Morpho-phylogenetic evidence reveals novel Bambusicolous fungi from Guizhou Province, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
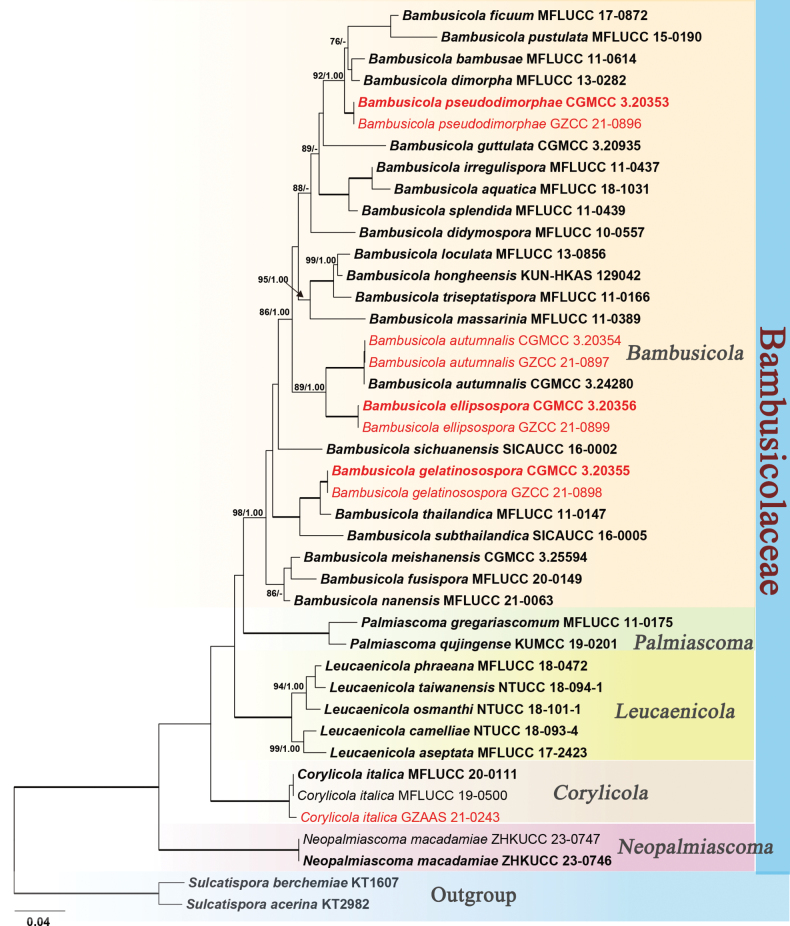 Figure 1
Figure 1 Figure 2
Figure 2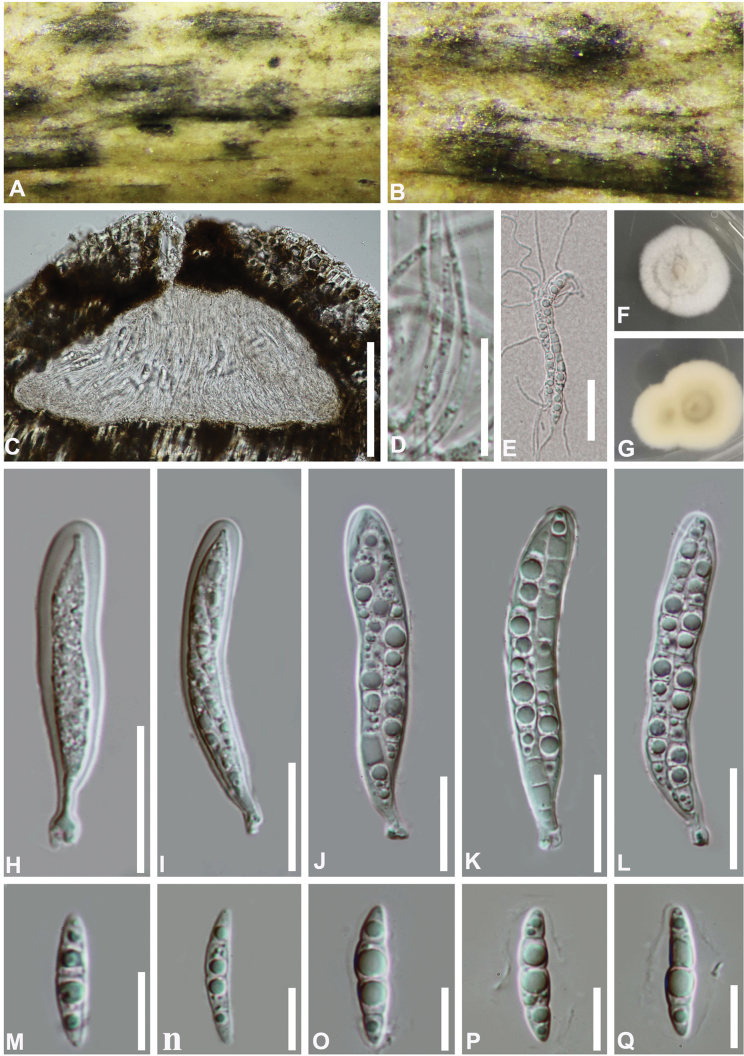 Figure 3
Figure 3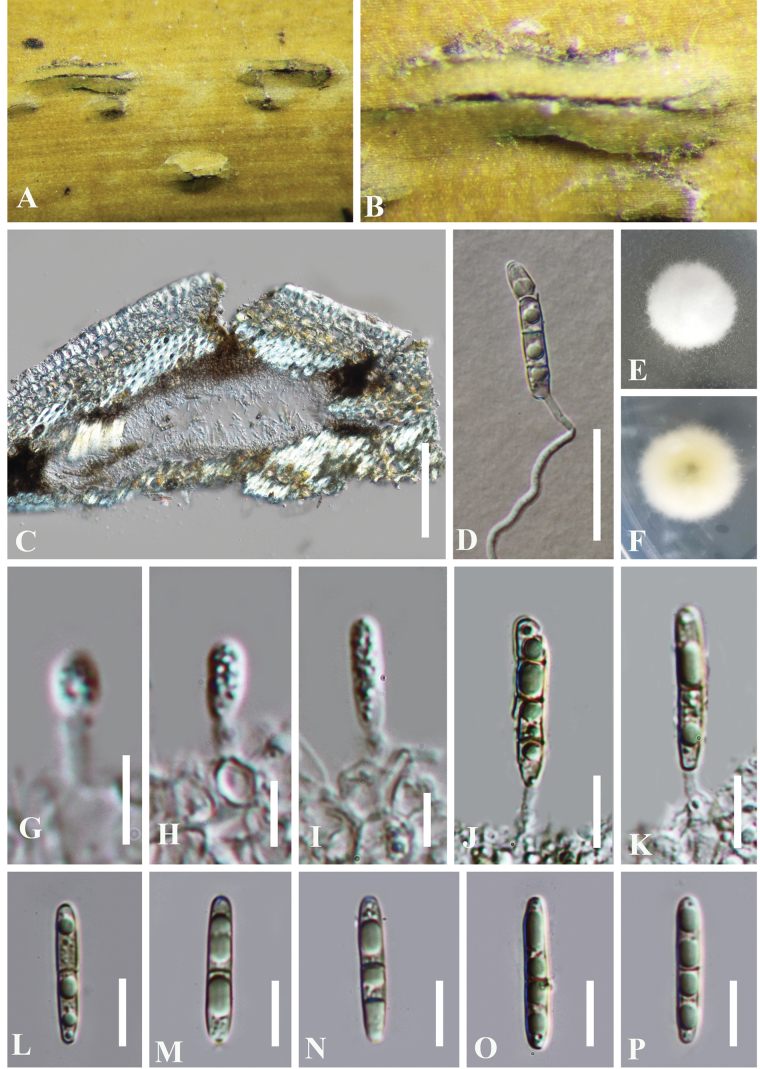 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7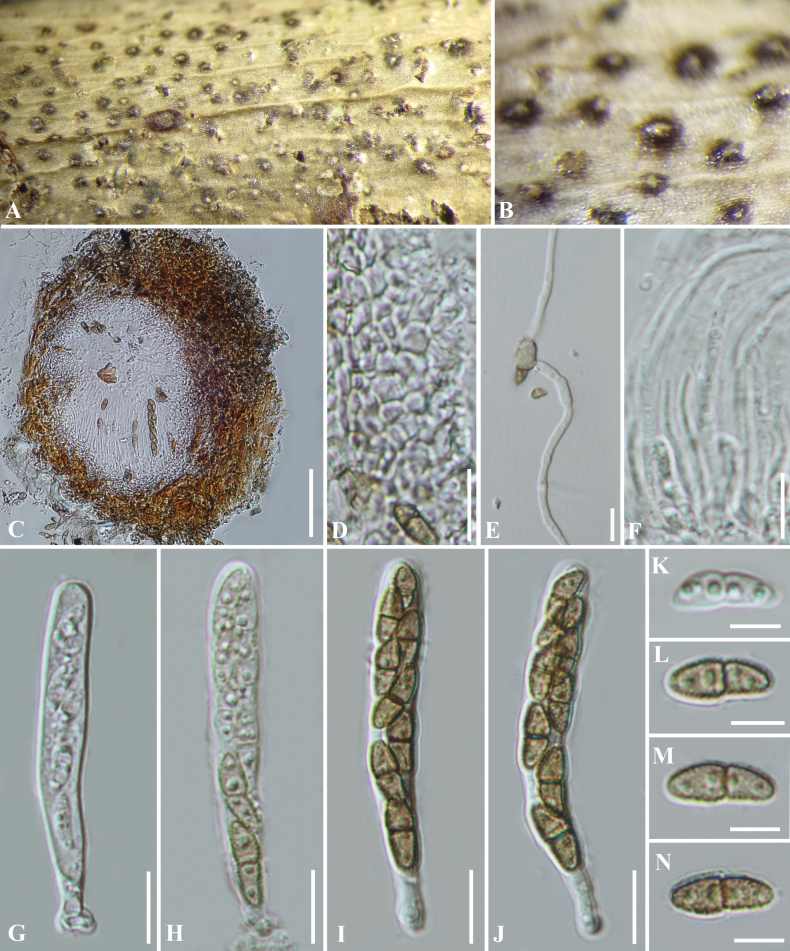 Figure 8
Figure 8| Taxa | Vouchers/Strains | GenBank accession numbers | ||||
|---|---|---|---|---|---|---|
|
|
|
|
|
| ||
|
| MFLUCC 18-1031T |
|
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
| MFLUCC 11-0614T |
|
|
|
|
|
|
| MFLUCC 10-0557T |
|
|
|
|
|
|
| MFLUCC 13-0282T |
|
|
|
| N/A |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
| MFLUCC 17-0872T |
| N/A |
| N/A |
|
|
| MFLUCC 20-0149T |
|
|
|
| N/A |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
| KUN- |
|
|
|
| N/A |
|
| MFLUCC 11-0437T |
|
|
|
|
|
|
| MFLUCC 13-0856T |
|
|
|
|
|
|
| MFLUCC 11-0389T |
|
|
|
|
|
|
|
|
|
| N/A |
| |
|
| MFLUCC 21-0063T | N/A |
|
| N/A | N/A |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
| MFLUCC 15-0190T |
|
|
|
|
|
|
| SICAUCC 16-0002T |
|
|
|
|
|
|
| MFLUCC 11-0439T |
|
|
|
|
|
|
| SICAUCC 16-0005T |
|
|
|
|
|
|
| MFLUCC 11-0147T | N/A |
|
|
|
|
|
| MFLUCC 11-0166T | N/A |
|
|
| N/A |
|
| MFLUCC 19-0500 |
|
|
|
| N/A |
|
| MFLUCC 20-0111T |
|
|
|
|
|
|
|
|
|
| N/A |
| |
|
| MFLUCC 17-2423T |
|
|
|
|
|
|
| NTUCC 18-093-4T |
|
|
|
|
|
|
| NTUCC 18-101-1T |
|
|
|
|
|
|
| MFLUCC 18-0472T |
| N/A |
|
|
|
|
| NTUCC 18-094-1T |
|
|
| N/A | N/A |
|
| ZHKUCC 23-0746T |
|
|
|
|
|
|
| ZHKUCC 23-0747 |
|
|
|
|
|
|
| MFLUCC 11-0175T |
|
|
|
| N/A |
|
| KUMCC 19-0201T |
|
|
|
| N/A |
|
| KT2982 |
|
|
| N/A |
|
|
| KT1607 |
|
|
| N/A |
|
| Morphological characteristics | |||
|---|---|---|---|
| Ascomata | 320–345 μm diam.; 170–230 μm high | 310–400 μm diam.; 90–155 μm high | 326–377 μm diam.; 226–277 μm high |
| Asci | 60–90 × 10–12 μm; (x̄ = 77 × 11 μm, n = 20) | (60–)70– 90(−97) × 10–12(−14) μm; (x̄ = 75.5 × 12 μm, n = 20) | (48–)65–86(–119) × (7.5–)9–12 μm; (x̄ = 75.6 × 11 μm, n = 40) |
| Ascospores | (16–)20–25 × 4–6 μm; (x̄ = 22 × 5 μm, n = 30) | (16–)18– 22(−24) × (3.5–)4–6 μm; (x̄ = 2 0.8 × 5 μm, n = 30) | (20–)21–24(–26) × (4–)5–6(–6.5) μm; (x̄ = 22.6 × 5.7 μm, n = 50) |
| Culture colour | white, reverse pale yellow | white to cream | grey to grey brown |
| Culture texture | felted, with white dense mycelium on the surface | fluffy to floccose | fluffy |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions · Fungal Biology and Applications
