Maternal transfer of oral vaccine induced anti-OspA antibodies protects Peromyscus spp. from tick-transmitted Borrelia burgdorferi
Jose F. Azevedo, Greg Joyner, Suman Kundu, Kamalika Samanta, Maria Gomes-Solecki

TL;DR
Oral vaccination of mother mice with OspA protects their offspring from tick-transmitted Lyme disease bacteria.
Contribution
Demonstrates maternal transfer of anti-OspA antibodies can block Borrelia burgdorferi infection in offspring.
Findings
Vaccinating dams until parturition and 2 weeks postpartum conferred protection to pups against B. burgdorferi.
Pups showed no detectable B. burgdorferi DNA or viable spirochetes in heart and bladder tissues.
Maternal vaccination timing significantly affects antibody transfer and infection prevention.
Abstract
The efficacy and duration of passive immunity protection depend on maternal antibody levels and transfer efficiency. We investigated whether oral vaccination of Peromyscus leucopus dams with recombinant outer surface protein A (OspA)-expressing Escherichia coli could induce maternal transfer of anti-OspA antibodies and protect pups from Borrelia burgdorferi challenge. Dams were vaccinated until breeding pairs were created (i), until parturition (ii), and until pups were 2 weeks old (iii). Pups were challenged with nymphal Ixodes scapularis-transmitted B. burgdorferi at ~4 weeks of age. Anti-OspA IgG was quantified in dams and pups, and anti-B. burgdorferi IgG was quantified in pups. B. burgdorferi burden was assessed by flaB quantitative PCR in pups’ tissues ~4 weeks after tick challenge, and viability of B. burgdorferi was assessed by culture of the heart tissue. P. leucopus pups born…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
