Exploration of Hiptage (Malpighiaceae) diversity in Vietnam reveals a new species with wingless fruits

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
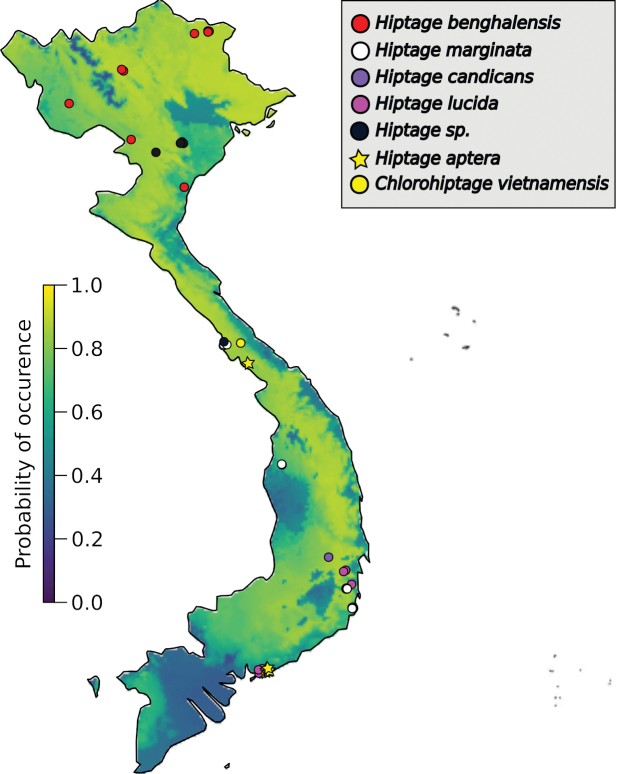 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3| Taxon name | Location | GenBank accession | Year | Geographical coordinates | Original_ID |
|---|---|---|---|---|---|
|
| M’Drak District, Dak Lak Province, Vietnam |
| 2022 |
| 01_220324_Daklak |
|
| Ninh Hoa District, Khanh Hoa Province, Vietnam |
| 2022 |
| 03_220512_KhanhHoa |
| Long Dien District, Ba Ria – Vung Tau Province, Vietnam |
| 2022 |
| 04_220516_VungTau | |
| Long Dien District, Ba Ria – Vung Tau Province, Vietnam |
| 2022 |
| 06_220516_VungTau | |
| Dat Do District, Ba Ria – Vung Tau Province, Vietnam |
| 2022 |
| 07_220516_VungTau | |
| Cam Lam District, Khanh Hoa Province, Vietnam |
| 2022 |
| 14_220707_KhanhHoa | |
| Cam Lam District, Khanh Hoa Province, Vietnam |
| 2023 |
| 37_230625_KhanhHoa | |
|
| Dat Do District, Ba Ria – Vung Tau Province, Vietnam |
| 2024 |
| 08_240118_VungTau |
| Da Krong District, Quang Tri Province, Vietnam |
| 2022 |
| 19_221201_QuangTri | |
| Dat Do District, Ba Ria – Vung Tau Province, Vietnam |
| 2024 |
| 61_240408_VungTau | |
| Dat Do District, Ba Ria – Vung Tau Province, Vietnam |
| 2024 |
| 62_240601_VungTau | |
| Tan Thanh District, Ba Ria – Vung Tau Province, Vietnam |
| 2024 |
| 64_240623_VungTau | |
| Dat Do District, Ba Ria – Vung Tau Province, Vietnam |
| 2024 |
| 65_240623_VungTau | |
|
| Van Yen District, Yen Bai Province, