Colonization dynamics of Streptococcus pneumoniae are determined by polymorphisms in the BlpAB transporter
Surya D. Aggarwal, Jacqueline Toussaint, John A. Lees, Jeffrey N. Weiser

TL;DR
This study shows that Streptococcus pneumoniae strains with a functional BlpAB transporter colonize human airways longer and compete better, explaining why this transporter persists despite being redundant.
Contribution
The study identifies the evolutionary advantage of BlpAB(+) Streptococcus pneumoniae strains through their impact on colonization dynamics and competitive success.
Findings
BlpAB(+) strains exhibit slower loss of clonal diversity during colonization in mice.
BlpAB(+) strains have a median carriage duration ~177 days longer in human populations.
Functional BlpAB provides a competitive advantage during co-colonization experiments.
Abstract
Colonization of the human airways, the first step in the pathogenesis of Streptococcus pneumoniae (Spn), is the determining factor in the ecological spread of the bacterium. Since co-colonization by multiple strains is common, within-host bacterial competition contributes to the success of Spn strains. Competition both between and within strains is mediated by bacteriocin gene clusters, notably the quorum sensing-regulated bacteriocin-like peptide (blp) locus. A key component of this system is the BlpAB transporter that exports pheromones and bacteriocins expressed by the blp locus. However, ~75% of Spn strains lack a functional BlpAB transporter and instead rely on the paralogous ComAB transporter for this export, raising questions about the evolutionary persistence of BlpAB(+) strains. Using molecular barcoding, we demonstrate that BlpAB(+) and BlpAB(−) strains show major differences…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
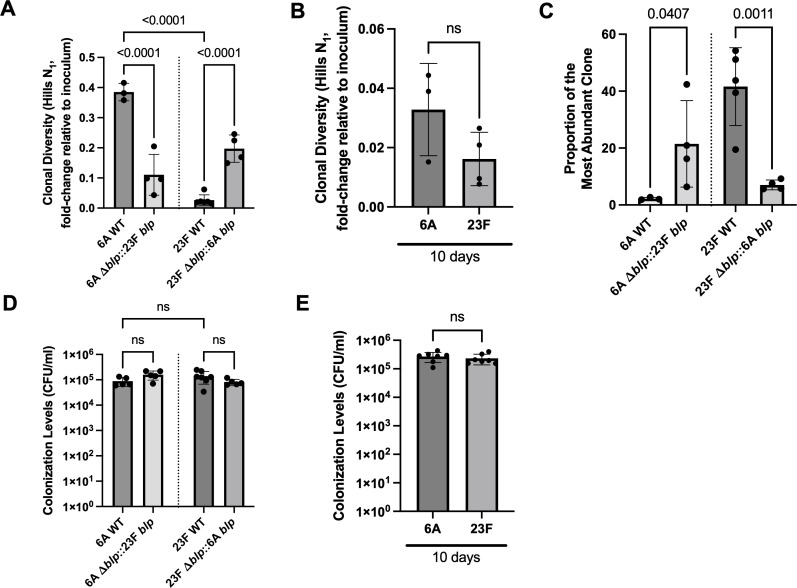 Fig 1
Fig 1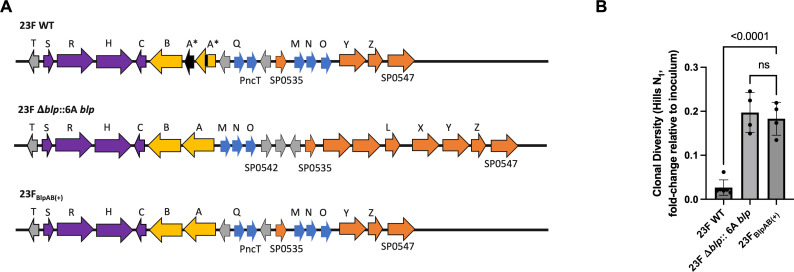 Fig 2
Fig 2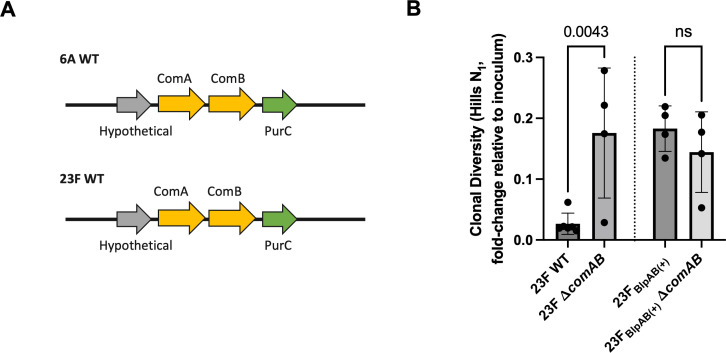 Fig 3
Fig 3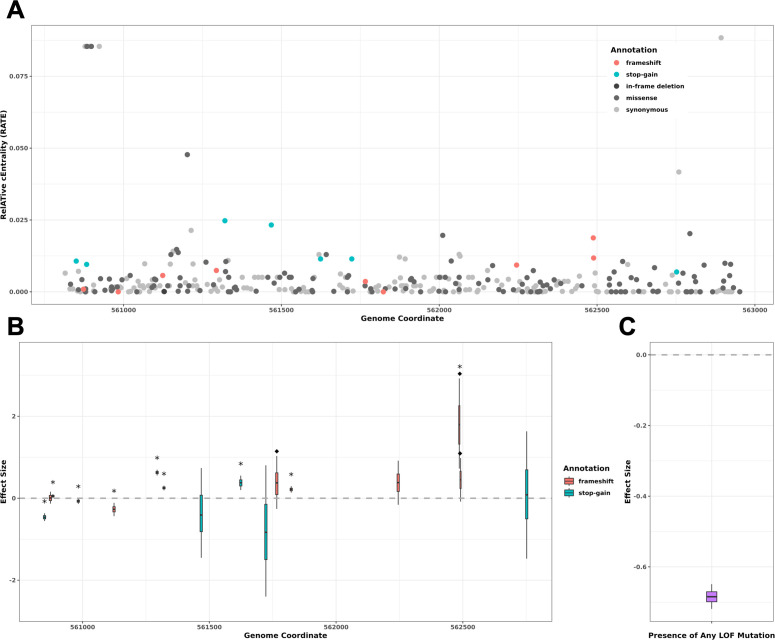 Fig 4
Fig 4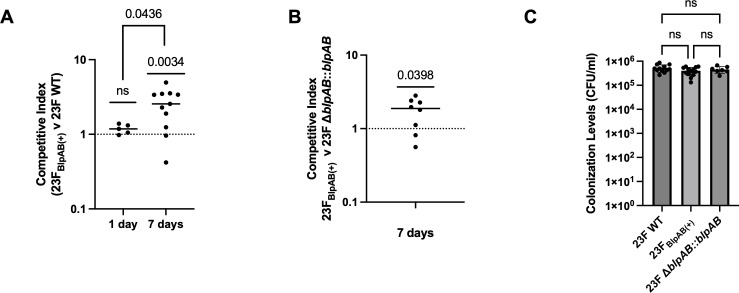 Fig 5
Fig 5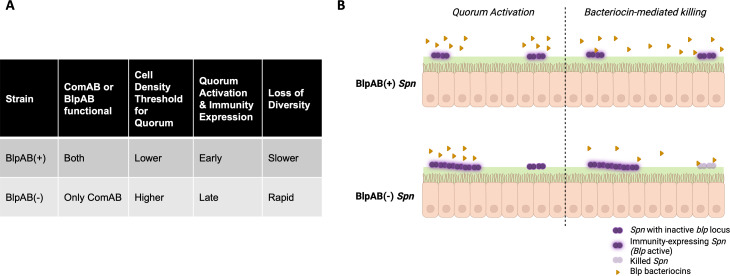 Fig 6
Fig 6- —National Institute of Allergy and Infectious Diseaseshttp://dx.doi.org/10.13039/100000060
- —National Institute of Allergy and Infectious Diseaseshttp://dx.doi.org/10.13039/100000060
- —European Molecular Biology Laboratoryhttp://dx.doi.org/10.13039/100013060
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPneumonia and Respiratory Infections · Bacterial Infections and Vaccines · Streptococcal Infections and Treatments
INTRODUCTION
Streptococcus pneumoniae (Spn) remains a major human pathogen and a leading etiological agent responsible for the deaths of children below the age of 5 (1–3). Its pathogenic phase occurs when Spn invades other body sites, such as lungs and bloodstream, thereby resulting in invasive inflammatory diseases (4). Typically, Spn leads a commensal lifestyle within the human upper respiratory tract (URT), where it frequently colonizes the mucosal surfaces (5, 6). Successful colonization is not only a prerequisite for invasive disease but also drives the ecological spread of the bacterium by facilitating transmission between hosts (5).
The prevalence of Spn carriers is especially high among children, with carriage rates ranging between 23% and 65% (5, 7–9). Surveys of carriers have shown that colonization events are of variable duration, ranging from days to months. Furthermore, they have provided evidence that a single host can be co-colonized by multiple Spn strains and that these co-colonization episodes are rather frequent in settings where Spn colonization is prevalent (10, 11). In these instances, within-host bacterial competition will play a critical role in impacting Spn population structure and the ecological success of a lineage within the host population. Spn encodes numerous putative bacteriocin gene clusters that can mediate such competitive behaviors (12, 13). Of these, the bacteriocin-like peptide (blp) locus has been shown to facilitate inter- and intrastrain Spn competition in vivo (14, 15). In addition to bacteriocin peptides, the blp locus also encodes immunity proteins, regulatory components, ABC transporters, and proteins of unknown function. The immunity factors provide protection from the activity of cognate bacteriocins and are critical factors in regulating blp-mediated strain predation. Genes of the blp locus are expressed in response to signaling by the BlpC quorum sensing peptide pheromone (16).