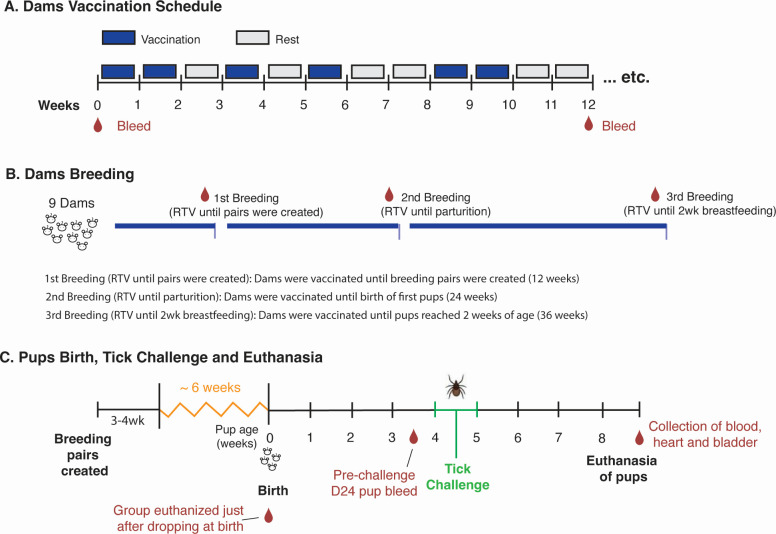 Fig 1
Fig 1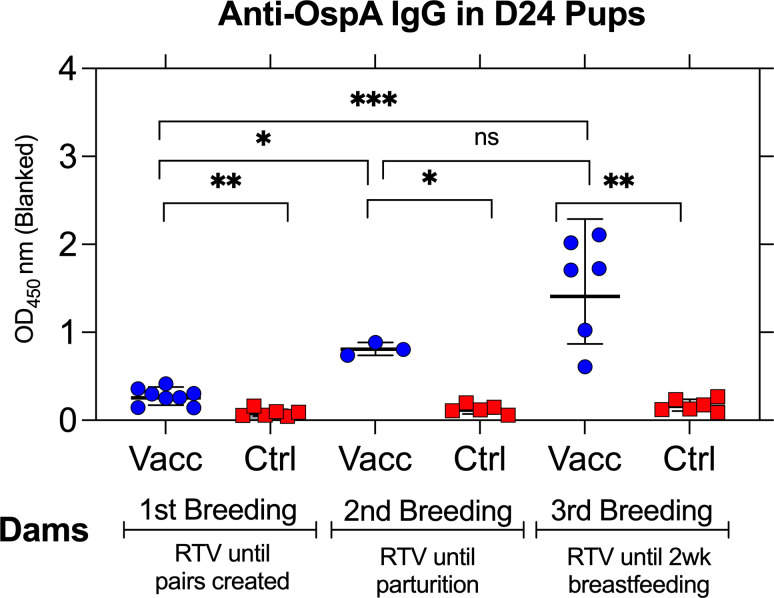 Fig 2
Fig 2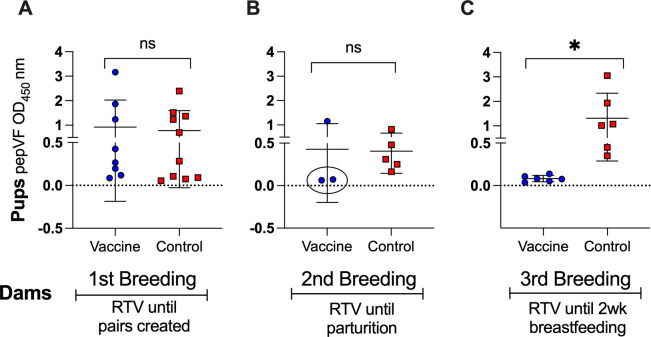 Fig 3
Fig 3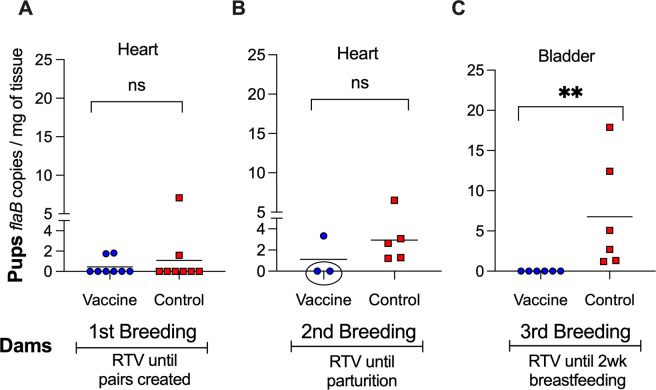 Fig 4
Fig 4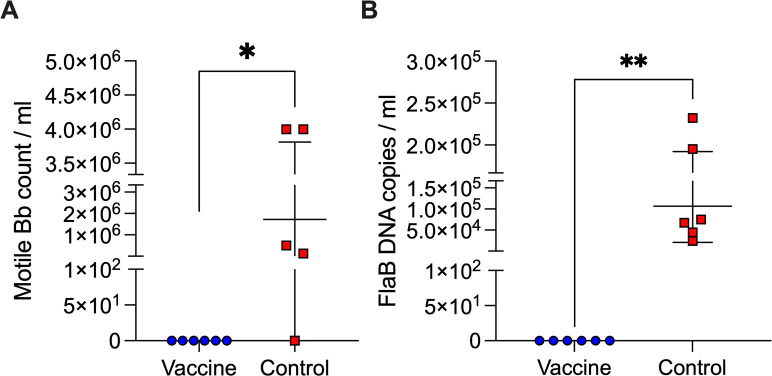 Fig 5
Fig 5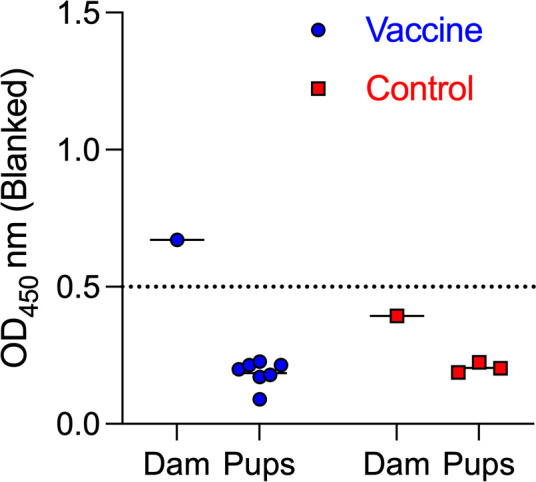 Fig 6
Fig 6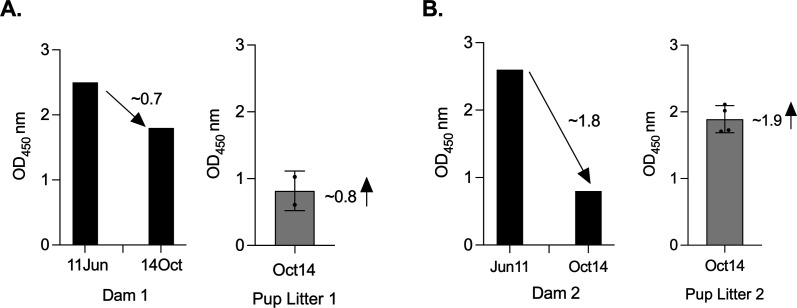 Fig 7
Fig 7- —HHS | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- —HHS | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- —HHS | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- —HHS | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Viral Infections and Vectors · Vector-Borne Animal Diseases
INTRODUCTION
Lyme disease, caused by Borrelia burgdorferi, is the most prevalent vector-borne disease in North America (1). The white-footed mouse (Peromyscus leucopus) serves as the primary reservoir host (2), maintaining B. burgdorferi in enzootic cycles and facilitating transmission to Ixodes ticks that subsequently infect humans and other mammals (3). Unlike humans, reservoir hosts do not develop significant disease following B. burgdorferi infection, yet they serve as a crucial source of spirochete acquisition for larval Ixodid ticks (4). Thus, interrupting the transmission of B. burgdorferi (5) has been proposed as a strategy for controlling Lyme disease (6, 7).
