Xenopus laevis as an infection model for human pathogenic bacteria
Ayano Kuriu, Kazuya Ishikawa, Kohsuke Tsuchiya, Kazuyuki Furuta, Chikara Kaito

TL;DR
Juvenile Xenopus frogs can be used as a model to study human pathogenic bacteria and test antimicrobial treatments.
Contribution
This study demonstrates that Xenopus laevis is a viable model for evaluating virulence and antimicrobial efficacy of human pathogenic bacteria.
Findings
Xenopus frogs die when injected with pathogenic bacteria like Staphylococcus aureus and Pseudomonas aeruginosa.
Antibiotics can reduce mortality caused by these bacteria in Xenopus frogs.
Virulence gene-deleted bacterial strains show reduced pathogenicity in Xenopus frogs.
Abstract
Animal infection models are essential for understanding bacterial pathogenicity and corresponding host immune responses. In this study, we investigated whether juvenile Xenopus laevis could be used as an infection model for human pathogenic bacteria. Xenopus frogs succumbed to intraperitoneal injection containing the human pathogenic bacteria Staphylococcus aureus, Pseudomonas aeruginosa, and Listeria monocytogenes. In contrast, non-pathogenic bacteria Bacillus subtilis and Escherichia coli did not induce mortality in Xenopus frogs. The administration of appropriate antibiotics suppressed mortality caused by S. aureus and P. aeruginosa. Strains lacking the agr locus, cvfA (rny) gene, or hemolysin genes in S. aureus, LIPI-1-deleted mutant of L. monocytogenes, which attenuate virulence within mammals, exhibited reduced virulence in Xenopus frogs compared with their respective wild-type…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
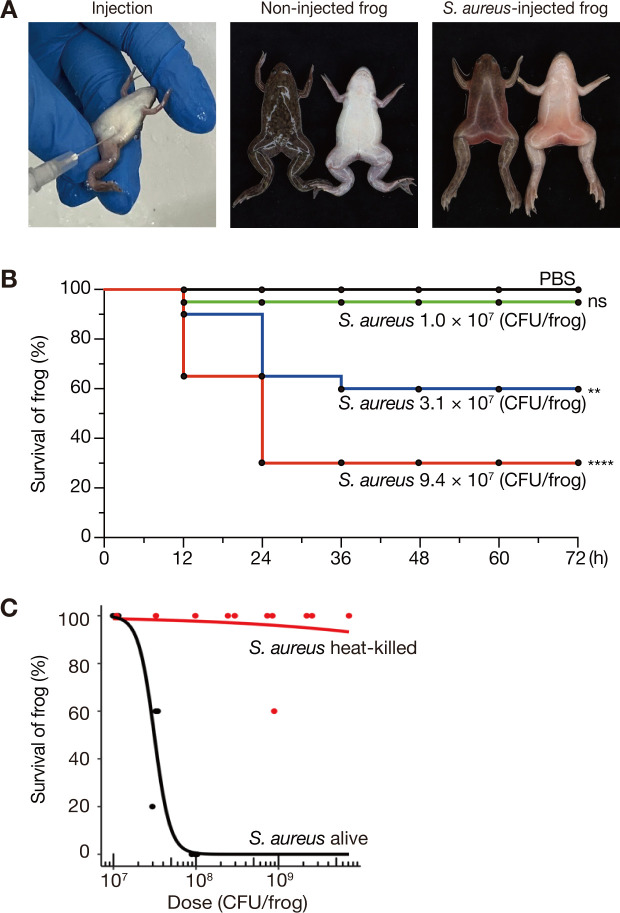 Fig 1
Fig 1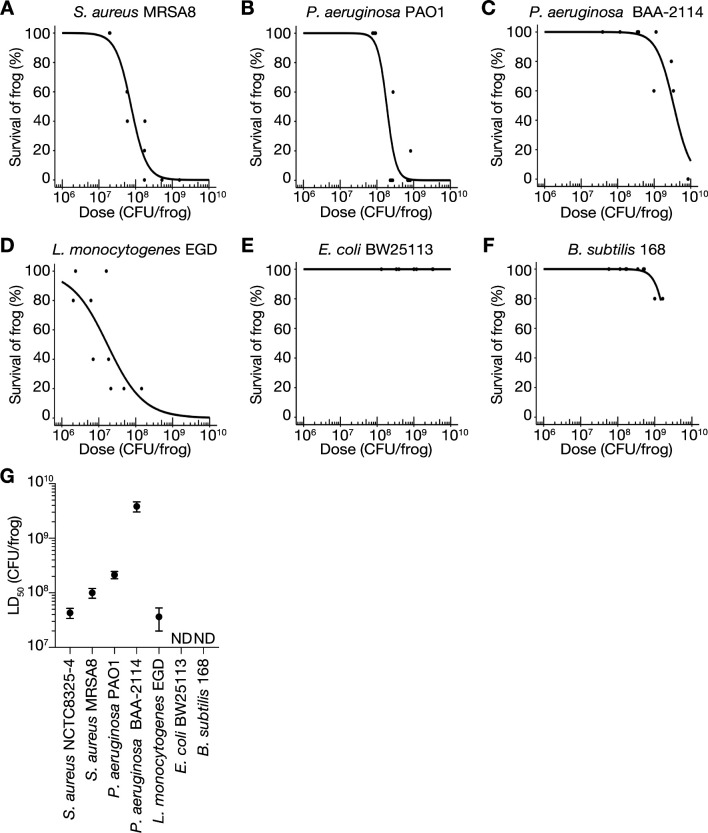 Fig 2
Fig 2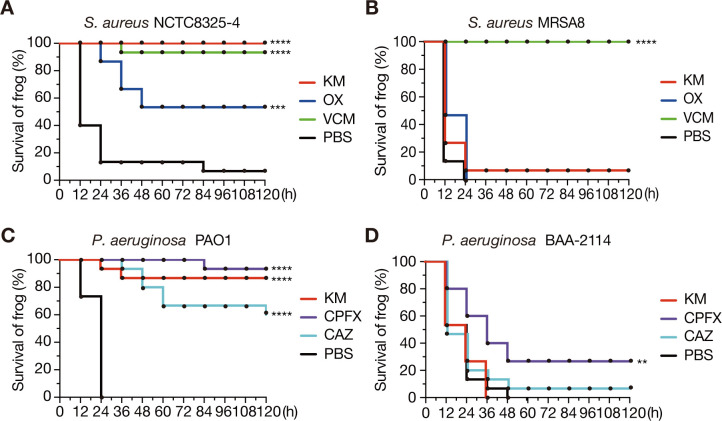 Fig 3
Fig 3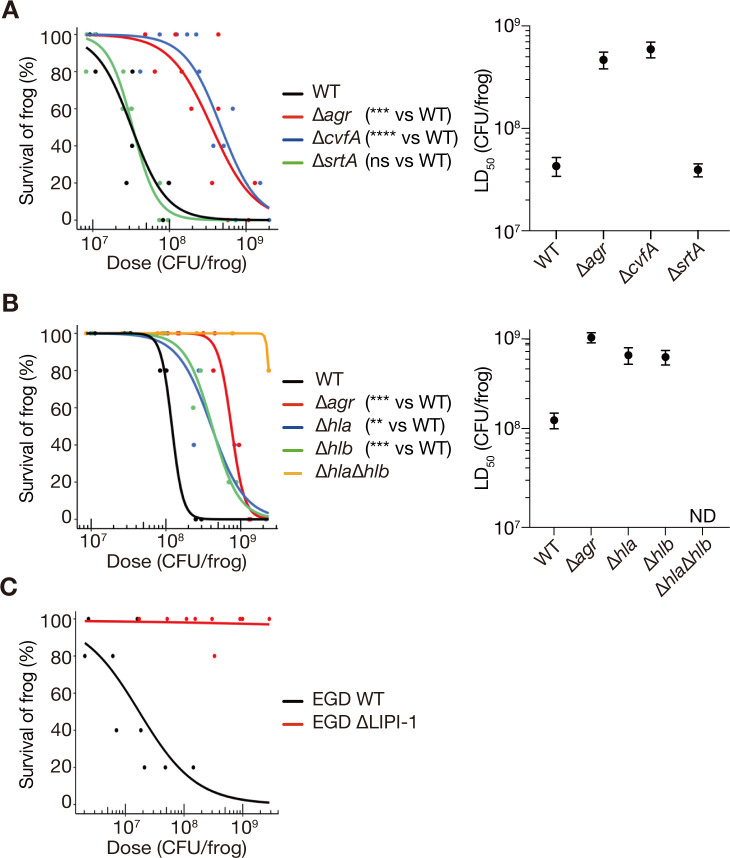 Fig 4
Fig 4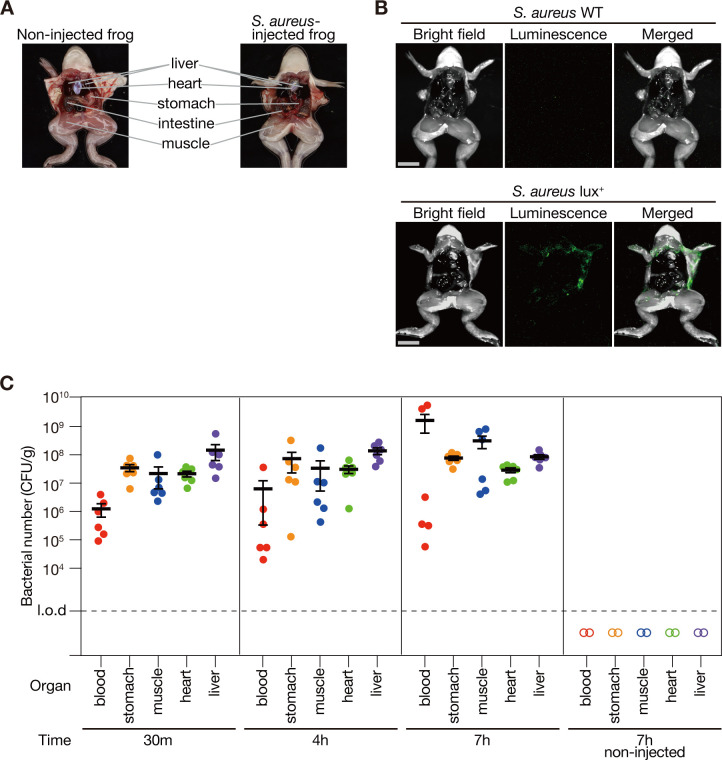 Fig 5
Fig 5| Strain or plasmid | Genotypes or characteristics | Source or reference |
|---|---|---|
| Strains | ||
| | ||
| NCTC8325-4 | NCTC8325 cured of phages | ( |
| MRSA8 | Clinical isolate, methicillin-resistant | ( |
| CKP1129 | NCTC8325-4 | ( |
| M1844 | NCTC8325-4 | ( |
| M2316 | NCTC8325-4 | ( |
| M1007 | NCTC8325-4 Δ | ( |
| M1811-1 | NCTC8325-4 Δ | ( |
| M1007-1 | NCTC8325-4 Δ | ( |
| NCTC8325-4-lux | NCTC8325-4 pND50-gmkp1-lux | This study |
| | ||
| PAO1 | Laboratory strain | ( |
| BAA-2114 | Clinical isolate, multi-drug resistant | ATCC |
| | ||
| EGD | serotype 1/2 a | ( |
| EGDLIP | EGD ΔLIPI-1::Ermr | This study |
| | ||
| BW25113 | NBRP | |
| | ||
| 168 |
| BGSC |
| Plasmids | ||
| pND50 | An integration vector, Cmr | ( |
| pHS-MCS | An integration vector, Ermr | ( |
| Primer | Target | Sequence (5'−3') |
|---|---|---|
| prfA_DS_KpnI |
|
|
| prfA_DS_XhoI |
| |
| plcB_DS_XhoI |