Introduction
Bambusoideae, a subfamily of Poaceae, exhibits remarkable species diversity (Shi et al. 2020). As a dominant group within this subfamily, bamboos provide substantial economic, ecological, and social benefits, making them an essential resource in modern forestry and regional economies (Lobovikov et al. 2007; Shukla et al. 2016). Fungi associated with bamboo, collectively called bambusicolous fungi, inhabit various bamboo tissues (Hino 1938; Hyde et al. 2002). These fungi include pathogens that compromise bamboo health (Kuai 1996), endophytes that promote growth and enhance stress resistance (Yan et al. 2023), and saprobes that facilitate organic matter decomposition, nutrient cycling, and energy transformation within bamboo ecosystems (Senanayake et al. 2020; Feng et al. 2021; Jiang et al. 2021, 2022; Liang et al. 2023; Zhang et al. 2023; Yu et al. 2024). With increasing research on bambusicolous fungi, numerous new species continue to be reported (Kwon et al. 2022; Jiang et al. 2022; Yang et al. 2023; Yu et al. 2024).
The family Bambusicolaceae was established by Hyde et al. (2013) to accommodate the type genus Bambusicola based on comprehensive morphological and molecular evidence. Subsequent studies expanded the family to include additional genera, such as Leucaenicola, Longipedicellata, Neobambusicola, and Palmiascoma (Crous et al. 2014; Liu et al. 2015; Jayasiri et al. 2019). However, Neobambusicola and Longipedicellata were later transferred to Sulcatisporaceae and Longipedicellataceae, respectively, following detailed morphological and phylogenetic analysis (Tanaka et al. 2015; Phukhamsakda et al. 2016). More recently, the genera Corylicola, Flavosporella, and Neopalmiascoma have been incorporated into Bambusicolaceae (Wijesinghe et al. 2020; Yu et al. 2024; Zhang et al. 2024), bringing the current number of recognized genera to six: Bambusicola, Corylicola, Flavosporella, Leucaenicola, Neopalmiascoma, and Palmiascoma. Among these, Bambusicola remains the largest and most extensively studied genus. It is morphologically characterized by uni-to multi-loculate, immersed ascomata, bitunicate, cylindrical asci with a short furcate pedicel and a shallow apical ocular chamber and 2–3-seriate, slightly broad-fusiform, 1-septate hyaline ascospores, surrounded by a gelatinous sheath (Dai et al. 2012, 2017; Hyde et al. 2013; Liu et al. 2015). Its asexual morphs exhibit indistinct conidiophores, holoblastic, annelidic, discrete, cylindrical conidiogenous cells, and pale brown to dark brown, 1–3-septate, cylindrical conidia (Dai et al. 2012, 2017; Hyde et al. 2013; Liu et al. 2015; Jayasiri et al. 2019).