Although ubiquitous, the blp locus is one of the most heterogenous gene clusters in Spn (17–19). The exact composition of this locus shows extensive variability at all levels of genomic organization: gene presence or absence, gene order, as well as allelic diversity (17). Interestingly, only a small minority (<25%) of Spn strains possess an intact ABC transporter, BlpAB, that can mediate export of blp peptides [BlpAB(+) strains] (17, 20). While the complete array of mutations in blpA remains uncharacterized, it was previously reported that many Spn strains [BlpAB(−) strains] carry a 4 bp duplication in blpA that renders this gene non-functional (21). Regardless, BlpAB(−) Spn strains can utilize the functionally paralogous ComAB transporter for export of the blp peptides. The expression of ComAB is induced as a part of a distinct Com quorum sensing system that regulates competence. As both BlpAB and ComAB are promiscuous in the cargo they can transport, the kinetics of quorum sensing activation in these bacteria will be impacted based on whether they possess just one or both of these transporters (20, 22). It has been proposed that ancestrally, Spn strains possessed an intact BlpAB and that the bacterium has evolved over time to possess several independent frameshift mutations in blpA generating BlpAB(−) strains (20). Given this redundancy in the peptide transport systems, it remains puzzling as to why Spn strains with an intact BlpAB are still maintained in the population at a reasonably high frequency.
In our recent work, we utilized chromosomal barcoding of otherwise isogenic bacteria to investigate the dynamics of Spn population during colonization of the URT (14). This allowed us to study the trajectory of multiple clonal lineages during infection and identify factors that influence the outcome of infection. This work provided evidence of extensive intrastrain competition in vivo that results in a rapid loss of Spn clonal diversity. Quorum sensing-dependent activation of blp bacteriocins was responsible for this loss of diversity and kin predation (14). In this work, we questioned whether studying population dynamics both in experimental and natural colonization can shed light on the paradox of BlpAB(+) strains and divulge why they are maintained in the population.
RESULTS
Spn isolates exhibit differential dynamics in the loss of diversity
To study how differences in the blp locus impact population dynamics of Spn during colonization, we utilized two Spn isolates: type 23F (Spn 23F) and type 6A (Spn 6A). The blp loci of Spn 23F and 6A are variable in a range of aspects, including in the number of bacteriocin and immunity proteins encoded and the functionality of the BlpAB transporter (23). To examine Spn’s population dynamics in vivo, we used previously constructed, molecularly barcoded libraries of these bacterial isolates. These libraries were designed to have 7 nt barcodes of the sequence NNMCAATGNNMCAAN. The IgA1 protease cleaves human but not murine IgA and has no known impact on bacterial fitness in mice (24). The barcodes are present immediately upstream of a spectinomycin resistance cassette and in the disrupted iga gene. Both Spn 23F and 6A libraries are sufficiently diverse, containing 2,764 and 3,060 uniquely barcoded clones, respectively (14, 25). Each of these libraries contains isogenic clones, differentiated only by the sequence of their respective barcodes.