Outer surface protein A (OspA) is a major target for Lyme disease vaccine development (8) due to its essential role in B. burgdorferi persistence within the tick vector (9). OspA facilitates spirochete adherence to the tick midgut, allowing the bacterium to survive between blood meals (10). During tick feeding, B. burgdorferi downregulates OspA while upregulating OspC (11), which enables migration to the salivary glands and subsequent transmission to the host (12–14). Vaccines targeting OspA aim to elicit antibodies that neutralize spirochetes within the tick gut, preventing their transmission before they reach the mammalian host (15, 16). These vaccines have been explored in multiple contexts, including human vaccination (17, 18), direct vaccination of P. leucopus (19), and oral bait-delivered reservoir-targeted vaccines (RTVs) (6).
Beyond direct vaccination, maternal transfer of antibody provides an alternative strategy to protect neonates during the critical early stages of life (20). In the context of the ecology of vector-borne pathogens, this temporary immunity reduces early-life susceptibility to infection (21). Previous studies have demonstrated that oral vaccination of dams with OspA results in anti-OspA IgG in offspring (22), though the degree to which this protects against B. burgdorferi transmission remains unknown. In this study, we addressed this critical gap by investigating the extent of maternal antibody transfer in P. leucopus and assessing whether vaccination of dams with recombinant OspA-expressing Escherichia coli confers protection to neonates. We tested whether vaccination of dams until breeding pairs were created, until parturition, and until the pups reached 2 weeks of age influences antibody transfer and protection against tick-mediated B. burgdorferi challenge. Our findings have implications for reservoir-targeted vaccination strategies aimed at reducing Lyme disease risk by breaking the transmission cycle of B. burgdorferi.
MATERIALS AND METHODS
Vaccine preparation
Recombinant E. coli BL21(DE3)pLysS encoding B. burgdorferi OspA (E. coli-OspA) was produced as previously described (23). Briefly, E. coli-OspA was grown in Terrific Broth (TBY) at 37°C until OD_600_ ~0.8, induced with 1 mM isopropyl β-D-thiogalactopyranoside (IPTG, 97%, Thermo Scientific) for 3 h, transferred to filter-top flasks and inactivated with 1% β-propiolactone (BPL, 98%, AlfaAesar), incubated at room temperature at 100 rpm for 24 h, and harvested by centrifugation at 2,000 × g at 4°C. The bacterial biomass was resuspended in phosphate-buffered saline (PBS) and sprayed onto feed pellets. Pellets (RTV pellets) were allowed to dry between application layers. Inactivation was tested by inoculation of BPL-treated culture in TBY (1:200) and incubation overnight at 37°C, shaking at 250 rpm. The following day, culture inactivation was confirmed by checking turbidity and recording OD_600_ = 0.
Vaccination strategy
Dams (n = 9) received RTV pellets daily for 5 consecutive days per week, with a 2 day rest period over the weekend in which they received regular mouse chow. Control dams received uncoated pellets. Dams consumed RTV pellets continuously as per vaccination schedule up to 36 weeks (Fig. 1) and were assigned to three experimental groups based on breeding milestones. The level of IgG to OspA was analyzed in dams’ blood before selecting animals with high antibody in serum to create breeding pairs.
Experimental design. (A) Oral vaccination schedule of adult female Peromyscus leucopus: mice were allowed to eat RTV pellets on vaccination weeks; pellets were exchanged for regular feed during rest weeks; control mice received uncoated pellets. (B) Timeline for breeding of dams: dams were bled to evaluate serologic antibody to OspA, and three experimental groups were set as first, second, and third breeding based on duration of vaccination and breeding milestones. (C) Birth of pups, challenge with B. burgdorferi-infected I. scapularis ticks, and collection of samples.