|
|
| plcB_DS_XbaI |
| |
| agrA_R |
|
|
| hld_R |
| |
| srtA_F |
|
|
| srtA_R |
| |
| cvfA_F |
|
|
| cvfA_R |
| |
| hla_F |
|
|
| hla_R |
| |
| hlb_F |
|
|
| hlb_R | CCCATGGCTTAGGTTTTTCA | |
| pCK20_F | pCK20 |
|
| pCK20_R |
| |
| pSF151_F | pSF151 |
|
| pSF151_R |
| |
| LIPI-1_F_close | LIPI-1 |
|
| LIPI-1_R_close |
| |
| lux-F |
| |
| lux-R |
|
- —Japan Society for the Promotion of Sciencehttp://dx.doi.org/10.13039/501100001691
- —Japan Society for the Promotion of Sciencehttp://dx.doi.org/10.13039/501100001691
- —Japan Society for the Promotion of Sciencehttp://dx.doi.org/10.13039/501100001691
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBiochemical and Structural Characterization · Yersinia bacterium, plague, ectoparasites research · Antimicrobial Resistance in Staphylococcus
INTRODUCTION
According to a 2019 report, 13.6% of global deaths were associated with infections caused by bacterial pathogens, such as Staphylococcus aureus and Pseudomonas aeruginosa (1). S. aureus is a gram-positive pathogenic bacterium that resides in the nasal cavity of approximately 30% of healthy adults and causes pneumonia, osteomyelitis, and endocarditis (1). Drug-resistant S. aureus, known as methicillin-resistant S. aureus (MRSA), is spreading worldwide, necessitating the development of new treatments (2). P. aeruginosa is a bacterium commonly found in environments such as water and soil and in the intestinal tract of healthy humans. However, during infection, it can cause urinary tract infections, sepsis, and other diseases. Most antibiotics are ineffective against infections caused by multidrug-resistant P. aeruginosa (3). Listeria monocytogenes is found in river water and animal intestinal tracts and causes meningitis, septicemia, and fetal septic granuloma in foodborne infections (4). In the United States, approximately 20% of all foodborne deaths are attributed to L. monocytogenes infections (5). To establish treatment strategies for these bacterial infections, understanding bacterial pathogenic mechanisms and the host immune system to counter the bacterial infection is crucial.
Mammalian animal models, such as mice, have traditionally been used to elucidate bacterial and host factors involved in the bacterial infection process. These models have the advantage of having immune systems similar to those of humans; however, ethical and high-cost issues limit the use of large numbers. Recently, a global movement has been initiated to reduce animal testing and use alternative methods, as evidenced by the 2013 EU ban on the sale of cosmetics tested on animals (6). Alternatives to animal testing, including in vitro and in silico experiments and the use of cultured mammalian cells, have attracted attention (7, 8); however, they fail to reproduce the complex interactions occurring in animal bodies. Animal infection models remain necessary to investigate bacterial-host interactions and disease progression leading to death.