Given the increasing recognition of bambusicolous fungal diversity, our study aimed to explore the microfungi associated with decaying bamboo in Guizhou, China. As a result, we identified five species belonging to Bambusicolaceae. To resolve their taxonomic placement and phylogenetic relationships, we conducted morphological comparisons and multi-gene phylogenetic analyses using a concatenated dataset of SSU, ITS, LSU, rpb2, and tef1-α sequences. Our findings revealed three novel species: Bambusicolapseudodimorphae, B.gelatinosospora, and B.ellipsospora. Additionally, we have also identified two known species, namely Bambusicolaautumnalis and Corylicolaitalica. Notably, this study represents the first report of the asexual morph of B.autumnalis, and C.italica is newly recorded from China. We provide comprehensive morphological descriptions, illustrations, and molecular phylogenetic insights, contributing to the growing understanding of bambusicolous fungi.
Materials and methods
Sample collection, morphological studies, and isolation
Decayed culms were collected from Guizhou Province, China. Fungal fruiting bodies were examined using a stereomicroscope (Motic SMZ 168). Freehand sections of ascomata and other fungal structures were mounted in water for microscopic studies and photomicrography. Morphological observations were made using a Nikon ECLIPSE Ni compound microscope fitted with a Nikon DS-Ri2 digital camera. All measurements were made with Tarosoft Image FrameWork software (IFW) (Liu et al. 2010). Photo plates were processed with Adobe Photoshop CS6 software (Adobe Systems, USA). Single spore isolations were carried out following the method in Chomnunti et al. (2014). Type specimens were deposited in the herbarium of Cryptogams Kunming Institute of Botany Academia Sinica (HKAS), Kunming, China, and herbarium of Guizhou Academy of Agriculture sciences (GZAAS), Guiyang, China. Pure cultures were deposited in China General Microbiological Culture Collection Center (CGMCC), Beijing, China, and Guizhou Culture Collection (GZCC), Guiyang, China. Facesoffungi (http://www.facesoffungi.org/) numbers were obtained as in Jayasiri et al. (2015). The new species are registered in Index Fungorum (2025, http://www.indexfungorum.org/).