Breeding and litter management
Breeding pairs were established based on dams having anti-OspA titers (OD_450_ >0.8) in serum diluted 1:100 assessed by enzyme-linked immunosorbent assay (ELISA). Each vaccinated female was cohoused with an unvaccinated male for 3–4 weeks, after which pregnant females were single-housed until parturition (Fig. 1). The first group (Breeding 1) was created with five dams vaccinated for 12 weeks prior to breeding pairs being formed. The second group, Breeding 2, was created with four dams vaccinated for 24 weeks until parturition. The third group, Breeding 3, was created with the dams who maintained high levels of antibodies to OspA and remained on the vaccination regimen for 36 weeks, ensuring that immunization continued until their pups reached two weeks of age. Dams that lost the ability to reproduce or had low levels of serologic antibody to OspA were euthanized. A group of pups (n = 7) born after the third breeding were euthanized immediately after they dropped at parturition and blood was collected. Pups remained with their dams until 24–25 days of age, at which point they underwent nymphal tick challenge.
Blood collection
Blood samples were collected via submandibular vein bleed. Dams were sampled at day 0 and before each breeding. Pups were sampled on the day they were born, at day 24 (pre-challenge), and day ~71 (terminal bleed). Samples were allowed to clot for 30 min at room temperature before centrifugation at 2,000 × g for 10 min at 4°C. Serum was aliquoted and stored at −80°C until analysis.
Tissue collection and processing
At necropsy on day 71 (~9- to 10-week-old pups), heart and bladder tissues were harvested, transported on dry ice, and stored at −80°C. For organ culture experiments, freshly excised heart tissue was immediately placed into Barbour-Stoenner-Kelly H (BSK-H) medium supplemented with 6% rabbit serum (Sigma-Aldrich, St. Louis, MO) and incubated at 34°C for 3 weeks (on the 7th day of culture, the tissue was removed to avoid putrefaction). On day 21 of culture supernatants were analyzed for motile B. burgdorferi detection using a Petroff-Hausser chamber (dark-field microscopy, 200× magnification), and for DNA quantification via flaB quantitative PCR (qPCR), to assess the presence and viability of the spirochetes in cardiac tissue cultures.
Tick challenge and monitoring of B. burgdorferi infection
To assess susceptibility to B. burgdorferi infection, pups were challenged with infected nymphal I. scapularis ticks at ~4 weeks of age. Each pup was exposed to five to seven infected nymphs, which were allowed to feed for 72 hours in a controlled containment chamber. Flat nymphal ticks were carefully transferred from storage vials using fine forceps and placed at the base of the neck and upper back of gently restrained pups. After placement, pups were monitored to ensure proper tick attachment before being returned to their cages. After 72 hours, all engorged ticks naturally detached and were collected from the bottom of the cages. Detached ticks were stored at −20°C until further analysis for B. burgdorferi colonization via dark-field microscopy.
The infected nymphs used in this study were generated by feeding larval I. scapularis (Oklahoma Tick Laboratory) on a mouse infected with a multi-strain culture of B. burgdorferi sensu stricto. This B. burgdorferi culture was recovered from heart and bladder tissues of C3H/HeN mice infected with nymphal ticks collected in Massachusetts in 2020. To maintain infection across generations, the multi-strain B. burgdorferi inoculum was first introduced into C3H mice, after which successive mice were infected through tick-borne transmission. Larvae were allowed to feed on infected mice, acquire B. burgdorferi, and molt into infected nymphs, which were then used to challenge mice in a cycle that mimics natural enzootic transmission of B. burgdorferi.
Detection of anti-OspA and anti-PepVF IgG by ELISA
Serum antibody responses to OspA and to B. burgdorferi PepVF were assessed using an ELISA. PepVF contains two peptides from B. burgdorferi VlsE and FlaB proteins in tandem, separated by a 3 aa glycine linker, and is used in our lab to determine B. burgdorferi infection (24). Presence of antibody to PepVF indicates exposure to B. burgdorferi. Purified recombinant OspA (1 µg/mL) or recombinant PepVF (1 µg/mL) was adsorbed onto Nunc MaxiSorp plates (Thermo Fisher Scientific, Waltham, MA) by overnight incubation at 4°C. Following antigen coating, the plates were blocked with 5% bovine serum albumin in PBS-Tween (0.05%) for 1 hour at room temperature to prevent nonspecific binding. Serum samples were diluted 1:100 in blocking buffer and incubated on the plates for 2 hours at 37°C. To detect antigen-specific IgG, a goat anti-mouse IgG antibody conjugated to horseradish peroxidase (1:6,000; Jackson ImmunoResearch, West Grove, PA) was added, followed by a 1 hour incubation at 37°C. After washing, plates were developed using 3,3', 5, 5' - tetramethylbenzidine (TMB) substrate (Sigma-Aldrich) for 15 min before stopping the reaction with 1 N H_2_SO_4_. Optical density (OD) was measured at 450 nm and blanked against the control using a SpectraMax Plus ELISA reader (Molecular Devices, San Jose, CA). For cutoff determination, the positivity threshold was defined as mean + 3 SD of negative controls.
