and a TAMRA probe (CTCACCAAGGCGACGATCGGTAGC) obtained from Eurofins (Huntsville, AL). The results were reported as the number of Leptospira genome equivalents. The qPCR mixture consisted of 25 μM of each primer, 250 nM of the specific probe, and 2 μL of DNA sample, with a total volume of 20 μL. Duplicate reactions were performed. The amplification protocol included an initial step of 10 min at 95°C, followed by 40 cycles of amplification (15 s at 95°C and 1 min at 60°C). The analysis was conducted using the comparative threshold cycle (CT) method. A negative result was determined if no amplification occurred.
Measurement of Leptospira-specific antibody
Leptospira-specific antibody levels were measured using the enzyme-linked immunosorbent assay (ELISA). The process of preparing a leptospiral extract for LiC was followed as described [36]. Briefly, LiC was cultured in EMJH media until it reached optimal cell density. The bacterial cells were then separated by centrifugation to form a pellet. This pellet was subjected to incubation with BugBuster solution (1mL) at room temperature in a shaker (100 rpm) for 20 min and mixed thoroughly by vortexing. The resulting mixture was stored at -20°C. The whole cell extract of Leptospira was appropriately diluted in a sodium carbonate coating buffer with a concentration of 1X. A 96-well flat-bottom ELISA microtiter plate (Nunc-eBioscience) was coated overnight at 4^o^C with 100 µL 1X sodium carbonate coating buffer whole-cell extract of Leptospira (10^7^–10^8^ bacteria per well). After overnight incubation, the ELISA plate was washed with 1X PBST. The plate was blocked by a blocking buffer (100 µL/well) containing 1% BSA, followed by incubation for 1 h at 37^o^C. After washing, serum samples (1:100) was added, and the plate was incubated for 1 h at 37^o^C. The unbound primary antibody was removed by vigorous washing. Next, anti-mouse secondary antibodies for IgM IgG, IgG3, IgG1 or IgG2a (all from Cell signaling technology, CST) conjugated with horseradish peroxidase was added, which was incubated for 30 min, followed by standard color development using TMB Sureblue. Absorbance measurement was carried out at OD 450 nm using Molecular Devices Spetramax.
Expression of inflammatory and fibrosis markers
Total RNA was extracted using the RNeasy Mini Kit (QIAGEN) from kidney according to the manufacturer’s protocol. Complementary DNA (cDNA) was synthesized from the extracted RNA using the RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific). The resulting first-strand cDNA served as the template for reverse transcription PCR (RT-qPCR), which was performed on a QuantStudio 3 Real-Time PCR (Applied Biosystems) using the PowerTrack SYBR Green Master Mix (Applied Biosystems). Each RT-qPCR reaction (10 μL total volume) included the cDNA template and specific primers. The cycling conditions were as follows: an initial step at 50°C for 2 minutes, followed by denaturation at 95°C for 2 minutes, followed by 40 cycles of 95°C for 15 sec for denaturation, and 60°C for 1 min for annealing/extension. A melt curve analysis was conducted at the end of the amplification to confirm the specificity of the PCR products. Relative gene expression levels across samples were quantified by the double delta Ct (2^-∆∆Ct^) method, with glyceraldehyde 3-phosphate dehydrogenase (GAPDH) serving as the endogenous reference control. The primer sequences used are listed in S1 Table in the Source Data file.
Measurement of chemokines and cytokines in blood
Serum from experimental mice was derived from blood collected after euthanasia on day 15 post-infection and frozen at -80C. Analysis of circulating chemokines and cytokines was done using ProcartaPlex Multiplex Immunoassay (eBioscience) according to the manufacturer’s instructions. The data were acquired using a Luminex 200 reader. The concentrations of each inflammatory marker were determined based on a standard curve.
Statistical analysis
For the exploratory animal research described in this study, we used the “resource equation” approach to calculate sample size. Based on this approach, the acceptable range of degrees of freedom (DF) for the error term in an analysis of variance (ANOVA) is between 10–20 animals [37]. This method allows for determination of the minimum number of animals to produce significant results. We used GraphPad Prism 10 software. For analysis of weight loss (Fig 1) we used Ordinary 2-Way ANOVA; for survival (Fig 2) we used Mantel-Cox Log rank test; for burden and viability of LiC (Fig 3) we used Mann-Whitney U test; for analysis of inflammatory markers (Figs 4, and 5) we used Two-way ANOVA with Tukey’s multiple comparison test; for analysis of antibody class and IgG isotypes (Fig 6) we used Two-Way ANOVA with uncorrected Fisher’s LSD.
Supporting information
S1 TableExcel file containing the source data used in all figures, and S1 Table containing a list of primers for inflammatory and fibrosis markers.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Costa F, Hagan JE, Calcagno J, Kane M, Torgerson P, Martinez-Silveira MS, et al. Global Morbidity and Mortality of Leptospirosis: A Systematic Review. P Lo S Negl Trop Dis. 2015;9(9):e 0003898. doi: 10.1371/journal.pntd.0003898 26379143 PMC 4574773 · doi ↗ · pubmed ↗
- 2Haake DA, Levett PN. Leptospirosis in humans. Curr Top Microbiol Immunol. 2015;387:65–97. doi: 10.1007/978-3-662-45059-8_5 25388133 PMC 4442676 · doi ↗ · pubmed ↗
- 3Mc Bride AJ, Athanazio DA, Reis MG, Ko AI. Leptospirosis. Current Opinion in Infectious Diseases. 2005;18(5):376–86. doi: 10.1097/01.qco.0000178824.05715.2c 16148523 · doi ↗ · pubmed ↗
- 4Silva EF, Santos CS, Athanazio DA, Seyffert N, Seixas FK, Cerqueira GM, et al. Characterization of virulence of Leptospira isolates in a hamster model. Vaccine. 2008;26(31):3892–6. doi: 10.1016/j.vaccine.2008.04.085 18547690 PMC 2519131 · doi ↗ · pubmed ↗
- 5Lehmann JS, Matthias MA, Vinetz JM, Fouts DE. Leptospiral pathogenomics. Pathogens. 2014;3(2):280–308. doi: 10.3390/pathogens 3020280 25437801 PMC 4243447 · doi ↗ · pubmed ↗
- 6Philip N, Jani J, Azhari NN, Sekawi Z, Neela VK. In vivo and in silico Virulence Analysis of Leptospira Species Isolated From Environments and Rodents in Leptospirosis Outbreak Areas in Malaysia. Front Microbiol. 2021;12:753328. doi: 10.3389/fmicb.2021.753328 34803975 PMC 8602918 · doi ↗ · pubmed ↗
- 7Giraud-Gatineau A, Nieves C, Harrison LB, Benaroudj N, Veyrier FJ, Picardeau M. Evolutionary insights into the emergence of virulent Leptospira spirochetes. P Lo S Pathog. 2024;20(7):e 1012161. doi: 10.1371/journal.ppat.1012161 39018329 PMC 11285912 · doi ↗ · pubmed ↗
- 8van den Ingh TS, Hartman EG. Pathology of acute Leptospira interrogans serotype icterohaemorrhagiae infection in the Syrian hamster. Vet Microbiol. 1986;12(4):367–76. doi: 10.1016/0378-1135(86)90086-6 3538643 · doi ↗ · pubmed ↗