DNA extraction, PCR amplification and sequencing
Fungal mycelia were scraped from the pure culture which was growing on PDA (Potato Dextrose Agar) for one week at 25 °C in the dark. The total genomic DNA was extracted using an Ezup Column Fungi Genomic DNA Purification Kit (Sangon Biotech, China) from fresh fungal mycelia. Five gene regions, small subunit rDNA (SSU), internal transcribed spacer (ITS), large subunit rDNA (LSU), RNA polymerase II subunit 2 (rpb2) and translation elongation factor 1-alpha (tef1-α) were amplified using the primer pairs NS1/NS4 (White et al. 1990), ITS4/ITS5, LR0R/LR5 (Vilgalys and Hester 1990), fRPB2-5f /fRPB2-7cR (Liu et al. 1999) and ef1-983F/ef1-2218R (Rehner and Buckley 2005), respectively. Polymerase chain reaction (PCR) was carried out in a 25 μL reaction volume containing 12.5 μL 2 × PCR Master Mix (Sangon Biotech, China), 9.5 μL ddH_2_O, 1 μL of each primer and 1 μL DNA template. The amplification conditions for SSU, ITS, LSU and tef1-α gene regions followed Tennakoon et al. (2016), for rpb2 gene regions followed Du et al. (2024). PCR products were examined using 1.2% agarose electrophoresis gel stained with ethidium bromide and sequenced by Sangon Biotech (Shanghai) Co., Ltd, China. Newly generated nucleotide sequences were submitted in GenBank (Table 1).
Phylogenetic analyses
Analyses of Bambusicolaceae were performed respectively. Both phylogenetic analyses were performed based on SSU, ITS, LSU, rpb2 and tef1-α sequence data. The representative strains of Bambusicolaceae (Table 1) were referred to BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) results and relevant publications (Yu et al. 2022, 2024; Liang et al. 2023; Zhang et al. 2024). Sequences were aligned using MAFFT v. 7 (Katoh and Standley 2013). Manual adjustments were performed when necessary using BioEdit v. 7.0 (Hall 1999). Phylogenetic analyses of maximum likelihood (ML) and bayesian inference (BI) were carried out as detailed in Dissanayake et al. (2020) and performed with raxmlGUI v. 1.3 (Silvestro and Michalak 2012) and MrBayes v3.1. (Huelsenbeck and Ronquist 2001; Zhaxybayeva and Gogarten 2002; Nylander 2004). Phylogenetic trees were visualized with FigTree v1.4.2 (Rambaut 2012) and edited using Adobe Illustrator 2021 (2.6.0.44) and Adobe Photoshop CS6 software (Adobe Systems, USA).
Results
Phylogeny
To determine the phylogenetic placements of the new collections in this study, the combined SSU, ITS, LSU, rpb2 and tef1-α data sets, comprising 42 taxa with Sulcatisporaberchemiae KT1607 and Sulcatisporaacerina KT2982 as the outgroup taxa. The dataset comprises 4, 641 characters (SSU: 1-1001; ITS: 1002-1896; LSU: 1897-2701; rpb2: 2702-3691; tef1-α: 3692-4641) after alignment, including gaps. Maximum likelihood and bayesian analyses were performed, respectively, and both methods yielded consistent topologies. The best-scoring RAxML tree (Fig. 1) was obtained with a final likelihood value of -23443.659740. Estimated base frequencies were as follows: A = 0.243090, C = 0.252645, G = 0.271896, T = 0.228350; substitution rates AC = 1.173578, AG = 2.804498, AT = 0.917016, CG = 1.080920, CT = 5.912507, GT = 1.000000; The gamma distribution shape parameter alpha is equal to 0.305487 and the Tree-Length equal to 0.166660.
Phylogram generated from maximum likelihood analysis based on a combined dataset of SSU, ITS, LSU, rpb2 and tef1-α sequences. Bootstrap support values for ML (≥75%) and bayesian posterior probabilities (≥0.95) are given at the nodes (ML BS/PP). Branches with 100% ML BS and 1.00 PP are thickened. New strains are shown in red. The tree is rooted with Sulcatisporaberchemiae KT1607 and Sulcatisporaacerina KT2982, and ex-type strains are in bold.
Phylogenetic analyses showed that the nine newly obtained strains of Bambusicolaceae clustered into five clades and can be recognized as three new species (Bambusicolaellipsospora, B.gelatinosospora, B.pseudodimorphae) and two known species (B.autumnalis, Corylicolaitalica) (Fig. 1).
Taxonomy
Bambusicola
ellipsospora
Taxon classificationFungiPleosporalesBambusicolaceae
Y. Feng, Z.Y. Liu & Jian K. Liu sp. nov.
DFB58706-5BD2-5D60-8A27-987323760EF0
Index Fungorum: IF903223
Facesoffungi Number: FoF15907
Holotype.
HKAS 112592.
Etymology.
The epithet refers to the ellipsoidal ascospores.
Description.
Saprobic on dead bamboo culms. Sexual morph: Ascomata 215–363 μm diam., 180–288 μm high, solitary, scattered to gregarious, immersed to erumpent, later becoming superficial, forming brown to dark brown, lenticular spots on the host surface with a slit. Peridium comprising several layers of cells of textura angularis, inner layers comprising hyaline to dark brown, outer layers composed of thick, dark brown to black cells. Hamathecium of dense, 0.5–1.8 μm wide, filamentous, branched, septate, smooth-walled, trabecular pseudoparaphyses, anastomosing at the apex, embedded in a hyaline, gelatinous matrix. Asci 70–92 × 15–21 μm (x̄ = 82 × 17 μm, n = 20), 8-spored, bitunicate, fissitunicate, broadly cylindrical to cylindri-clavate, with a short pedicel, apically rounded with a well-developed ocular chamber. Ascospores (18–)19–24(–25) × (5–)6–8(–9) μm (x̄ = 22 × 7 μm, n = 30), overlapping, biseriate, hyaline, straight, ellipsoidal to fusiform, with obtuse ends, 1-septate, constricted at the septum, smooth-walled with 4 large guttules, surrounded by a thin-gelatinous sheath. Asexual morph: Undetermined.