Molecular detection of B. burgdorferi DNA by qPCR
Total DNA was extracted from heart and bladder tissues and heart culture media using the DNeasy Blood & Tissue Kit (Qiagen, Germantown, MD) according to the manufacturer’s protocol. qPCR was performed to quantify B. burgdorferi flaB gene copies per milligram of tissue or per milliliter of culture media. qPCR was performed to quantify B. burgdorferi flaB gene copies in DNA extracted from heart and bladder tissues, as well as culture supernatants. Reactions were carried out in a 25 µL volume using 2X TaqMan Master Mix (Thermo Fisher Scientific), with final primer and probe concentrations of 200 nM. Each reaction contained 5 µL of DNA template, and all samples were run in duplicate to ensure reproducibility. The primer and probe set used for TaqMan-based detection of the flaB gene was as follows: forward primer: 5′-AAGCAATCTAGGTCTCAAGC-3′; reverse primer: 5′-GCTTCAGCCTGGCCATAAATAG-3′; probe: 5′-FAM-AGATGTGGTAGACCCGAAGCCGAG-TAMRA-3′. Amplification was performed using a QuantStudio 7 Real-Time PCR System (Applied Biosystems) under the following cycling conditions: an initial denaturation at 95°C for 10 min, followed by 45 cycles of 95°C for 15 seconds and 60°C for 1 min. Fluorescence was measured at each cycle, and flaB copy numbers were determined by comparison to a standard curve generated from known concentrations of B. burgdorferi genomic DNA.
Statistical analysis
Statistical analyses were conducted using GraphPad Prism 9.0 (GraphPad Software, San Diego, CA). To assess the distribution of the data, the Shapiro-Wilk test was applied to determine normality. For group comparisons, an unpaired t-test was used when data followed a normal distribution, while the Mann-Whitney U-test was applied to non-normally distributed data. A significance threshold of P < 0.05 was used for all statistical tests.
RESULTS
Transfer of anti-OspA antibodies from dams to pups
Pups were born to dams undergoing a continuous vaccination schedule. To determine the extent of maternal antibody transfer, serum anti-OspA IgG titers were measured in pups when they reached day 24 (pre-challenge). Pups born from dams vaccinated until breeding pairs were formed are grouped under first breeding. Pups born from dams vaccinated until parturition are grouped under second breeding. Pups born from dams vaccinated until the pups were 2 weeks old are grouped under third breeding. The results are shown in Fig. 2. Anti-OspA IgG was significantly increased in day 24 pups born from vaccinated dams during the first, the second, and the third breeding, in contrast to pups born to control unvaccinated dams. Nevertheless, we note that pups born from the first breeding of vaccinated dams had an average anti-OspA IgG OD_450_ ~0.271 (SD ± 0.094); pups born from the second breeding of vaccinated dams had an average anti-OspA IgG OD_450_ ~0.809 (SD ± 0.073); and pups born from the third breeding of vaccinated dams had an average anti-OspA IgG OD_450_ ~1.532 (SD ± 0.591). The data show that continued vaccination of the dams past parturition maintained sufficient levels of protective antibody (OD_450_ >0.8) in blood.
*Transfer of OspA antibodies from dams to pups. ELISA quantification of pre-challenge anti-OspA IgG in sera from day 24 pups born to vaccinated dams. OD at 450 nm (blanked) is shown for each group. Each point represents an individual pup, with lines indicating group means ± SD. Statistical analysis by Mann-Whitney U-test, *P < 0.05, **P < 0.005, **P < 0.0005.
Seroconversion to B. burgdorferi in tick-challenged pups
To assess whether pups were protected or infected with B. burgdorferi after tick challenge, anti-PepVF IgG was measured in serum samples collected at euthanasia (~day 71), at least 3 weeks after the last day of challenge (Fig. 3). Pups (n = 8) from the first breeding of vaccinated dams had equivalent anti-PepVF IgG as control pups (n = 10), confirming infection with tick-transmitted B. burgdorferi in both groups (P = 0.7642) (Fig. 3A). Although 2/3 pups from the second breeding of vaccinated dams did not have anti-PepVF antibodies (Fig. 3B, circle), overall differences with the control group (n = 5) were not significant (P = 0.9432). Strikingly, pups (n = 6) from the third breeding of vaccinated dams (Fig. 3C) had anti-PepVF IgG levels below the cutoff in contrast to the control group (n = 6), P = 0.0147, indicating absence of infection. These results suggest that maternal vaccination that extended until and beyond parturition conferred anti-OspA passive immunity to pups.