Infants are a major reservoir for Spn and display higher rates of carriage (26). They are also more likely to carry multiple Spn strains simultaneously (11). Hence, we utilized an infant mouse model of colonization for our studies. Following inoculation of barcoded libraries in an infant mouse model of colonization, we observed that both Spn 23F and 6A strains show a loss of clonal diversity, measured by Hill’s N1 coefficient, at 1 day post-inoculation (dpi) (Fig. 1A). Hill’s N1 clonal diversity metric enables the comparison of barcode richness and the abundance of each barcode across samples. The presence of more unique barcodes in a sample will result in its overall diversity being higher. While the clonal diversity of the Spn 23F strain decreased to 2.67% of the inoculum at 1 dpi, the corresponding diversity of the Spn 6A strain fell to only 38.5% at 1 dpi. By 10 dpi, the clonal diversity of the Spn 6A strain had declined to 3.28% of the inoculum, comparable to the clonal loss that Spn 23F exhibits within the first day following inoculation (Fig. 1A and B). The Spn 23F strain underwent a further loss of diversity over time, with its diversity declining to 1.67% at 10 dpi (Fig. 1B). Despite the decrease in diversity, there was no difference in the overall colonization levels of Spn 23F and 6A strains at both 1 and 10 dpi (Fig. 1D and E). These results suggest that while both Spn 23F and 6A strains undergo a loss of diversity during colonization, there are distinct differences in the dynamics with which they lose this clonal population, with Spn 23F showing a more rapid loss of diversity.
Genetic heterogeneity of the blp locus among Spn isolates impacts their population dynamics in vivo. Graphs (A and B) depict fold change in Hill’s N1 clonal diversity (relative to inoculum) at (A) 1 dpi and (B) 10 dpi, (C) proportion of the most abundant clone at 1 dpi, and (D and E) colonization levels of Spn 6A and 23F strains in the URT at (D) 1 dpi and (E) 10 dpi. Inoculum: 1 × 105 CFU. Each dot represents an individual animal (mean ± SD). P values were calculated by one-way analysis of variance followed by Sidak’s multiple comparisons. ns, statistically not significant.
Variability in the blp locus is responsible for the strain-dependent differences in the loss of clonal diversity
Next, we investigated whether the variability in the blp loci between the Spn 23F and 6A strains could explain the differential dynamics in the loss of diversity between them. To address this question, we constructed blp locus-switch mutants, i.e., the Spn 6A strain expressing the blp locus from the 23F strain (6A Δblp::23F blp) and the Spn 23F strain expressing the blp locus from the 6A strain (23F Δblp::6A blp). To construct these mutants, the native blp locus from each strain was deleted and replaced with the blp locus from the other strain. The resulting strains, 6A Δblp::23F blp and 23F Δblp::6A blp, were molecularly barcoded and contained 2,624 and 2,670 uniquely barcoded clones, respectively (Fig. S1A and B).
We then studied the population dynamics of these blp locus-switch mutants during colonization. At 1 dpi, the 6A Δblp::23F blp strain showed a more rapid loss of diversity than the parental Spn 6A strain, with its clonal diversity falling to 11.1% compared to 38.5% for 6A wild type (WT) (Fig. 1A). Conversely, the 23F Δblp::6A blp strain showed a slower loss of diversity than the parental Spn 23F strain. The clonal diversity for 23F Δblp::6A blp had declined to 19.7% relative to 2.67% for 23F WT within the first day post-inoculation (Fig. 1A). There was no difference in the overall colonization levels of the blp locus-switch mutants compared to their respective parental WT strains (Fig. 1D). These findings suggest that the blp locus from the 23F strain is largely responsible for the more rapid loss of diversity.
Our previous work had shown that activation of BlpC-mediated signaling confers a competitive advantage that allows for a clonal lineage’s expansion at the expense of its kin (14). In accordance, a high abundance of a single clonal lineage corresponds to a bacterial population undergoing a rapid loss of diversity. To test this, we investigated the proportion of the most abundant clones in the populations recovered at 1 dpi. The mean richness of the most abundant clone in the 6A Δblp::23F blp population was higher at 21.5% compared to 2.2% in the 6A WT population (Fig. 1C). This result corresponded to a more rapid loss of diversity observed in the 6A Δblp::23F blp population relative to its parental WT. Additionally, we also observed a decrease in the proportion of the most abundant clone in the population of the 23F genetic background upon switching of the blp locus. The mean richness of the most abundant clone in the 23F Δblp::6A blp population fell to 7% from 41.6% observed in the 23F WT population (Fig. 1C). Together, these results suggest that genetic variability in the blp locus is responsible for differences in the rate of loss of diversity between Spn 23F and 6A isolates.
Functionality of the BlpAB transporter is critical in influencing the dynamics of loss of clonal diversity
Among other differences, one key distinction between the blp loci of the Spn 23F and 6A strains lies in the functionality of the BlpAB transporter. While Spn 6A possesses an intact BlpAB transporter [BlpAB(+) strain], Spn 23F possesses a 4 bp duplication in blpA leading to a frameshift, thus rendering BlpAB non-functional [BlpAB(−) strain]. While replacing the entire blp locus in the 23F strain from 6A WT (23F Δblp::6A blp) turns it into a BlpAB(+) strain, it also introduces other variations in the locus (Fig. 2A). To test whether the loss of an intact BlpAB transporter alone could explain differential dynamics in the loss of diversity, we made targeted mutations in the blpA gene. We deleted the 4 bp duplication in blpA from Spn 23F WT, turning the strain into a BlpAB(+) strain [23F_BlpAB(+)] (Fig. 2A). The 23F_BlpAB(+) strain was then molecularly barcoded to construct a library that contained 3,125 uniquely barcoded clones to enable the study of its population dynamics (Fig. S1C). Following inoculation, the clonal diversity of the 23F_BlpAB(+)_ population had fallen to 18.3% at 1 dpi, showing a slower loss than the parental 23 WT, a BlpAB(−) strain, whose diversity had fallen to 2.67% by this time (Fig. 2B). Furthermore, there was no difference in the rate of loss of diversity between the 23F Δblp::6A blp and 23F_BlpAB(+)_ strains (Fig. 2B). Colonization levels among the three strains remained unchanged (Fig. S2). This suggests that restoring the functionality of the BlpAB transporter is sufficient to result in a slower loss of diversity in the Spn population. Additionally, the difference in the expression of an intact BlpAB transporter between Spn 23F and 6A strains was sufficient to lead to differential dynamics in the loss of Spn diversity in the URT.