Bambusicolaellipsospora (HKAS 112592, holotype) A, B appearance of ascomata on the host C vertical section of an ascoma D peridium E pseudoparaphyses F, G cultures on PDAF from above G from below H–L asci M–Q ascospores. Scale bars: 100 µm (C); 10 µm (D, E, M–Q); 20 µm (H–L).
Culture characteristics.
Ascospores germinating on PDA within 12 h. Colonies on PDA slow growing, 13 mm diam. after 10 d at 25 °C in the dark, circular, white, velvety, with dense mycelium on the surface; in reverse brown in the center, becoming paler to white at the entire margin.
Material examined.
China • Guizhou Province, Chishui City. On dead culms of bamboo, 10 July 2019, Yao Feng CS018 (HKAS 112592, holotype; GZAAS 21-0503, isotype), ex-type living cultures CGMCC 3.20356 = GZCC 21-0803; ibid., CS036 (GZAAS 21-0391, paratype), living culture GZCC 21-899.
Notes.
Phylogenetic analyses revealed that the two strains of Bambusicolaellipsospora (CGMCC 3.20356 and GZCC 21-0899) formed a distinct clade, sister to B.autumnalis (CGMCC 3.24280, CGMCC 3.20354, GZCC 21-0897) (Fig. 1). Bambusicolaellipsospora can be distinguished from B.autumnalis based on sequence divergence in tef1-α (874/927 bp) and rpb2 (699/784 bp). Additionally, morphological differences further support their distinction, as B.ellipsospora produces ellipsoidal to fusiform ascospores with obtuse rounded ends, while B.autumnalis has fusiform and slightly curved ascospores with narrower acute ends (Liang et al. 2023). Moreover, the ascospore of B.ellipsospora collected in this study shows distinct constriction at the septum, whereas that of B.autumnalis only has slight constriction. Based on morphological and phylogenetic evidence, B.ellipsospora is recognized as a novel species.
Bambusicola
gelatinosospora
Taxon classificationFungiPleosporalesBambusicolaceae
Y. Feng, Z.Y. Liu & Jian K. Liu sp. nov.
35975FA1-8AEE-5A35-8D46-F6C6ABB14D31
Index Fungorum: IF903224
Facesoffungi Number: FoF15908
Holotype.
HKAS 112599.
Etymology.
The epithet refers to the gelatinous sheath of ascospores.
Description.
Saprobic on dead bamboo culms, forming raised, black spots on the host surface. Sexual morph: Ascomata 320–345 μm diam, 170–230 μm high, solitary, scattered to gregarious, immersed, subglobose with a flattened base. Peridium comprising several layers of cells of textura angularis, less distinguished from the host tissue. Hamathecium of dense, 1–2 μm wide, filamentous, branched, septate, smooth-walled, trabecular pseudoparaphyses, anastomosing at the apex, embedded in a hyaline, gelatinous matrix. Asci 60–90 × 10–12 μm (x̄ = 77 × 11 μm, n = 20), 8-spored, bitunicate, fissitunicate, broadly cylindrical to cylindrical-clavate, with a short pedicel, apically rounded with a well-developed ocular chamber. Ascospores (16–)20–25 × 4–6 μm (x̄ = 22 × 5 μm, n = 30), overlapping, biseriate, hyaline, fusiform, slightly bent with acute ends, 3-septate, slightly constricted at the septa, smooth-walled, guttulate, surrounded by a gelatinous sheath. Asexual morph: Undetermined.
Bambusicolagelatinosospora (HKAS 112599, holotype) A, B appearance of ascomata on the host C vertical section of an ascoma D pseudoparaphyses E germinated ascospores F, G cultures on PDAF from above G from below H–L asci M–Q ascospores. Scale bars:100 µm (C); 30 µm (E); 20 µm (H–L); 15 µm (D, M–Q).
Culture characteristics.
Colonies on PDA slow growing, reaching 15 mm diam. after 10 d at 25 °C in dark. circular, felted, with white dense mycelium on the surface. In reverse pale yellow, becoming brown from the center, with an entire margin.
Material examined.
China • Guizhou Province, Xingyi City. On dead stem of bamboo, 5 September 2019, Yao Feng, XY19-11 (HKAS 112599, holotype; GZAAS 21-0502, isotype), ex-type living cultures CGMCC 3.20355 = GZCC 21-0802; ibid., on dead branches on bamboo, 5 September 2019, Yao Feng, XY19a (GZAAS 21-0390; paratype), living cultures GZCC21-0898.
Notes.
Phylogenetic analyses revealed that the two strains of Bambusicolagelatinosospora (CGMCC 3.20355 and GZCC 21-0898) clustered with B.thailandica (MFLUCC 11-0147) and B.subthailandica (SICAUCC 16-0005) as a distinct lineage (Fig. 1). Morphologically, B.gelatinosospora closely resembles B.thailandica and B.subthailandica, sharing key characteristics such as ostiolate ascomata, cylindrical to cylindri-clavate asci, and hyaline, fusiform, 3-septate ascospores with acute ends (Dai et al. 2017; Yang et al. 2019). Although these species are morphologically indistinguishable, their cultures exhibit differences in colors and colony morphology (Table 2). Additionally, they can be differentiated by their minimal sequence similarities. In a comparison of ITS, LSU, rpb2 and tef1-α nucleotides, B.gelatinosospora (CGMCC 3.20355) has 98%, 99%, 97% and 99% similarity, in ITS (702/713 bp, 1 gap), LSU (817/819 bp, no gaps), rpb2 (949/974 bp, no gaps), and tef1-α (914/920 bp, no gap), which is different from B.thailandica (MFLUCC 11-0147). These findings combined with its morphological and molecular distinctiveness strongly supported the conclusion that Bambusicolagelatinosospora represents a new species within the genus Bambusicola.
Bambusicola
pseudodimorphae
Taxon classificationFungiPleosporalesBambusicolaceae
Y. Feng, Z.Y. Liu & Jian K. Liu sp. nov.
AB9367A4-1559-5985-A9C9-E75214A6AA8B
Index Fungorum: IF903225
Facesoffungi Number: FoF15909
Holotype.
HKAS 112597.
Etymology.
Species epithet refers to the similar conidia to Bambusicoladimorpha which have two types in culture.
Description.
Saprobic on bamboo culms. Sexual morph: Undetermined. Asexual morph: Conidiomata initially embedded in the epidermis of bamboo culms and later breaking through the epidermis,solitary to gregarious, subglobose with a flattened base, brown to dark brown. 78–161 μm high, 180–270 μm diam. Conidiomatal wall comprising 3–5 layers of cells of textura angularis, with dark brown outer layers and hyaline inner layers, less distinguished from the host tissue. Conidiophores reduced to conidiogenous cells. Conidiogenous cells 9–32 × 1–2 μm (x̄ = 23 × 2 μm, n = 15), holoblastic, monoblastic, cylindrical, smooth, hyaline. Conidia 24–27 × 3–4 μm (x̄ = 26 × 4 μm, n = 30), initially hyaline to pale brown, aseptate, becoming yellowish brown, 2–3-septate when maturity, cylindrical, obtuse at both ends, smooth-walled, straight, guttulate.
Bambusicolapseudodimorphae (HKAS 112597, holotype) A, B conidiomata on the host C vertical section of a conidioma D germinated conidium E, F cultures on PDAE from above F from below G−K conidiogenous cells and developing conidia L−P conidia. Scale bars: 100 µm (C); 20 µm (D); 5 µm (G−I); 10 µm (J−P).
Culture characteristics.
Colonies on PDA slow growing, 16 mm diam. after 10 d at 25 °C in dark, circular, with an entire margin, white, fluffy, with dense mycelium on the surface. Reverse white with a yellowish center. After being cultivated on PDA for two months, Bambusicolapseudodimorphae generated two types of conidia, macro- and microconidia. Macroconidia 20–26(–29) × 3–5 μm (x̄ = 24 × 4 μm, n = 30), cylindrical, with obtuse and rounded ends, 3-septate, pale brown to brown, smooth-walled. Microconidia 3–4 × 1–3 μm (x̄ = 4 × 2 μm, n = 30), ellipsoidal, pale brown, aseptate.
Reproduced Bambusicolapseudodimorphae (CGMCC 3.20353, ex-type) A, B colony on PDAC−F conidiogenous cells with conidia G−J macroconidia K microconidia. Scale bars: 10 µm (C−K).
Material examined.
China • Guizhou Province, Xingyi City. On dead bamboo stems, 5 September 2019, Yao Feng, XY19-3 (HKAS 112597, holotype; GZAAS 21-0501, isotype), ex-type living cultures CGMCC 3.20353 = GZCC21-0801; ibid., 6 September 2019, Yao Feng, XY19b (GZAAS 21-0388, paratype), living culture GZCC 21-0896.
Notes.
Phylogenetic analyses revealed that the two strains of Bambusicolapseudodimorphae clustered with B.bambusae, B.dimorpha, B.ficuum, and B.pustulata, forming a distinct lineage within Bambusicola (Fig. 1). Morphologically, B.pseudodimorphae closely resembles is similar to B.dimorpha in having two types of conidia. However, B.pseudodimorphae differs by having smaller conidiomata (78–161 × 180–270 μm vs. 200–250 × 210–340 μm) and longer conidia (24–27 μm long, 3-septate vs. 13–21 μm long, 1-septate) (Thambugala et al. 2017). Since our collection is an asexual morph, and the other three species (B.bambusae, B.ficuum, and B.pustulata) are known as sexual morphs, direct comparisons between them are not feasible. Molecular analyses further support the distinction of B.pseudodimorphae as a novel species. The tef1-α and rpb2 sequences of B.pseudodimorphae differ from those of B.bambusae by 3% (32/924 bp) and 3% (35/1001 bp) nucleotides, respectively. Additionally, the rpb2 sequence of B.pseudodimorphae showed 96% (945/984 bp, one gap) and 97% (853/880 bp, no gap) sequence similarity with B.pustulata and B.dimorpha, respectively. Based on the morphology and molecular evidence, B.pseudodimorphae is identified as a new species following the species delineation guidelines of Jeewon and Hyde (2016).
Bambusicola
autumnalis
Taxon classificationFungiPleosporalesBambusicolaceae
R.R. Liang, S.N. Zhang & Jian K. Liu, Phytotaxa 601(3): 208 (2023)
94562EFE-AD97-5611-A9EB-0A00DBC9F8DF
Description.
Saprobic on dead bamboo culms. Sexual morph: Ascomata 263–370 μm diam., 185–202 μm high, solitary, scattered to gregarious, initially growing under the host surface, breaking through the surface after maturity, with cracks on the host, subglobose with a flattened base, some arranged in rows, brown to dark brown. Peridium comprising several layers of cells of textura angularis, with dark brown outer layers and hyaline inner layers. Hamathecium of dense, 1–2 μm wide, filamentous, branched, indistinctly septate, smooth-walled, trabeculate pseudoparaphyses, anastomosing at the apex, embedded in a hyaline, gelatinous matrix. Asci 64–95(−102) × (10–)11−14 μm (x̄ = 83 × 12 μm, n = 20), 8-spored, bitunicate, broadly cylindrical to cylindrical-clavate, with a short pedicel, apically rounded with a well-developed ocular chamber. Ascospores (24–)26–29 × 4–6 μm (x̄ = 26 × 5 μm, n = 30), overlapping, biseriate, hyaline, fusiform, with acute ends, 1-septate, slightly constricted at the septa, asymmetric, slightly curved, smooth-walled with 2–4 guttules, surrounded by a thin, inconspicuous mucilaginous sheath. Asexual morph: Coelomycetous, produced on PDA. Conidiomata pycnidial, solitary to gregarious, superficial. Conidiogenous cells 5–17 × 2–4 μm (x̄ = 9 × 3 μm, n = 15), holoblastic, integrated or discrete, subcylindrical to cylindrical, hyaline, smooth, occasionally with an enlarged structure at the top. Conidia with two types, macro- and microconidia. Macroconidia 23–31(–34) × 4–6(–7) μm (x̄ = 28 × 5 μm, n = 30), cylindrical to ellipsoidal, with rounded ends, 3-septate, pale brown to brown, slightly constricted at the septa, smooth. Microconidia 3–6 × 1–2(–3) μm (x̄ = 5 × 2 μm, n = 30), subglobose to ellipsoidal, with rounded to obtuse ends, pale brown, aseptate, smooth-walled.
Bambusicolaautumnalis (HKAS 112598) A, B appearance of ascomata on the host C pseudoparaphyses D vertical section of an ascoma E peridium F germinated ascospore G, H cultures on PDAG from above H from below I–M asci N–R ascospores. Scale bars: 100 µm (D); 25 µm (E, F); 20 µm (I–M); 15 µm (C, N–R).
Culture characteristics.
Colonies on PDA reaching 16 mm diam. after 10 d at 25 °C in dark, white, circular, velvety, with uneven margin. Reverse white with a yellowish center.
Material examined.
China • Guizhou Province, Xingyi City. On dead bamboo stems, 5 September 2019, Yao Feng, XY19-4 (HKAS 112598,GZAAS 21-0500, living cultures CGMCC 3.20354 = GZCC 21-0800; ibid., 6 September 2019, Yao Feng, XY19-1 (GZAAS 21-0389), living culture GZCC 21-0897.
Reproduced Bambusicolaautumnalis (CGMCC 3.20354) A, B colony on PDAC−F conidiogenous cells with macroconidia G−K macroconidia L−N conidiogenous cells with microconidia O microconidia. Scale bars: 10 µm (C−O).
Notes.
Liang et al. (2023) described Bambusicolaautumnalis as a novel species based on its sexual morph and phylogenetic analysis. The phylogenetic results (Fig. 1) confirmed that our new collections are conspecific with B.autumnalis, and this study expands upon their findings by providing a detailed description of the asexual morph, thereby enhancing the taxonomic understanding of this species.
Corylicola
italica
Taxon classificationFungiPleosporalesBambusicolaceae
Wijesinghe, Camporesi, Yong Wang bis & K.D. Hyde, Biodiversity Data Journal 8 (e55957): 8 (2020)
ED4AE79B-9061-5BFA-9B15-8B1973B7873F
Description.
Saprobic on the branches of Prunusserrulata. Sexual morph: Ascomata 186–292 µm high, 179–267 µm diam., solitary, scattered to gregarious, immersed or erumpent, uniloculate with an ostiole. Peridium 15–30 µm, comprising several layers of cells of textura angularis, with dark brown outer layers and hyaline inner layers. Hamathecium of dense, 1–2 μm wide, filamentous, branched, septate, smooth-walled, cellular pseudoparaphyses, anastomosing at the apex, embedded in a hyaline, gelatinous matrix. Asci 53–66 × 6–8 µm (x̄ = 61 × 7 µm, n = 20), 8-spored, bitunicate, fissitunicate, broadly cylindrical to cylindrical-clavate, with a short pedicel, apically rounded. Ascospores 8–17 × 3–4 µm (x̄ = 13 × 4 µm, n = 30), overlapping, hyaline, fusiform, broader at the top and narrower at the bottom, with a septum in the middle that is deeply constricted, translucent to light brown or brown, rough. Asexual morph: Undetermined.
Corylicolaitalica (GZAAS 21-0243) A, B appearance of ascomata on host C vertical section of an ascoma D peridium E germinated ascospore F pseudoparaphyses G−J asci K−N ascospores. Scale bars: 50 µm (C); 10 µm (D−J); 5 µm (K−N).
Culture characteristics.
Ascospores germinating on PDA within 12 h. Colonies on PDA slow-growing, 9 mm diam. after 10 d at 25 °C in the dark, circular, irregular edges, with dense grayish white mycelium on the surface, in reverse brown to black.
Material examined.
China • Guizhou Province, Xingyi City. On dead stem of Prunusserrulata, 10 July 2018, Yao Feng, nky 141 (GZAAS 21-0243), living culture GZCC 21-0757.
Notes.
The morphology and phylogenetic results confirm that our new collections are identical to Corylicolaitalica. The genus Corylicola was established by Wijesinghe et al. (2020) with C.italica as the type species. This species was originally described from a hanging branch of Corylusavellana in Italy (Wijesinghe et al. 2020). This study represents the first record of C.italica in China.
Discussion
This study presents a comprehensive taxonomic investigation of newly isolated fungal species associated with bamboo, contributing to the growing knowledge of fungal diversity within this unique ecological niche. We describe two new sexual morphs, Bambusicolaellipsospora and B.gelatinosospora, as well as a new asexual morph, B.pseudodimorphae. Additionally, we provide a holomorphic description of B.autumnalis and report Corylicolaitalica for the first time in China. Within the genus Bambusicola, eleven species have been described based on their sexual morphs, including B.aquatica, B.autumnalis, B.bambusae, B.ficuum, B.fusispora, B.hongheensis, B.loculata, B.pustulata, B.subthailandica, B.thailandica, and B.meishanensis. In contrast, five species, B.guttulata, B.irregulispora, B.nanensis, B.sichuanensis, and B.splendida, are classified based solely on their asexual morphs (Dai et al. 2015; Yang et al. 2019; Brahmanage et al. 2020; Monkai et al. 2021; Phukhamsakda et al. 2022; Yu et al. 2022, 2024; Liang et al. 2023). Notably, holomorph are only known for four species: B.didymospora, B.dimorpha, B.massarinia, and B.triseptatispora (Dai et al. 2012, 2017; Thambugala et al. 2017). An intriguing observation in Bambusicola is the ability of certain species, such as B.dimorpha, B.nanensis, B.sichuanensis, and B.pseudodimorphae, to produce both macroconidia and microconidia in culture (Thambugala et al. 2017; Yang et al. 2019; Phukhamsakda et al. 2022; this study). This phenomenon may be linked to environmental factors, particularly nutrient depletion in the growth medium, which could induce the production of conidia in different forms as an adaptive response. Further studies are required to elucidate the genetic and physiological mechanisms regulating this morphological plasticity.