To avoid ethical and cost problems associated with mammals, invertebrates, such as nematodes (9), fruit flies (10), silkworms (11), honey worms (12), and crickets (13), as well as vertebrate fish like zebrafish (14, 15), are being used as infection models (16, 17). These animals allow large-scale testing of bacterial pathogenicity and antimicrobial agents. However, invertebrates lack antibody-mediated adaptive immune mechanisms, making it impossible to analyze bacterial interactions with the adaptive immune system. In addition, their open vasculature, unlike vertebrates, uniquely affects the pathways for bacterial spread via body fluids. Although zebrafish possess immune systems relatively similar to those of humans (14, 15), their respiratory system, which relies on gill breathing, differs significantly from that of humans.
Establishing a non-mammalian infection model with new advantages is expected to advance research on infectious diseases. Xenopus laevis, an amphibian vertebrate, has been widely used in developmental biology for several decades (18). Xenopus laevis harbors organs, organ arrangements, and circulatory systems similar to those of humans (19). Moreover, its immune system closely resembles that of humans and possesses key factors of innate and acquired immunity, including macrophages, neutrophils, B cells, T cells, immunoglobulins, complement systems, and antimicrobial peptides (20, 21). These characteristics suggest that X. laevis could be useful for analyzing bacterial dissemination and immune interaction comparable with those in humans. The entire genome sequence of X. laevis has been completed (22), and genome editing using TALEN and CRISPR Cas9 is now possible (23–25). Xenopus frogs can be kept at high densities in aquariums without aeration because of their lung breathing (26). Fifty juvenile Xenopus frogs, each approximately 3 cm in length, can be maintained in approximately 10 L of water. In addition, they can be easily injected using human clinical tuberculin syringes without anesthetic treatment, as ice-cold water is effective. These features provide marked advantages in experiments that require a large number of animals to test using injections. Xenopus frogs have been used as an infection model for Mycobacterium marinum (19), which infects various aquatic organisms. However, they have not yet been employed as infection models for human pathogenic bacteria. Furthermore, no reports have demonstrated the use of Xenopus frogs to evaluate bacterial virulence genes or antimicrobial agent efficacy. In this study, we aimed to establish a Xenopus frog infection model for human pathogenic bacteria.