Loss of functionality in BlpAB leads to rapid loss of clonal diversity. (A) Gene map depicts the heterogeneity in blp loci from Spn strains 23F WT, 23F Δblp::6A blp, and 23FBlpAB(+). A denotes a 4 bp duplication in blpA, resulting in its premature termination. Purple denotes regulatory genes; yellow denotes BlpAB transporter; blue indicates putative bacteriocins; orange indicates putative immunity proteins; and gray denotes hypothetical proteins. Map not to scale. (B) Graph depicts fold change in Hill’s N1 clonal diversity (relative to inoculum) of Spn strains in the URT at 1 dpi. Inoculum: 1 × 105 CFU. Each dot represents an individual animal (mean ± SD). P values were calculated by one-way analysis of variance followed by Sidak’s multiple comparisons. ns, statistically not significant.*
ComAB is necessary for rapid loss of clonal diversity in BlpAB(−) strains
Spn strains can utilize the paralogous ComAB transporter to secrete blp pheromones and bacteriocins (20, 22). We thus hypothesized that in BlpAB(−) strains with a non-functional BlpAB transporter, ComAB is required to cause the rapid loss of clonal diversity observed in these strains. To test this hypothesis, we deleted ComAB from 23F WT, a BlpAB(−) strain, and 23F_BlpAB(+), i.e., 23F ΔcomAB and 23F_BlpAB(+) ΔcomAB, respectively. Constructing these mutations in the same genomic background allowed us to evaluate the contribution of ComAB to the phenotype in both BlpAB(−) and BlpAB(+) backgrounds. The ΔcomAB strains possessed molecular barcodes to test their population dynamics in vivo. The 23F ΔcomAB and 23F_BlpAB(+)_ ΔcomAB libraries contained 2,953 and 3,017 uniquely barcoded clones, respectively (Fig. S3).
It is worth noting that unlike the BlpAB transporter, the ComAB transporter is intact in almost all Spn strains (22). As such, both Spn 23F and 6A strains possess a functional ComAB transporter (Fig. 3A). In the mouse model, deletion of comAB did not impact colonization levels at 1 dpi in either the BlpAB(−) or BlpAB(+) background (Fig. S4). However, deletion of comAB was sufficient to cause a slower loss of diversity in the BlpAB(−) background. The clonal diversity in 23F ΔcomAB declined to 17.6% at 1 dpi compared to 2.67% for 23F WT (Fig. 3B). This suggested that ComAB is necessary to cause the rapid loss of clonal diversity observed in BlpAB(−) strains. In the BlpAB(+) strain, 23F_BlpAB(+), comAB deletion did not impact population dynamics. While the diversity in the 23F_BlpAB(+) ΔcomAB strain had fallen to 14.4% at 1 dpi, the corresponding value in the 23F_BlpAB(+)_ strain was similar at 18.3% (Fig. 3B). This indicates that in the presence of intact BlpAB, blp secretion through ComAB does not augment changes in the Spn population structure.
Rapid loss of diversity in BlpAB(−) strains is dependent on ComAB. (A) Gene map depicts that comAB loci across Spn 6A and 23F isolates are conserved. Map not to scale. (B) Contribution of comAB in impacting Spn population dynamics. Graph depicts fold change in Hill’s N1 clonal diversity (relative to inoculum) of ΔcomAB Spn strains [23F ΔcomAB and 23FBlpAB(+) ΔcomAB] in the URT at 1 dpi. Data from mice infected with Spn 23F WT and 23FBlpAB(+) strains are also included for comparison. Inoculum: 1 × 105 CFU. Each dot represents an individual animal (mean ± SD). P values were calculated by one-way analysis of variance followed by Sidak’s multiple comparisons. ns, statistically not significant.
Loss of function in blpA is associated with significantly shorter carriage duration
To assess the impact of variation in blpA on competition and carriage characteristics in the natural population, we performed an association study with 2,105 diverse Spn isolates on all variants in the blpA region regardless of their expected functional consequence (Fig. 4A and B). These isolates were obtained from a 2014 longitudinal pneumococcal carriage study of a refugee camp, Maela, the population of which had not received anti-pneumococcal vaccines (27). The study contained 3,085 nasopharyngeal isolates collected from about 1,000 infants and approximately a quarter of their mothers. Because co-carriage was common and carriage rates were high, this population was ideal for studying the impact of BlpAB heterogeneity on Spn colonization dynamics (11). Loss of function (LOF) is common in blpA and occurs through different individual mutations (Fig. 4A). This effect was specific to blpA since variants in the blpA locus display higher significance compared to a housekeeping gene, rpsB (Fig. S5). Furthermore, we also performed a burden test using the presence of any putative LOF variant to determine the strength and direction of association between LOF in the blpA region and host carriage duration (Fig. 4C). We hypothesized that competitive interactions in vivo determine a strain’s success during colonization, thereby impacting its ecological persistence.
Loss of function in blpA is associated with shorter host carriage duration. (A) RelATive cEntrality (RATE) values, a measure of ranked significance, obtained from an association study of all 329 variants identified in the blpA gene, ordered by position along the genome and colored by predicted functional consequence. Higher RATE values indicate greater significance. (B) Credible intervals (CIs) of effect sizes (EFs) of the 16 putative loss-of-function (LOF) variants in the association study. The inner CI represents a probability of 0.5; the outer CI represents a probability of 0.95; and the black line marks the median. Variants with a 0.95 probability CI that does not overlap with an effect size of 0 are considered significant and are marked with an asterisk. A diamond indicates a minor allele frequency of ≥0.05. Direction of association is consistent with the effect size sign (±), and an EF = 1 corresponds to roughly 253 days. (C) Burden test displaying the effect of the presence of at least one LOF mutation in the blpA gene on host carriage duration, which is significantly reduced (median EF = −0.7).
LOF in blpA was significantly associated with shorter host carriage duration, with the median estimate of effect size (−0.7) corresponding to a decrease of roughly 177 days (Fig. 4C). The effect of individual variants was less distinct, with no one LOF variant observed to have a considerably larger effect size than others. However, one LOF variant (A→AGCTT at chromosomal position 562488) was observed at substantially higher frequency than others and is comparable to the 4 bp duplication previously identified as the putative causal variant in lab strains (Fig. 4B). Importantly, suppression of confounding population structure effects dampens the signal from variants not present across multiple strains or present only in strains associated with high or low carriage duration, which is likely for low-frequency and strain-specific variants. The significantly longer host carriage duration observed in isolates with an intact blpA suggests that it may be advantageous during colonization.
Several LOF variants in blpA appear to have been gained independently across separate lineages, indicating potential degenerative evolutionary pressure on the blpAB locus (Fig. S6). However, the ratio of non-synonymous to synonymous substitution rates (dN/dS), and the average number of coding changes between a pair of sequences from different isolates, πaa, indicated that blpA (dN/dS = 0.18, πaa = 0.076) and blpB (dN/dS = 0.12, πaa = 0.023) were both moderately conserved in line with previous estimates of pneumococcal core genes (28). Because the presence of a premature stop codon results in gaps for all downstream residues, which are not considered non-synonymous in these metrics, dN/dS and πaa in this case are more representative of the evolutionary pressures present on functional genes than those that have been truncated by a LOF mutation.