*Seroconversion to B. burgdorferi in tick-challenged pups. (A–C) ELISA quantification of anti-PepVF IgG in terminal sera from pups born to vaccinated dams. Pups were bled ~4 weeks post-tick challenge. OD at 450 nm (blanked) is shown for each group. Each data point represents an individual pup, with lines indicating group means ± SD. Statistics by unpaired t-test: ns, not significant, P < 0.05.
Burden of B. burgdorferi in tissues from tick-challenged pups
To provide additional evidence of B. burgdorferi dissemination, we purified DNA from heart and bladder organs from pups euthanized >4 weeks after tick challenge (~ day 71) and performed qPCR targeting the B. burgdorferi flaB gene (Fig. 4). In pups from the first breeding, there are no differences in flaB loads in heart tissues from vaccinated and control groups; in pups from the second breeding, the same two mice (2/3) in the vaccinated group that did not seroconvert to B. burgdorferi also did not have flaB in heart tissues, whereas 5/5 mice in the control group had flaB DNA in heart tissues. Differences between the groups are not significant (P = 0.3393). However, in pups from the third breeding, no detectable flaB copies were found in bladder tissues from the vaccinated group (0/6), in contrast to the pups from the control group that were all positive (6/6) for B. burgdorferi flaB (P = 0.0022). Absence of B. burgdorferi flaB DNA in pups from the second and third breeding correlates with absence of PepVF seroconversion, further confirming the protective effect of maternal antibody transfer when dams were vaccinated until and beyond parturition.
*Burden of B. burgdorferi in tissues from tick-challenged pups. (A–C) qPCR quantification of the flaB gene in heart and bladder tissues collected from pups ~4 weeks post tick challenge. Each data point represents an individual pup, with lines indicating group means. Statistics by Mann-Whitney U-test: ns, not significant, *P < 0.005.
Viability of B. burgdorferi in tissues from tick-challenged pups
To further assess B. burgdorferi persistence, heart tissues from pups born from third breeding dams were cultured in BSK-H medium at 34°C for 21 days, and live spirochetes were counted using a Petroff-Hausser chamber under a dark-field microscope (Fig. 5A). Only pups (4/5) from unvaccinated control dams had detectable motile B. burgdorferi in culture, confirming that live spirochetes had successfully colonized cardiac tissue after tick challenge. The control group exhibited a range of 10^5^–10^6^ spirochetes per milliliter of culture, while pups from vaccinated dams had no visible live bacteria, P = 0.0152. qPCR analysis of heart culture supernatants (Fig. 5B) confirmed that high flaB copy numbers (mean 10^5^) were detected in control heart cultures (5/5), whereas no B. burgdorferi DNA was present in pups from vaccinated dams (0/5), P = 0.0022. This indicates that dams that received the vaccine until 2 weeks post-parturition produced pups that were protected from cardiac colonization by B. burgdorferi.
*Viability of B. burgdorferi in tissues from tick-challenged pups. Quantification of B. burgdorferi in cultures of the heart tissue from pups at ~4 weeks post-challenge: (A) enumeration of motile B. burgdorferi using a Petroff-Hausser chamber under a dark-field microscope; (B) qPCR quantification of B. burgdorferi flaB in heart culture media. Each data point represents an individual pup, with lines indicating group means ± SD. Statistics by Mann-Whitney U-test, *P < 0.05 and *P < 0.005.
Absence of OspA IgG in serum from pups euthanized after birth
To determine whether maternal antibodies were transferred in utero, OspA ELISA was performed on serum from pups euthanized immediately after they dropped at birth in comparison to the respective dam (Fig. 6). In contrast to the respective dam, no anti-OspA IgG was detected in newborns from the vaccinated dam, confirming that placental transfer of antibodies was absent.
Absence of OspA IgG in serum from pups euthanized after birth. ELISA quantification of anti-OspA IgG in serum from pups euthanized immediately after birth from dams vaccinated until 2 weeks parturition (third breeding). OD at 450 nm (blanked) is shown. Each data point represents an individual pup, with lines indicating group means. The cutoff value (dashed line) represents the mean + 3 SD of negative controls.