Animal infection models should be effective for evaluating bacterial virulence genes. The agr locus (27) and srtA (28) and cvfA (rny) genes (29) encode important S. aureus virulence factors. The agr locus encodes quorum sensing-related proteins and regulates toxin and cell surface protein expression (30). The srtA gene is required to anchor cell wall proteins involved in bacterial adhesion to host cells (31). The cvfA gene encodes RNase Y, which is essential for hemolysin production (29, 32, 33). Similarly, Listeria Pathogenicity Island 1 (LIPI-1) region in L. monocytogenes (34) encodes prfA, plcA, hly, mpl, actA, and plcB (35, 36). Hemolysin LLO (hly) (37) and phospholipase C (PLC) (plcA and plcB) (38) facilitate L. monocytogenes escape from phagosomes or autophagy (35, 39). These virulence genes of S. aureus and L. monocytogenes are crucial for bacterial pathogenicity in animal models such as mice (40–43), rabbits (44), crickets (13), silkworms (28, 42), and nematodes (45, 46). Therefore, these bacterial genes are suitable for examining the effectiveness of novel animal infection models to evaluate bacterial virulence. Animal infection models should also be effective for assessing the efficacy of antimicrobial agents. In this study, methicillin-susceptible and methicillin-resistant S. aureus strains, and drug-susceptible and multidrug-resistant P. aeruginosa strains were used to evaluate the efficacy of clinically used antimicrobial agents.
RESULTS
Intraperitoneal injection of S. aureus can kill Xenopus frogs
To determine whether Xenopus frogs are susceptible to human pathogenic bacteria, bacterial suspensions of S. aureus were intraperitoneally injected into the Xenopus frogs. The frogs died within 24 h post-injection, and their entire body exhibited reddening (Fig. 1A).
*Intraperitoneal injection of S. aureus is lethal to Xenopus frogs. (A) Intraperitoneal injection of frogs is shown (left panel). Frogs injected with S. aureus NCTC8325-4 strain (2.8 × 108 CFU/frog) were dead within 24 h post-injection (right panel). Frogs injected with PBS survived (center panel). (B) Time-course survival analysis of frogs intraperitoneally injected with varying doses of S. aureus NCTC8325-4 strain. Five frogs were injected per sample, and survival data were pooled from four independent experiments (n = 20). ns: P > 0.05, **: P < 0.01, ***: P < 0.0001. (C) Dose-response survival curve of frogs injected with live or heat-killed S. aureus NCTC8325-4. Overnight culture of S. aureus was autoclaved at 121°C for 20 min before use as heat-killed bacteria. The experiment was performed using at least three serial dilutions of bacterial solution and independently performed three times. The data point indicates the survival rate of five frogs injected with a specific dose of bacteria. Logistic regression analysis was performed using all survival data points to calculate LD50.
To assess whether mortality was dependent on the number of injected bacteria, frogs were injected with varying concentrations of S. aureus suspension, and their survival rate was observed over time. Mortality was observed between 12 and 36 h after injection, which increased with higher bacterial concentrations (Fig. 1B). These results indicated that S. aureus infection leads to frog death in a dose-dependent manner.
To confirm that the death of frogs post-injection was caused by live bacteria, we examined whether heat-killed S. aureus exhibited similar lethal activity. The survival rate of frogs injected with live S. aureus decreased, depending on the number of bacteria at 24 h post-injection, whereas that of frogs injected with heat-killed S. aureus did not decrease at 24 h post-injection (Fig. 1C). The median lethal dose (LD_50_) of live S. aureus was 3.2 × 10^7^ CFU/frog, whereas that of heat-killed S. aureus exceeded 7.1 × 10^9^ CFU/frog. These results suggested that frogs died owing to the biological activity of viable S. aureus, such as bacterial growth and toxin production.
Various human pathogenic bacteria can kill Xenopus frogs
To determine whether the frog-killing ability is conserved among human pathogenic bacteria other than S. aureus NCTC8325-4, we examined the virulence of other S. aureus strains, P. aeruginosa, and L. monocytogenes against Xenopus frogs. Intraperitoneal injection of MRSA (MRSA8), P. aeruginosa strain PAO1, multidrug-resistant P. aeruginosa (BAA-2114), and L. monocytogenes strain EGD caused frog death based on the bacterial number (Fig. 2A through D), similar to S. aureus NCTC8325-4 (Fig. 1C). Additionally, we examined whether laboratory strains of non-pathogenic bacteria such as E. coli (47) and B. subtilis (48) induce mortality. Intraperitoneal injections of E. coli BW25113 and B. subtilis 168 showed little killing activity in frogs within the tested bacterial concentrations (Fig. 2E and F). LD_50_ calculated from survival curves showcased that P. aeruginosa BAA-2114 had the highest LD_50_ of 3.8 × 10^9^ CFU/frog, whereas S. aureus NCTC8325-4 and L. monocytogenes EGD exhibited lower LD_50_ values (< 4.3 × 10^7^ CFU/frog) (Fig. 2G). LD_50_ values of E. coli BW25113 and B. subtilis 168 were not determined, as they exceeded 3.3 × 10^9^ and 1.6 × 10^9^ CFU/frog, respectively (Fig. 2G). These results suggested that nonpathogenic bacteria have limited lethality in frogs, whereas human pathogenic bacteria demonstrate frog-killing ability, which varies among bacterial species and strains.
Various species of human pathogenic bacteria induce mortality in Xenopus frogs. (A–F) Dose-response survival curves of frogs injected intraperitoneally with S. aureus MRSA8 (A), P. aeruginosa PAO1 (B), P. aeruginosa BAA-2114 (C), L. monocytogenes EGD (D), E. coli BW25113 (E), and B. subtilis 168 (F) generated from the survival rate at 24 h post-injection. Five frogs were injected with three different concentrations of bacterial suspensions in a single experiment, and survival data obtained from at least three independent experiments are shown. (G) Based on the survival data (A–F), logistic regression analysis was performed using the statistical software R to determine LD50. Error bars indicate standard error. LD50 for NCTC8325-4 was calculated from the survival data in Fig. 1C. ND indicates not determined.
Evaluation of antibacterial drug efficacy in Xenopus
To determine whether Xenopus frogs can serve as a model for evaluating antimicrobial efficacy, we examined the therapeutic effects of antimicrobials on frog death induced by human pathogenic bacteria. First, the toxicities of kanamycin (KM), oxacillin (OX), vancomycin (VCM), ciprofloxacin (CPFX), and ceftazidime (CAZ) were tested, confirming that none of the tested antibiotics caused frog mortality at the antimicrobial concentrations used in this study (Table S1). Antimicrobials were administered immediately after bacterial injection, and frogs were considered recovered if their survival rate exceeded 50% at 120 h post-infection.