Presence of strains with an intact blpAB combined with the high frequency of non-functional blpAB loci indicates stable maintenance of both these forms of blpAB in the population. Together, these results are consistent with past findings describing negative frequency-dependent selection in bacteriocin production (14, 15, 19).
BlpAB(+) strains have a fitness advantage over BlpAB(−) strains during co-colonization
Having observed that possessing an intact BlpAB results in prolonged carriage in a natural population, we tested whether this also impacts the experimental colonization success of Spn. To this end, we performed competition experiments wherein both BlpAB(−) and BlpAB(+) versions of the 23F strain were co-inoculated in the same mouse in a colonization model. At 1 dpi, neither of the strains had a competitive advantage over one another (Fig. 5A). However, at 7 dpi, the 23F_BlpAB(+)_ strain outcompeted the 23F WT, a BlpAB(−) strain (Fig. 5A). We also validated these results by competing the 23F_BlpAB(+)_ strain with a corrected mutant that expresses the native blpAB with 4 bp duplication in blpA (23F ΔblpAB::blpAB) at 7 dpi. In this case as well, the 23F_BlpAB(+)_ strain had a competitive advantage over the 23F ΔblpAB::blpAB strain (Fig. 5B). It is worth noting that there was no difference in the inherent colonization capacity of each of these strains at 7 dpi when inoculated individually (Fig. 5C). These results indicate that during co-colonization episodes, expressing a functional BlpAB provides Spn with a fitness advantage over BlpAB(−) strains that are exclusively dependent on ComAB for their blp cargo secretion. Thus, despite exhibiting reduced intrastrain competition and slower loss of clonal diversity, BlpAB(+) strains have a distinct advantage during co-colonization that could explain why these strains are maintained in the host population.
BlpAB(+) strains have a competitive advantage over BlpAB(−) Spn during colonization. Graphs depict the competitive index between (A) 23FBlpAB(+) vs 23F WT strains at 1 and 7 dpi, and (B) 23FBlpAB(+) vs 23F ΔblpAB::blpAB strains at 7 dpi. Competitive index greater than 1 indicates an advantage for the 23FBlpAB(+) strain. Each dot represents an individual animal, with the horizontal bar representing the median value. The P value for each condition was calculated by one-sample t and Wilcoxon tests compared to a hypothetical value of 1. Additionally, the competitive index at 1 and 7 days was also compared by unpaired t-test. (C) Graph depicts colonization levels of Spn strains at 7 dpi. Each dot represents an individual animal (mean ± SD). Inoculum: 1 × 105 CFU. P values were calculated by one-way analysis of variance followed by Tukey’s multiple comparisons. ns, statistically not significant.
DISCUSSION
The diversification of Spn into BlpAB(+) and BlpAB(−) types, both of which are maintained within the host population, is an intriguing phenomenon that raises questions regarding the benefits and costs associated with this system. In this work, we demonstrate that Spn strains exhibit distinct behaviors in vivo based on whether they express an intact BlpAB transporter. While BlpAB(−) strains undergo a rapid loss of clonal diversity and extensive intrastrain competition, the BlpAB(+) strains experience a more gradual loss of diversity (Fig. 6). These differences in intrastrain population dynamics translate into a competitive advantage for the BlpAB(+) strain over its BlpAB(−) counterpart during co-colonization. This correlates with increased carriage duration of BlpAB(+) strains in a human population, highlighting the benefit associated with expressing an intact BlpAB transporter.
Proposed model. (A) Table demonstrating that BlpAB(+) strains possessing both ComAB and BlpAB transporters have a lower cell density threshold to activate their quorum, thereby activating bacteriocins and cognate immunity proteins early. As such, these strains exhibit a slower loss of diversity. (B) Schematic demonstrates the kinetics of quorum activation and bacteriocin-mediated killing in both BlpAB(+) and BlpAB(−) strains. In BlpAB(+) strains, owing to their lower cell density threshold for quorum activation, multiple sub-populations can attain the requisite quorum, leading to bacteriocin and immunity protein production. This results in a slower loss of diversity since bacteriocins are not able to kill cognate immunity-expressing Spn cells. In contrast, in BlpAB(−) cells, higher cell density is needed for quorum activation. As such, a sub-population that is first to reach that quorum threshold produces bacteriocins and kills other sub-populations that have yet to have a chance to activate their immunity proteins.
During colonization, Spn cells do not exist as a single uniform population but rather as discrete sub-populations that localize into spatially resolved pockets within the URT (11, 29). As such, activation of quorum sensing in one sub-population gives rise to transcriptomic heterogeneity within the entire colonizing population (14). This will result in a situation where the sub-population that is first to activate quorum sensing produces bacteriocins and the associated immunity proteins (14, 30). This strategy of co-expression of bacteriocins and immunity proteins provides protection from bactericidal suicide. However, other sub-populations may not have yet had an opportunity to activate quorum sensing and, as such, expression of their immunity proteins. These differences in immunity protein expression among colonizing Spn sub-populations present opportunities for bacteriocin-mediated killing of otherwise isogenic kin. An important step in the activation of the Blp quorum sensing pathway is the secretion of the BlpC quorum sensing pheromone through ABC transporters such as BlpAB (16, 21, 31). Our work demonstrates that Spn strains exhibit differential rates of intrastrain competition and colonization success based on whether they express an intact BlpAB transporter or not.
In addition to BlpAB, Spn can utilize the paralogous competence (com) system transporter, ComAB, for secreting peptides of the blp locus. Both ComAB and BlpAB are promiscuous and share the same substrate pool encompassing competence-stimulating peptide (CSP), BlpC pheromone, and blp bacteriocins (20, 22). Signaling via CSP, which drives induction of the com system, kickstarts a regulatory cascade that also results in blp activation (32, 33). It is interesting that in BlpAB(+) strains, blp activation can also only occur independently of com activation (22). In these strains, the presence of BlpAB augments CSP secretion to increase com activation. Thus, com (and consequently blp) activation can occur at lower cell densities in BlpAB(+) strains. In the URT, this results in a situation wherein many sub-populations of BlpAB(+) Spn may be able to independently activate their blp systems and thereby express immunity proteins. The early production of cognate immunity proteins will protect the cells from blp bactericidal activity and prevent a rapid loss of diversity (Fig. 6A and B). As such, we observe a more gradual loss of diversity in the BlpAB(+) population.