Anti-OspA IgG depletion from vaccinated dam serum after lactation
To investigate whether maternal antibody depletion occurred due to antibody transfer to offspring, serum anti-OspA IgG was measured in dams used for the third breeding at two time points (11 June and 14 October) and in the respective litters, on the latter time point, before tick challenge (Fig. 7). We found that the reduction in OspA antibody measured between the two time points in the dams corresponds to the increase in OspA antibody measured in the respective litters in the latter time point: ~0.7OD_450_ for Dam 1 (Fig. 7A) and ~1.8OD_450_ for Dam 2 (Fig. 7B). These findings suggest that the transfer of antibody from dam to pups may be dependent on the efficiency of the pups’ suckling.
Anti-OspA IgG depletion from vaccinated dams serum after pups' suckling. ELISA quantification of anti-OspA IgG in serum from two dams used in the 3rd breeding and the respective litters: (A) Dam 1/Litter 1, (B) Dam 2/Litter 2. Black histograms represent OD450 from dams serum collected at two time-points ~ 4 months apart. Grey bar graphs represent OD450 from the respective litters serum collected on the latter time point, before tick challenge. Each data point represents an individual pup, with lines indicating group means ± SD. Decreasing and increasing arrows depict OD450 differences in dams and litters.
DISCUSSION
Our study demonstrates that vaccination of Peromyscus leucopus dams against B. burgdorferi leads to transfer of anti-OspA IgG to offspring and that the extent of transfer and protective efficacy depends on maintenance of maternal immunization. Pups born to dams vaccinated until parturition and until 2 weeks post-parturition exhibited significantly higher anti-OspA IgG levels than those from dams vaccinated until breeding pairs were created. After challenge with B. burgdorferi-infected I. scapularis ticks, pups born to vaccinated dams that maintained high systemic anti-OspA IgG had no anti-B. burgdorferi antibody in serum, no B. burgdorferi DNA in bladder or heart tissues, and no motile B. burgdorferi in the heart tissue. Furthermore, we detected no serologic anti-OspA IgG in offspring from vaccinated dams, euthanized immediately after birth, before suckling. The data indicate that anti-OspA antibody is transferred from P. leucopus dams to pups via lactation and that it protects against tick-transmitted B. burgdorferi.
Vaccination of P. leucopus dams until the breeding pairs were created (first breeding) did not protect P. leucopus pups from tick-mediated infection due to the absence of sufficient levels of anti-OspA antibodies (OD_450_ <0.8, Fig. 2) in pups born ~10 weeks after the dams were vaccinated. This was surprising given our previous studies in P. leucopus (25) and in C3H/HeN mice (22) in which we vaccinated mice with the RTV. In the first study, we vaccinated P. leucopus with 20–30 units of live RTV for 1–4 months and found that mice maintained anti-OspA antibody for up to a year (25). In the second study, we found that C3H/HeN dams vaccinated with 20 units of live RTV over a period of 8 weeks produced pups 33–79 days after vaccination that maintained protective titers of OspA antibody from 2 to 9 weeks after birth (22). The major difference between the studies is that in the current study, we used RTV inactivated with BPL, whereas in the former studies, we used live RTV. This change was intentional to evaluate a vaccine that mimics the product currently licensed by USDA, which is inactivated with BPL (26). These data indicate that the production of anti-OspA antibody by P. leucopus requires persistent vaccination, if the vaccine formulation is inactivated likely because it does not colonize the mouse gut.
Previous studies show that the transfer of maternal IgG provides passive immunity but does not necessarily confer sterilizing protection against vector-borne pathogens (20, 21). The degree of IgG transfer and its impact on early-life immunity can vary depending on maternal antibody titers, the mechanism of transfer (transplacental vs lactogenic), the duration of exposure (27), and the neutralizing capability of the antibody (25). In our study, the neutralizing capability of anti-OspA antibody transferred from P. leucopus dams to its pups is evidenced by the absence of B. burgdorferi antibody in serum of pups (2/3 in Breeding 2 and 6/6 in Breeding 3, Fig. 3), as well as absence of DNA in the heart and bladder tissues (the same mice 2/3 in Breeding 2, and 6/6 in Breeding 3, Fig. 4) and complete absence of motile spirochetes in cultures from the heart tissue (6/6 in Breeding 3, Fig. 5). All pups from Breeding 1, born to vaccinated and control dams, had antibodies to B. burgdorferi antigens after tick challenge (Fig. 3), but we did not detect B. burgdorferi DNA in the heart tissue of these mice (Fig. 4). We perform two independent tests (antibody to B. burgdorferi in serum and B. burgdorferi DNA in tissues), and we accept one positive result as evidence of B. burgdorferi dissemination (24). This method offsets risks incurred from tracking all data points in experiments that can last up to 1 year. Thus, for Breeding 1, we consider all mice in this experimental group to be infected and that this was associated with insufficient production of anti-OspA antibody by the respective vaccinated dams. Final efficacy of our intervention is determined by the culture of B. burgdorferi from tissues of tick-challenged mice. Figure 5 shows conclusively that pups born to dams vaccinated until 2 weeks post-parturition did not have viable B. burgdorferi in cultures from the heart tissue. This shows that for tick-transmitted B. burgdorferi, if P. leucopus is persistently vaccinated and produces sufficient anti-OspA antibody to be transferred to pups, the offspring is protected for at least 4 weeks post-parturition. In addition, we show that immediately after birth, pups do not have anti-OspA antibodies in serum (Fig. 6). Thus, we infer that P. leucopus pups acquire OspA antibodies during suckling. This observation aligns with our previous studies in C3H mice demonstrating that anti-OspA IgG transfer via milk is a primary mechanism of passive immunity (22).