MRSA8, an MRSA strain of S. aureus, is resistant to oxacillin (β-lactam) and kanamycin (aminoglycoside) but remains sensitive to vancomycin (11, 13, 49). BAA-2114 is a multidrug-resistant P. aeruginosa isolated from the sputum samples of patients with cystic fibrosis (50), which displays resistance to kanamycin, ceftazidime (β-lactam), and ciprofloxacin (fluoroquinolones) (51–53).
Frogs injected with NCTC8325-4, a methicillin-susceptible strain of S. aureus, without antibacterial injections exhibited a survival rate of less than 10%, whereas the administration of KM, OX, or VCM after injecting bacterial suspension resulted in survival rates of >50% at 120 h post-infection (Fig. 3A). Frogs injected with S. aureus MRSA8 had 0% survival at 120 h post-infection, whereas those injected with VCM after MRSA8 achieved a 100% survival rate in the same time (Fig. 3B). In contrast, injecting KM or OX following MRSA8 resulted in nearly 0% survival at 120 h post-infection (Fig. 3B). These results indicated that NCTC8325-4 infection in frogs can be treated with KM, OX, and VCM, whereas MRSA8 infection responds only to VCM.
*Antimicrobial agents inhibit bacterial infections in Xenopus frogs. Therapeutic effects of kanamycin (KM), oxacillin (OX), and vancomycin (VCM) on frogs injected with S. aureus NCTC8325-4 (A) and MRSA8 (B) were examined. Similarly, the effects of kanamycin (KM), ciprofloxacin (CPFX), and ceftazidime (CAZ) on frogs infected with P. aeruginosa PAO1 (C) and BAA-2114 (D) were examined. Immediately after intraperitoneal injection of bacterial suspensions, frogs were intraperitoneally administered with 50 µL of 2 mg/mL antimicrobial agent (100 µg/frog) or PBS, and survival was measured every 12 h. Five frogs were used per agent, and survival data were pooled from three independent experiments (n = 15). Significant differences between PBS- and antimicrobial-injected groups were determined using the log-rank test (**P < 0.01, ***P < 0.001, ***P < 0.0001).
Frogs injected with P. aeruginosa PAO1 without antimicrobials showed 0% survival at 120 h post-infection, whereas those treated with KM, CPFX, or CAZ following PAO1 administration showed survival rates of >50% (Fig. 3C). Similarly, frogs injected with P. aeruginosa BAA-2114 without antimicrobial agents were 0% at 120 h post-infection, whereas those injected with KM, CPFX, or CAZ after BAA-2114 showed less than 50% survival rates in the same timeframe (Fig. 3D). These findings suggested that PAO1 infections in frogs can be effectively treated with KM, CPFX, or CAZ, whereas BAA-2114 infections are refractory to these antimicrobial agents.
Evaluation of bacterial virulence genes
To determine whether the virulence genes of pathogenic bacteria could be evaluated using Xenopus frogs, virulence gene deletion mutants of S. aureus and L. monocytogenes were intraperitoneally injected into Xenopus frogs, and their viability was measured 24 h post-infection. S. aureus NCTC8325-4 mutants with deleted agr locus (27), srtA (28), and cvfA (29) killed frogs in a dose-dependent manner (Fig. 4A). The LD_50_ values of agr- and cvfA-deleted mutants were more than 10-fold higher than those of the wild-type strain (Fig. 4A). In contrast, the LD_50_ of srtA-deleted mutant was nearly the same as that of the wild-type strain (Fig. 4A). These results suggested that agr and cvfA contribute to S. aureus virulence against Xenopus frogs, whereas srtA does not.
*Deletion of bacterial virulence genes reduces bacterial killing activity against Xenopus frogs. (A) Comparison of the frog-killing abilities of S. aureus NCTC8325-4 strain (WT) and strains with deleted agr, srtA, and cvfA. (B) Comparison of the frog-killing abilities of NCTC8325-4 WT and strains with deleted agr, hla, hlb, and hla/hlb. (C) Comparison of the frog-killing ability of L. monocytogenes EGD (WT) and a strain with deleted LIPI-1. The data for L. monocytogenes EGD WT are the same as those presented in Fig. 2D. For all panels, bacterial suspensions were injected intraperitoneally into frogs, and survival was measured at 24 h post-injection. Five frogs were injected with each of three different concentrations of bacterial suspensions per strain in a single experiment. Dose-response survival curves were generated using logistic regression based on data from at least three independent experiments, and LD50 and standard error were calculated. Significant differences between gene-deleted and WT strains were determined using the likelihood ratio test (**: P < 0.01, ***: P < 0.001, ***: P < 0.0001). Error bars represent standard error.
Given that the agr- and cvfA-deleted mutants attenuated virulence against Xenopus frogs and agr and cvfA are required for hemolysin production (30, 54), we hypothesized that the reduced hemolysin production in these mutants may contribute to their attenuated virulence. To verify this, we assessed the virulence of hemolysin gene deletion mutants in Xenopus frogs. Deletion mutants of hla (encoding α-hemolysin) and hlb (encoding β-hemolysin) are known to reduce virulence in mammals (28, 44). The LD_50_ values of hla- and hlb-deleted mutants were higher than those of the wild-type strain (Fig. 4B). In addition, the LD_50_ of the hla/hlb double deletion mutant exceeded 2.4 × 10^9^ CFU/frog, which was higher than the LD_50_ values of mutants harboring either deletion (Fig. 4B). These results suggested that hla and hlb act in an additive manner to kill Xenopus frogs.