In contrast, the BlpAB(−) strains are exclusively dependent on ComAB for blp activation and bacteriocin secretion. Spn exhibits within-strain stochasticity in the pattern of com activation. While ComAB is upregulated during com activation, it is also produced at basal levels within the cell (22). The induction of ComAB expression during competence is transient and restricts bacteriocin secretion to short bursts following com-dependent blp activation. This temporal pattern and higher cell density threshold for com activation further promote transcriptional heterogeneity in the BlpAB(−) population in the URT. In this scenario, many sub-populations will not have reached quorum and activated their blp immunity proteins. This will result in bacteriocin-mediated killing and a rapid loss of diversity (Fig. 6A and B).
Population data from the natural host demonstrate evidence of balanced polymorphism wherein blpA, both with and without the 4 bp duplication that renders BlpAB non-functional, is maintained within the Spn population. However, if ComAB and BlpAB are functionally redundant, why do a small minority of strains (<25%) still possess an intact BlpAB? Our data suggest that during co-colonization, encoding for an intact BlpAB [or BlpAB(+) type] provides a moderate competitive advantage over BlpAB(−) Spn. While the BlpAB(−) type undergoes rapid loss of diversity, the niche vacated by the killed clones may be occupied by clones of the BlpAB(+) type. The delayed induction of immunity proteins in the BlpAB(−) population results in a fitness cost in the context of early bacteriocin production by the BlpAB(+) Spn. In support, previous work has shown that in a BlpAB(−) population, the blp locus has a cost that results in a BlpAB(−) Spn being outcompeted by an isogenic Δblp strain in vivo (14). However, possessing an intact BlpAB provides the bacteria with a competitive advantage over its Δblp counterpart in the URT (22). In the human data, these findings translate into prolonged colonization duration for BlpAB(+) strains. In high co-carriage settings where colonization by multiple strains is frequent (10, 11), this fitness advantage of the BlpAB(+) type results in these strains being maintained in the host population. However, in low co-carriage settings such as in a vaccinated population where co-carriage will be less likely, it is possible that this advantage may no longer outweigh the metabolic cost of expressing BlpAB. This is especially possible, given the functional redundancy of BlpAB and ComAB. It would be interesting to test if the competitive advantage of BlpAB(+) strains is diminished in low co-carriage settings. It also remains to be seen whether the loss of functional BlpAB provides BlpAB(−) strains with an advantage during host-to-host transmission. Regardless, the early domination of a single clonal lineage in this case increases the likelihood of the clonal lineage’s success within the host population. These possibilities could explain why the BlpAB(−) type predominates the Spn population.
The blp locus exhibits extensive genetic variability, especially with regard to the presence or absence of genes and in allelic diversity (17). Despite this, each strain possesses an average of around four bacteriocins. There is tremendous diversity in the bacteriocin content of each strain, with each strain producing cognate immunity proteins specific to the arsenal they encode. In our work, we show BlpAB(+) Spn has an advantage over the BlpAB(−) type during interstrain competition in otherwise isogenic strains. The diversity of bacteriocins does not preclude co-colonization in the human population, as Spn strains with distinct bacteriocin content are routinely found to co-colonize (34). Rather, our findings suggest that the kinetics of the blp locus activation and within-strain population dynamics of each strain may cumulatively determine long-term colonization success during co-colonization.
While this study focused on the population dynamics of Spn strains during nasopharyngeal colonization, our previous work has demonstrated that Spn also undergoes rapid loss of diversity during lung infection (23). This loss of diversity in the lungs facilitates the release of pro-inflammatory factors, including pneumolysin, that drive the development of pneumonia and septicemia (23). It remains to be studied as to how an intact BlpAB transporter influences the dynamics of bacterial turnover in the lung. From an evolutionary perspective, however, invasive disease is an ecological dead end in the life cycle of the bacteria (5). As such, Spn factors primarily adapt to selective pressures during colonization and transmission. In line with this, no difference has been observed in the distribution of BlpAB(+) strains between invasive and colonizing Spn populations (35).
Another interesting aspect of the blp system is its co-regulation with the com system. The reasons why Spn has evolved crosstalk between these two systems have evoked much interest in the field. It has been proposed that blp-mediated bactericidal killing facilitates the release of extracellular DNA that can be utilized by Spn during competence (20). Importantly, our work suggests that transcriptional heterogeneity arising in colonizing Spn sub-populations in vivo will create opportunities where blp-mediated release of genetic material can be accessed during com activation by remaining sub-populations. This may hold yet greater significance in BlpAB(+) strains, wherein the blp system can also turn on before the com system. This access to genetic resources may enable Spn adaptation to the selective pressures of the host environment.
The similarities between the blp and com loci point toward the diversification of their functions following a gene duplication event. However, questions regarding the evolution of these systems and their ancestral state remain. Given this homology between the two systems, it is tempting to speculate that Spn strains possessed an intact BlpAB transporter ancestrally, and this transporter has been lost over time. These BlpAB(−) strains are not “cheaters” in the traditional sense, since they can utilize the ComAB transporter for secreting their cargo. Nevertheless, they are likely more metabolically efficient and increase the chances of clonal dominance during Spn transmission, highlighting the selfish nature of the blp locus.
Finally, our data demonstrate that in high co-carriage settings where competition is likely most consequential, Spn strains with an intact BlpAB colonize humans for a median of 177 days longer relative to strains with a LOF mutation in the transporter. These effects are conserved in strains belonging to many different lineages, indicating that this may not solely be attributed to lineage effects. In support, our in vivo studies of targeted mutations show that otherwise isogenic BlpAB(+) strains have a competitive advantage over their BlpAB(−) counterparts during co-colonization. These results, demonstrating longer carriage duration in the natural population and a competitive advantage during co-colonization, explain why Spn strains with a functional BlpAB continue to persist in the population.
Thus, our work investigating in vivo population structures of Spn provides insight into how genetic variability of the blp locus shapes Spn population dynamics. It also reveals the importance of changes in population dynamics, mediated by bacterial competition, in impacting Spn success during colonization.
MATERIALS AND METHODS
Bacterial strains and growth conditions
Spn 23F (P2499) and 6A (P1476) isolates were used for animal studies in this work. P2499 is a streptomycin-resistant derivative of a 23F clinical isolate which has previously been used for in vivo studies (14, 26, 36). P1476 is also a streptomycin-resistant strain derived from a 6A clinical isolate that has previously been used for in vivo studies (25). All bacterial strains used in this experimental work are listed in Table S1. Colonies were grown from frozen stocks by streaking on tryptic soy agar (TSA)-II agar plates supplemented with 5% sheep blood (BD BBL, New Jersey, USA). Unless otherwise stated, starter cultures were prepared by inoculating streaked colonies in tryptic soy (TS) broth statically at 37°C until they reached an optical density at 620 nm (OD_620_) of 1.0. The cells were then pelleted, washed, and resuspended in sterile phosphate-buffered saline (PBS) for mouse inoculations. Bacterial numbers were enumerated by plating serial dilutions on TSA plates supplemented with 100 µL of catalase (38,000 U/mL; Worthington Biochemical Corporation, New Jersey, USA) and the desired antibiotic (250 µg/mL kanamycin, 200 µg/mL streptomycin, or 200 µg/mL spectinomycin) and incubated overnight at 37°C with 5% CO_2_.