A potential question may arise from differences in Fig. 2 data showing a significant increase of anti-OspA IgG in pups euthanized on the day of birth from dams vaccinated until parturition in Breeding 2 and data in Fig. 6 showing the absence of anti-OspA IgG in pups euthanized immediately after birth in Breeding 3. This is explained by the fact that pups from the second breeding were born overnight and were euthanized in the morning. Thus, for pups born from the third breeding and to confirm if antibody was transferred via lactation, as we showed previously for C3H mice (22), we euthanized the pups immediately after they dropped to prevent any potential suckling time.
The decline in maternal anti-OspA IgG over time, particularly during lactation (Fig. 7), suggests a depletion of circulating IgG that could have impacted the duration of passive protection provided to pups. This observation is consistent with findings in other models where maternal antibody levels decrease postnatally as antibodies are transferred to offspring (20).
From a reservoir-targeted vaccine intervention perspective, our findings suggest that if maternal anti-OspA antibodies reduce tick colonization efficiency or delay early-stage infection in P. leucopus, this may have broader ecological implications by disrupting pathogen transmission cycles. Further studies are needed to assess the effect of passively transferred anti-OspA antibody in the reduction of nymphal infection prevalence.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Marques AR. 2010. Lyme disease: a review. Curr Allergy Asthma Rep 10:13–20. doi:10.1007/s 11882-009-0077-320425509 · doi ↗ · pubmed ↗
- 2Levine JF, Wilson ML, Spielman A. 1985. Mice as reservoirs of the Lyme disease spirochete. Am J Trop Med Hyg 34:355–360. doi:10.4269/ajtmh.1985.34.3553985277 · doi ↗ · pubmed ↗
- 3Donahue JG, Piesman J, Spielman A. 1987. Reservoir competence of white-footed mice for Lyme disease spirochetes. Am J Trop Med Hyg 36:92–96. doi:10.4269/ajtmh.1987.36.923812887 · doi ↗ · pubmed ↗
- 4Mather TN, Mather ME. 1990. Intrinsic competence of three ixodid ticks (Acari) as vectors of the Lyme disease spirochete. J Med Entomol 27:646–650. doi:10.1093/jmedent/27.4.6462388239 · doi ↗ · pubmed ↗
- 5Tsao J, Barbour AG, Luke CJ, Fikrig E, Fish D. 2001. Osp A immunization decreases transmission of Borrelia burgdorferi spirochetes from infected Peromyscus leucopus mice to larval Ixodes scapularis ticks. Vector Borne Zoonotic Dis 1:65–74. doi:10.1089/15303660175013770512653137 · doi ↗ · pubmed ↗
- 6Richer LM, Brisson D, Melo R, Ostfeld RS, Zeidner N, Gomes-Solecki M. 2014. Reservoir targeted vaccine against Borrelia burgdorferi: a new strategy to prevent Lyme disease transmission. J Infect Dis 209:1972–1980. doi:10.1093/infdis/jiu 00524523510 PMC 4038139 · doi ↗ · pubmed ↗
- 7Gomes-Solecki M. 2014. Blocking pathogen transmission at the source: reservoir targeted Osp A-based vaccines against Borrelia burgdorferi. Front Cell Infect Microbiol 4:136. doi:10.3389/fcimb.2014.0013625309883 PMC 4176399 · doi ↗ · pubmed ↗
- 8Schaible UE, Kramer MD, Eichmann K, Modolell M, Museteanu C, Simon MM. 1990. Monoclonal antibodies specific for the outer surface protein A (Osp A) of Borrelia burgdorferi prevent Lyme borreliosis in severe combined immunodeficiency (scid) mice. Proc Natl Acad Sci USA 87:3768–3772. doi:10.1073/pnas.87.10.37682339119 PMC 53984 · doi ↗ · pubmed ↗