Xenopus frogs injected with the LIPI-1-deleted mutant of L. monocytogenes EGD showed minimal mortality at 24 h post-infection (Fig. 4C). The LD_50_ of wild-type strain was 3.6 × 10^7^ CFU/frog, whereas that of the LIPI-1-deleted mutant was greater than 2.8 × 10^9^ CFU/frog ([Fig. 2G and 4C](#F2 F4)). These results suggested that the LIPI-1 locus of L. monocytogenes is essential for its virulence against Xenopus frogs.
Intraperitoneal injection of S. aureus into Xenopus frogs causes bacterial dissemination
We examined the visceral morphology of Xenopus frogs intraperitoneally injected with S. aureus NCTC8325-4 and compared it with that of non-injected frogs (Fig. 5A). The stomachs of non-injected frogs were pale pink, and the muscles were soft and movable. In contrast, the stomachs of injected frogs, examined immediately after death at 7 h post-injection, were red and deformed, and rigor mortis had set in, with hard and immobile muscles (Fig. 5A). In addition, blood obtained by amputating the upper legs of non-injected frogs was dark red, whereas that of infected frogs immediately after death appeared light red.
Bacteria disseminate from the intraperitoneal cavity to the whole body in Xenopus frogs. (A) Photograph of a Xenopus frog with its abdomen open. The left shows a non-injected frog. The right shows a frog that died 7 h after intraperitoneal injection of S. aureus NCTC8325-4 strain (8.6 × 108 CFU/frog). (B) S. aureus NCTC8325-4 strain (WT) or NCTC8325-4 strain carrying pND50-gmkp1-lux was injected intraperitoneally into the frog, and the abdomen was opened 7 h post-injection to observe luminescence. Bright field (left), luminescence (center), and merged images (right) are displayed. Scale bar: 10 mm. The bacterial dose administered was 8.6 × 108 CFU/frog. (C) Xenopus frogs were intraperitoneally injected with S. aureus NCTC8325-4 strain (8.6 × 108 CFU/frog), and bacterial CFUs were quantified in blood and organs at 30 min, 4 h, and 7 h post-injection. At each time point, three animals were analyzed in a single experiment, and two independent experiments were performed (n = 6). Mean CFU count from six frogs at each time point is represented by a solid horizontal bar, with the error bar indicating standard error. Open symbols indicate data from PBS-injected control frogs (n = 2). A dashed line indicates a limit of detection (liver, stomach, and muscle, 1 × 103 CFU/g; heart, 4 × 103 CFU/g; blood, 2 × 102 CFU/g).
To investigate the distribution of S. aureus in Xenopus frogs after intraperitoneal injection, frogs were intraperitoneally injected with an S. aureus strain expressing the lux operon, which enables a luminescence reaction without the need for external substrates. Seven hours post-infection, luminescence was observed at the periphery of the abdominal cavity (Fig. 5B). Conversely, no luminescence was detected in frogs administered with the S. aureus strain lacking the lux operon (Fig. 5B). These results suggested that S. aureus intraperitoneally injected into Xenopus frogs disseminated throughout the abdominal cavity.
To further investigate the systemic distribution of S. aureus in Xenopus, Xenopus frogs were intraperitoneally injected with S. aureus and dissected at 30 min, 4 h, and 7 h post-injection to quantify bacterial counts in the blood and various organs (stomach, leg muscle, heart, and liver). S. aureus was detected at all time points and in all examined organs, with bacterial numbers maintained from 30 min to 7 h post-infection (Fig. 5C). In contrast, S. aureus was not detected in the organs of the frogs injected with PBS (Fig. 5C). Therefore, all detected S. aureus were derived from the intraperitoneally injected bacteria. Notably, S. aureus was observed in the blood and in the leg muscles that do not directly contact the abdominal cavity, suggesting that intraperitoneally injected S. aureus spreads throughout the body via the bloodstream.
DISCUSSION
Our results indicate that Xenopus frogs are susceptible to intraperitoneal injection of the human pathogenic bacteria, S. aureus, P. aeruginosa, and L. monocytogenes and lead to mortality. Furthermore, clinically effective antimicrobial agents suppressed frog death infected with S. aureus and P. aeruginosa, whereas bacterial mutant strains with deleted virulence genes in S. aureus and L. monocytogenes exhibited attenuated virulence. This study is the first to suggest that Xenopus frogs are a viable animal infection model for evaluating both the efficacy of antimicrobial agents and the virulence gene function of human pathogenic bacteria.
This study showed that S. aureus disseminated into the blood and organs of Xenopus frogs following intraperitoneal injection, resulting in systemic infections. In humans, intraperitoneal infections caused by laparotomy or intestinal obstruction can progress to sepsis (55, 56). In addition, intraperitoneally injected bacteria have been detected in the blood and liver of the mouse infection model (57, 58). These observations suggest that the intraperitoneal bacterial infection in Xenopus frogs can simulate bacterial sepsis in mammals, where intraperitoneally infected bacteria disseminate throughout the body.
In this study, we found that hla, hlb, as well as agr and cvfA, which regulate the transcriptional expression of hla and hlb, are required for the lethality of S. aureus in Xenopus frogs (Fig. 4A and B). S. aureus strains lacking hla and hlb showcase reduced virulence in rabbits (44). In human alveolar epithelial cells, α-hemolysin interacts with the host cell surface protein ADAM10 and forms a heptameric β-barrel structure, creating pores in the cell membrane (59). ADAM10 knockout in mouse alveolar epithelial cells confers resistance to lethal pneumonia caused by S. aureus (60). β-Hemolysin is a neutral sphingomyelinase that degrades sphingomyelin in cell membranes (61). According to Xenbase, a database of X. laevis and X. tropicalis, X. laevis has ADAM10 (target of α-hemolysin) and a sphingomyelin synthase (target of β-hemolysin), suggesting that α-hemolysin and β-hemolysin may exert similar effects in X. laevis as in mammals. These findings support the use of Xenopus frogs for analyzing bacterial virulence mechanisms mediated by α- and β-hemolysins.