Construction of mutants
Spn mutants were constructed as previously described (14, 37–40). Briefly, colonies were picked and inoculated in acidic Columbia broth (pH 6.6) and grown until an OD_595_ of 0.05, followed by the addition of 5 µg/mL of a 1:1 mix of CSP1 and CSP2, along with 500 ng of transforming DNA. Cultures were incubated statically at 37°C with 5% CO_2_ followed by plating on TSA plates supplemented with 100 µL of catalase (38,000 U/mL, Worthington Biochemical Corporation) and the desired antibiotic (250 µg/mL kanamycin, 200 µg/mL streptomycin, or 10 µg/mL novobiocin). The blp locus-switch mutants were constructed in two steps. First, the native blp locus was deleted from the strains. The blp locus contained a number of hypothetical peptides in addition to bacteriocins, immunity proteins, regulatory genes, and ABC transporters (23). The entire blp locus spanning from blpT to sp_0547 was deleted in these mutants. For this, the blp locus was replaced with a Janus cassette (containing ~1 kb in flanking regions both upstream and downstream of the region of interest) in P2499 and P1476 to obtain 23F blp::Janus (P2700) and 6A blp::Janus (P2720) constructs. These strains were kanamycin resistant. To construct blp locus-switch mutants, P2700 (and P2720) was transformed with genomic DNA from P1476 (and P2499) to obtain P2718 (and P2723). The resulting strains, P2718 and P2723, were back-crossed twice to obtain the blp locus-switch mutants, P2748 (23F Δblp::6A blp) and P2745 (6A Δblp::23F blp). These strains were streptomycin resistant but kanamycin sensitive. To create the 23F_BlpAB(+)_ strain, we first deleted the blpABC operon in P2499 by replacing it with a Janus cassette to obtain the kanamycin-resistant 23F blpABC::Janus (P2762). Targeted mutation PCR was used to construct a PCR fragment containing a targeted deletion of the 4 bp (AAGC) duplication in blpA. P2762 was then transformed with this PCR fragment to obtain the streptomycin-resistant 23F_BlpAB(+)_ strain, P2766. The strain was verified by Sanger sequencing. The 23F ΔblpAB::blpAB strain (P2949) was created by transforming P2762 with the PCR fragment containing the native blpAB loci from 23FWT. To construct ΔcomAB strains, P2499 and P2766 barcoded libraries were transformed with a PCR fragment replacing comAB with a Janus cassette. The resulting strains were kanamycin resistant. To construct a novobiocin-resistant version of 23F WT, P2499 was transformed with a PCR fragment containing a point mutation in gyrB (41). The resulting strain, P2923, was novobiocin resistant. Mutants were confirmed by PCR following each step. All the primers used in this work are listed in Table S2.
Construction of barcoded library
The barcoded library was designed as previously described (14). Briefly, 7 nt barcodes of the sequence NNMCAATGNNMCAAN with intervening fixed sequences were designed to avoid the presence of start and stop codons. The pooled plasmid library previously obtained from Escherichia coli was transformed into Spn strains as described above to obtain molecularly barcoded libraries. The barcoded Spn transformants were selected on TS plates supplemented with 100 µL of catalase (38,000 U/mL; Worthington Biochemical Corporation) and 200 µg/mL of spectinomycin. The resulting barcoded Spn library was grown, sequenced, and stocked at −80°C.
Library sequencing
Genomic DNA from the samples was isolated using MasterPure Complete DNA & RNA Purification Kit (Lucigen, Middleton, WI, USA) as per manufacturer instructions. Barcodes were amplified from genomic DNA using nested PCR, wherein the first step consisted of amplifying the iga region (five cycles) followed by amplification of the barcodes (35 cycles). Primers used for amplification of the barcodes contained the adapters to be used for sequencing library preparation. These amplicons were then purified using the QIAquick PCR purification kit (Qiagen, Germantown, MD, USA) as per manufacturer instructions. Purified samples were then shipped to Azenta Life Sciences (South Plainfield, New Jersey, USA) for sequencing using their Next-Gen Amplicon-EZ service.
Analysis of barcoded sequencing data
The data were analyzed as previously described (14). Reads were aligned to a reference sequence using Python. First, Trimmomatic was used for quality control to trim adapter sequences and low-quality bases from the reads (sliding window size: 3, sliding window quality: 20, leading and trailing quality: 15, minimum length: 75) (42). The reads were then aligned to a reference sequence by BWA (matching score: 10, mismatch penalty: 2) and outputted in a .sam file (43). The remainder of the analysis was done using R. The barcode sequence was extracted from the aligned reads by concatenating bases at known variable positions while filtering out incomplete or ambiguous barcodes. A table detailing each barcode detected and the number of times it was found was compiled. To account for variability in the number of total reads, we standardized samples by computing rarefaction and extrapolation of clonal diversity using iNEXT (44, 45). The clonal diversity was expressed using Hill numbers with q = 1 (Hill’s N1), which is the exponential of Shannon index (45). Shannon diversity index (H) was calculated as , where p_i_ denotes the proportion of the population made up of the clone i. The Shannon index estimates the uncertainty associated with predicting the identity of a species randomly chosen from a community.
Animal studies
Wild-type C57BL/6J (strain 00664) mice were purchased from The Jackson Laboratory (Bar Harbor, ME, USA). The mouse colony was bred and maintained in a conventional mouse facility. Infant pups were housed with the dam until weaning at 3 weeks of age. Adult mice were fed ad lib the PicoLab Rodent Diet 20, a 20% protein diet formulation, and were given water for consumption. All the animals were kept on a light cycle of 12 hours on, 12 hours off with a temperature in the animal facility of 70°F (±2°F).
Infant colonization model
Three- to four-day-old infant pups were inoculated intranasally with 10^5^ CFU of Spn in 3 µL of sterile PBS with a pipette tip, without anesthesia. The pups were returned to their dam for the duration of the experiment. At the end of the experiments, mice were euthanized at the indicated time point by CO_2_ asphyxiation followed by cardiac puncture. The Spn colonization density of the upper respiratory tract was measured as previously described (46). Briefly, the trachea was lavaged using a 30 gauge needle for infants with 300 µL of sterile PBS collected from the nares. Forty microliters of this retrotracheal lavage was used to enumerate bacteria by viable plating serial dilutions on TSA-catalase plates supplemented with the appropriate antibiotic (200 µg/mL streptomycin) and incubated overnight at 37°C with 5% CO_2_. The remaining lavage was grown in TS broth supplemented with 200 µg/mL spectinomycin at 37°C until it reached an OD_620_ of 1.0 for genomic DNA isolation.