Currently, Bambusicola comprises 20 species (Species Fungorum, accessed 12 February 2025), with 19 species reported from dead bamboo (Dai et al. 2012, 2015, 2017; Dong et al.2020; Thambugala et al. 2017; Monkai et al. 2021; Yang et al. 2019; Yu et al. 2022, 2024, Liang et al. 2023) and only Bambusicolaficuum identified from Ficus (Boonmee et al. 2021). Examining of species distributions within the genus suggests a strong host preference, with most species exhibiting specificity for bamboo (Dai et al. 2012, 2015, 2017; Dong et al.2020; Thambugala et al. 2017; Yang et al. 2019; Monkai et al. 2021; Yu et al. 2022, 2024; Liang et al. 2023). This raises important ecological questions: Do Bambusicola species establish specialized relationships with bamboo through physiological and biochemical mechanisms? Furthermore, does this host specificity make them particularly vulnerable to environmental changes such as climate fluctuations, pest and disease outbreaks, or habitat destruction? Understanding the ecological roles of Bambusicola species is essential for assessing their contributions to bamboo ecosystems. Future research should focus on the mechanisms underlying their interactions with bamboo, including potential mutualistic or pathogenic associations. Such studies will contribute to taxonomic resolution and provide insights into conservation strategies to preserve fungal diversity and maintain ecosystem stability.
Supplementary Material
XML Treatment for Bambusicola ellipsospora
XML Treatment for Bambusicola gelatinosospora
XML Treatment for Bambusicola pseudodimorphae
XML Treatment for Bambusicola autumnalis
XML Treatment for Corylicola italica
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Boonmee S Wanasinghe DN Calabon MS Huanraluek N Chandrasiri SKU Jones GEB Rossi W Leonardi M Singh SK Rana S Singh PN Maurya DK Lagashetti AC Choudhary D Dai YC Zhao CL Mu YH Yuan HS He SH Phookamsak R Jiang HB Martín MP Dueñas M Telleria MT Kałucka IL Jagodziński AM Liimatainen K Pereira DS Phillips AJL Suwannarach N Kumla J Khuna S Lumyong S Potter TB Shivas RG Sparks AH Vagheff N Abdel-Wahab MA Abdel-Aziz FA Li GJ Lin WF Singh U Bhatt RP Lee HB Nguyen TTT Kirk PM Dutta AK Acharya K Sarma VV Niranjan M Rajeshkumar KC Ashtekar N Lad S Wijayawardene NN Bhat DJ Xu RJ Wijesinghe SN Shen HW Luo ZL Zhang JY Sysouphan · doi ↗ · pubmed ↗
- 2Brahmanage RS Dayarathne MC Wanasinghe DN Thambugala KM Jeewon R Chethana KWT Samarakoon MC Tennakoon DS De Silva NI Camporesi E Raza M Yan JY Hyde KD (2020) Taxonomic novelties of saprobic Pleosporales from selected dicotyledons and grasses.Mycosphere 11: 2481–2541. 10.5943/mycosphere/11/1/15 · doi ↗
- 3Chomnunti P Hongsanan S Aguirre-Hudson B Tian Q Peršohm D Dhami MK Alias AS Xu JC Liu XZ Stadler M Hyde KD (2014) The sooty moulds.Fungal Diversity 66(1): 1–36. 10.1007/s 13225-014-0278-5 · doi ↗
- 4Crous PW Wingfield MJ Schumacher RK Summerell BA Giraldo A GenéJ Guarro J Wanasinghe DN Hyde KD Camporesi E Jones EBG Thambugala KM Malysheva EF Malysheva VF Acharya KÁlvarez J Alvarado P Assefa A Barnes CW Bartlett JS Blanchette RA Burgess TI Carlavilla JR Coetzee MPA Damm U Decock C Aden Breeÿen Ade Vries B Dutta AK Holdom DG Rooney-Latham S Manjón JL Marincowitz S Mirabolfathy M Moreno G Nakashima C Papizadeh M Shahzadeh Fazeli SA Amoozegar MA Romberg MK Shivas RG Stalpers JA Stielow B Stukely MJC Swart WJ Tan Y Pvan der Bank M Wood AR Zhang Y Groenewald JZ (2014) Fungal Planet descript · doi ↗ · pubmed ↗
- 5Dai DQ Bhat DJ Liu JK Chukeatirote E Zhao RL Hyde KD (2012) Bambusicola, a new genus from bamboo with asexual and sexual morphs. Cryptogamie.Mycologie 33(3): 363–379. 10.7872/crym.v 33.iss 3.2012.363 · doi ↗
- 6Dai DQ Bahkali AH Li WJ Bhat DJ Zhao RL Hyde KD (2015) Bambusicolaloculata sp. nov. (Bambusicolaceae) from bamboo.Phytotaxa 213: 122–130. 10.11646/phytotaxa.213.2.5 · doi ↗
- 7Dai DQ Phookamsak R Wijayawardene NN Li WJ Bhat DJ Xu JC Taylor JE Hyde KD Chukeatirote E (2017) Bambusicolous fungi.Fungal Diversity 82: 1–105. 10.1007/s 13225-016-0367-8 · doi ↗
- 8Dissanayake AJ Bhunjun CS Maharachchikumbura SSN Liu JK (2020) Applied aspects of methods to infer phylogenetic.Mycosphere: Journal of Fungal Biology 11(1): 2652–2676. 10.5943/mycosphere/11/1/18 · doi ↗