During intraperitoneal infection in Xenopus frogs, the srtA-deleted mutant in this study showed virulence comparable with that of the wild-type S. aureus (Fig. 4A). In rats and mice, the pathogenicity of srtA-deleted mutants is lower than that of the wild-type strain administered via various methods, including intraperitoneal injection (62–64). Similarly, in silkworms and crickets, the srtA-deleted mutant is less virulent than the wild-type strain in systemic infection models induced by intrahemolymph injection (13, 28). In contrast, in the nematode infection model, where bacteria were fed to the nematodes, srtA-deleted mutant displayed similar pathogenicity to that of the wild-type strain (65, 66). As srtA deficiency attenuates virulence in mice following intraperitoneal injection (62), this suggests that the non-attenuated virulence of srtA-deleted mutants observed in Xenopus frogs is not due to the infection route, but rather to host animal-specific differences. The srtA gene is involved in anchoring more than 10 different cell wall adhesion proteins (67, 68). The molecular mechanisms underlying why cell adhesion by these cell wall proteins is not required for intraperitoneal infection in Xenopus frogs need to be analyzed in the future.
The mortality of Xenopus frogs caused by drug-susceptible and drug-resistant strains of S. aureus and drug-susceptible strains of P. aeruginosa was suppressed by the administration of appropriate antimicrobial agents. However, multidrug-resistant P. aeruginosa remained lethal despite antimicrobial treatment (Fig. 3). Hence, Xenopus frogs could prove useful for evaluating antibiotic efficacy against sepsis caused by S. aureus and P. aeruginosa. Xenopus juvenile frog is approximately 2.5 g, making it smaller than mice (approximately 20–30 g in weight), which allows for the evaluation and exploration of antimicrobial agents using approximately 10-fold smaller drug doses. For example, previous studies demonstrated that intraperitoneal injection of 1 mg vancomycin every 12 h increased the survival rates of mice infected with MRSA (69, 70), whereas this study revealed that only once intraperitoneal injection of 0.1 mg vancomycin increased the survival rates of Xenopus frogs infected with MRSA. In this study, antimicrobials were intraperitoneally injected immediately after injecting bacterial suspensions. Future studies should evaluate the efficacy of other antimicrobial administration routes used in clinical practice, such as oral and subcutaneous administration. The Xenopus infection model has the potential for identifying and analyzing key human pathogenic bacterial genes involved in sepsis progression and evaluating therapeutic interventions.
MATERIALS AND METHODS
X. laevis
X. laevis juvenile frogs were purchased from a breeding company (Xenopus Yoshoku Kyozai, Ibaraki, Japan). The frogs were 3–4 cm in size and weighed approximately 2.5 g. Fifty frogs were housed in a 13.6 L (37 cm W × 22 cm D × 25 cm H) aquarium, which was half-filled with water. The aquarium was maintained at a room temperature of 22°C. The frogs were fed with dried goldfish food (Cat. 4971453050347, Itosui, Tokyo, Japan) every 2 days. The rearing water was replaced with fresh dechlorinated water once every 2 days (71).
Bacterial strains
All bacterial strains used in this study are listed in Table 1. S. aureus gene deletion mutants for agr locus (72), srtA (28), cvfA (29), hla (28), hlb (28), and hla/hlb (28) were constructed in our previous studies. We confirmed the deletion of the respective genes by PCR, using genomic DNA as the template and oligonucleotide primers (Table 2). L. monocytogenes mutant strain lacking LIPI-1 was constructed according to a previously described method (73). The downstream regions of prfA and plcB were amplified via PCR and ligated into pHS-MCS. L. monocytogenes EGD strain was electroporated with the obtained plasmids, resulting in a LIPI-1-deleted mutant. The genomic DNA of L. monocytogenes was extracted using the QIAamp Blood Mini Kit (QIAGEN, Venlo, Netherlands) as previously described (74), and LIPI-1 deletion was confirmed through PCR.
Preparation of bacterial solution
A single S. aureus colony on tryptic soy agar was inoculated into 5 mL of tryptic soy broth medium and aerobically incubated at 37°C overnight. The overnight culture was centrifuged at 10,400 × g for 10 min, and the bacterial pellet was resuspended in phosphate-buffered saline (PBS) and diluted to prepare bacterial solutions at appropriate concentrations. The same procedure was performed for P. aeruginosa and L. monocytogenes, using lysogeny broth and brain heart infusion media, respectively. In multiple independent experiments, the optical density at 600 nm (OD_600_) of overnight cultures was measured, and a calibration curve was generated to determine the relationship between CFU and OD_600_. This calibration curve was used to estimate the CFU injected into frogs based on the OD_600_ of overnight cultures in the infection experiments.
Preparation of antimicrobial solutions
Antimicrobial solutions were prepared as follows: 10 mg each of kanamycin monosulfate (TCI, Tokyo, Japan), oxacillin (SIGMA, St. Louis, MO), vancomycin hydrochloride (FUJIFILM, Tokyo, Japan), and ceftazidime hydrate (LKT, St. Paul, MN) was dissolved in 5 mL PBS to achieve a concentration of 2 mg mL^−1^. Ciprofloxacin hydrochloride monohydrate (TCI, Tokyo, Japan) was dissolved in Milli-Q water to a concentration of 2 mg mL^−1^. All antimicrobial solutions were stored at −30°C and thawed before use.
Intraperitoneal injections
Xenopus frogs were anesthetized by placing them on crushed ice for 2 min. A 50 µL bacterial solution was injected into the abdominal cavity of X. laevis, just above the left leg, using a 27G needle (outer diameter) of 3/4-inch length (Terumo, Tokyo, Japan) (Fig. 1A) attached to a 1 mL tuberculin syringe (Terumo, Tokyo, Japan). Five animals per sample were injected, placed in a 1 L beaker containing 500 mL water, and kept at 27°C in the dark. The frogs recovered from cold anesthesia and began swimming immediately after being placed in the beaker. To exclude the possibility of death due to low-temperature anesthesia or injection-related stress, a negative control group injected with PBS was prepared for all experiments.