Population genomic analyses
Whole-genome sequences and metadata were obtained from a 2014 longitudinal pneumococcal carriage study of a refugee camp, Maela, the population of which had not received anti-pneumococcal vaccines (27). Genotyping was performed using Snippy (v.4.6.0) for assembly and variant calling with a serotype 6B S. pneumoniae reference genome (National Center for Biotechnology Information [NCBI] Reference Sequence: NC_014498.1, BioSample: SAMN02604004). VCF file indexing, filtering to the blpA, blpB, or rpsB (47) region according to the reference annotation, cleaning, and merging were accomplished with bcftools (v.1.17) and samtools (v.1.19). Variant calling in the population identified nine frameshifts and seven stop gains as putative LOF variants due to their predicted functional consequence (premature truncation of blpA). A total of 313 synonymous and missense variants were found, along with one conservative in-frame deletion, but these variants were not expected to cause significant loss of function.
The association study and burden test were performed using a Bayesian linear mixed model implemented in Stan (v.2.34.0) and run with the variational inference algorithm with 10 k iterations. This regression model employs PopPUNK lineage clusters, here obtained using PopPUNK (v.2.6.5), as a random effect to control for lineage effects (48). cppRATE (v.0.2.0) (https://github.com/tmaklin/cpprate) was used to extract relative centrality (RATE) metrics from the fitted model. Multicollinearity in the form of linkage disequilibrium was observed between some blpA variants; this presented a challenge, as strongly correlated parameters cannot be jointly estimated due to identifiability issues. As two or more variants in perfect linkage disequilibrium (D′ = 1) will have the same fit and significance, all but one representative variant per D′ = 1 block were excluded prior to model fitting and RATE estimation. RATE values of the representative were applied to the excluded variants post-estimation.
To calculate the πaa for each gene, the gene region (reverse complemented to obtain the correct reading frame) was translated using Expasy’s dna2aa programmatic access interface (https://web.expasy.org/cgi-bin/translate/dna2aa.cgi, accessed 30 October 2024). The resulting amino acid sequences were aligned with MUSCLE (v.5.1), and πaa was calculated using DendroPy (v.5.0.1). A nucleotide alignment was then generated from this AA alignment using RevTrans (v.1.4), and a phylogenetic tree was created using IQ-TREE (v.2.2.6). With these as input, relative evolutionary rate inference was run on HyPhy (v.2.5.62) to infer dN/dS values.
A phylogenetic tree was generated for isolates with host carriage duration data using Parsnp (v.2.0.4) and the aforementioned reference genome (NCBI Reference Sequence: NC_014498.1, BioSample: SAMN02604004) (Fig. S6).
Competition experiment
For the competition experiment, the strains were grown in TS broth until they reached an OD_620_ of 1. The cells were collected, washed, and resuspended in sterile PBS. Both strains were then mixed at a ratio of 1:1. Infant pups were then inoculated intranasally as described above with 10^5^ CFU of the mix. At both 1 day and 7 dpi, URT lavages were collected, and the density of each strain was determined by selective plating on different antibiotics (200 µg/mL streptomycin, 200 µg/mL spectinomycin, or 10 µg/mL spectinomycin). A competitive index was calculated by comparing the output CFU ratio of the two strains obtained from the lavages to their input ratio from the inoculum. A competitive index greater than 1 suggests a competitive advantage for the first strain, while a value less than 1 indicates an advantage for the second strain.
Statistical analysis
The details of the statistical analyses are included in the figure legends. The statistical analyses were performed using GraphPad Prism (v.10.1.1; GraphPad Software Inc., San Diego, CA, USA) unless stated otherwise.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Collaborators GAR. 2022. Global mortality associated with 33 bacterial pathogens in 2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet 400:2221–2248. doi: 10.1016/S 0140-6736(22)02185-736423648 10.1016/S 0140-6736(22)02185-7PMC 9763654 · doi ↗ · pubmed ↗
- 2Collaborators GLRI. 2018. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory infections in 195 countries, 1990–2016: a systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect Dis 18:1191–1210. doi: 10.1016/S 1473-3099(18)30310-430243584 10.1016/S 1473-3099(18)30310-4PMC 6202443 · doi ↗ · pubmed ↗
- 3O’Brien KL, Wolfson LJ, Watt JP, Henkle E, Deloria-Knoll M, Mc Call N, Lee E, Mulholland K, Levine OS, Cherian T. 2009. Burden of disease caused by Streptococcus pneumoniae in children younger than 5 years: global estimates. The Lancet 374:893–902. doi:10.1016/S 0140-6736(09)61204-619748398 · doi ↗ · pubmed ↗
- 4Bogaert D, De Groot R, Hermans PWM. 2004. Streptococcus pneumoniae colonisation: the key to pneumococcal disease. Lancet Infect Dis 4:144–154. doi:10.1016/S 1473-3099(04)00938-7doi:14998500 · doi ↗ · pubmed ↗
- 5Weiser JN, Ferreira DM, Paton JC. 2018. Streptococcus pneumoniae: transmission, colonization and invasion. Nat Rev Microbiol 16:355–367. doi:10.1038/s 41579-018-0001-8doi:29599457 PMC 5949087 · doi ↗ · pubmed ↗
- 6De Lencastre H, Tomasz A. 2002. From ecological reservoir to disease: the nasopharynx, day-care centres and drug-resistant clones of Streptococcus pneumoniae. J Antimicrob Chemother 50 Suppl S 2:75–81. doi:10.1093/jac/dkf 511doi:12556436 · doi ↗ · pubmed ↗
- 7Abdullahi O, Karani A, Tigoi CC, Mugo D, Kungu S, Wanjiru E, Jomo J, Musyimi R, Lipsitch M, Scott JAG. 2012. The prevalence and risk factors for pneumococcal colonization of the nasopharynx among children in Kilifi District, Kenya. P Lo S One 7:e 30787. doi:10.1371/journal.pone.0030787 doi:22363489 PMC 3282706 · doi ↗ · pubmed ↗
- 8Lee GM, Kleinman K, Pelton S, Lipsitch M, Huang SS, Lakoma M, Dutta-Linn M, Rett M, Hanage WP, Finkelstein JA. 2017. Immunization, antibiotic use, and pneumococcal colonization over a 15-year period. Pediatrics 140:e 20170001. doi:10.1542/peds.2017-0001 doi:28978716 PMC 5654389 · doi ↗ · pubmed ↗