Calculation of LD50
Three different concentrations of bacterial solutions were employed for each experiment. Overnight cultures were centrifuged at 10,400 × g for 10 min, the bacterial pellet was resuspended in PBS, and a 3-fold dilution series was prepared using PBS. Five animals were injected per sample, and survival rates were recorded at 24 h post-infection. The experiment was performed at least three independent times, and the survival rate data and injected CFU were plotted on a graph. A dose-response survival curve was created using logistic regression using the statistical software R to obtain LD_50_ and standard error. Survival data for each experiment are presented in Table S1.
Luminescence measurement
DNA fragments containing the lux operon were amplified by PCR using genomic DNA from S. aureus Xen29 (80) as a template and a primer pair (Table 2). The amplified DNA was inserted into the BamHI and PstI recognition sites of a plasmid carrying the gmk promoter region (81), resulting in pND50-gmkp1-lux. S. aureus RN4220 strain was transformed with pND50-gmkp1-lux through electroporation, and the plasmid was subsequently transferred into NCTC8325-4 strain using phage 80α via transduction. The NCTC8325-4 strain carrying pND50-gmkp1-lux was intraperitoneally injected into Xenopus frogs. At 7 h post-injection, the abdomen was opened, and chemiluminescence was measured using a CCD camera (iBright 1500, Thermo Fisher, Waltham, MA). The camera was configured to an exposure time of 10 min, zoom 3.0, and focus 390. Three images were captured per animal under the same conditions, and the average function of ImageJ was used to compute the mean luminescence intensity from the three photographs.
Measurement of bacterial count in organs
Xenopus frogs were intraperitoneally injected with S. aureus NCTC8325-4. At 30 min, 4 h, and 7 h post-injection, the frogs were anesthetized by immersing them in 2 g L^−1^ ethyl m-aminobenzoate methanesulfonate solution (Nacalai Tesque, Kyoto, Japan) for 5 min. The right upper leg was amputated, and blood was collected. Muscles of the right upper leg, stomach, heart, and liver were excised. To remove residual blood, the organs other than blood were washed with PBS, weighed, and then homogenized in 400 µL PBS using TissueLyser II (QIAGEN) at 25 Hz for 1.5 min. A 10-fold dilution series of blood and organ homogenates was prepared using 0.9% NaCl. Approximately 5 µL of each dilution was spread onto mannitol salt agar plates, a selective medium for S. aureus, and incubated at 37°C overnight. The colonies were counted according to a previously defined method (82). Bacterial numbers per gram of blood and organ were calculated based on colony counts and dilution ratios.
Statistical analysis
The log-rank (Mantel-Cox) test was performed using GraphPad Prism software (ver. 10.2.3) to determine significant differences in the time-survival curves. A dose-response survival curve was generated via logistic regression to plot frog survival against CFU injected into frogs using R for Windows (ver. 4.3.3). The LD_50_ and standard error were estimated using the glm function from the MASS library (83). A likelihood ratio test was conducted using the anova function to determine significant differences between the two dose-response survival curves. The R code used for these analyses is registered in the GitHub repository (doi: 10.5281/zenodo.14978657).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ikuta KS, Swetschinski LR, Aguilar GR, Sharara F, Mestrovic T, Gray AP, Weaver ND, Wool EE, Han C, Hayoon AG. 2022. Global mortality associated with 33 bacterial pathogens in 2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet 400:2221–2248. doi:10.1016/S 0140-6736(22)02185-736423648 PMC 9763654 · doi ↗ · pubmed ↗
- 2Turner NA, Sharma-Kuinkel BK, Maskarinec SA, Eichenberger EM, Shah PP, Carugati M, Holland TL, Fowler VG. 2019. Methicillin-resistant Staphylococcus aureus: an overview of basic and clinical research. Nat Rev Microbiol 17:203–218. doi:10.1038/s 41579-018-0147-430737488 PMC 6939889 · doi ↗ · pubmed ↗
- 3Pachori P, Gothalwal R, Gandhi P. 2019. Emergence of antibiotic resistance Pseudomonas aeruginosa in intensive care unit; a critical review. Genes Dis 6:109–119. doi:10.1016/j.gendis.2019.04.00131194018 PMC 6545445 · doi ↗ · pubmed ↗
- 4Schlech WF. 2019. Epidemiology and clinical manifestations of Listeria monocytogenes infection. Microbiol Spectr 7. doi:10.1128/microbiolspec.gpp 3-0014-2018 PMC 1102608231837132 · doi ↗ · pubmed ↗
- 5Scallan E, Hoekstra RM, Angulo FJ, Tauxe RV, Widdowson M-A, Roy SL, Jones JL, Griffin PM. 2011. Foodborne illness acquired in the United States--major pathogens. Emerg Infect Dis 17:7–15. doi:10.3201/eid 1701.p 1110121192848 PMC 3375761 · doi ↗ · pubmed ↗
- 6Petetta F, Ciccocioppo R. 2021. Public perception of laboratory animal testing: historical, philosophical, and ethical view. Addict Biol 26:e 12991. doi:10.1111/adb.1299133331099 PMC 9252265 · doi ↗ · pubmed ↗
- 7Štampar M, Žegura B. 2024. In vitro hepatic 3D cell models and their application in genetic toxicology: a systematic review. Mutat Res Genet Toxicol Environ Mutagen 900:503835. doi:10.1016/j.mrgentox.2024.50383539617595 · doi ↗ · pubmed ↗
- 8Jean-Quartier C, Jeanquartier F, Jurisica I, Holzinger A. 2018. In silico cancer research towards 3R. BMC Cancer 18:408. doi:10.1186/s 12885-018-4302-029649981 PMC 5897933 · doi ↗ · pubmed ↗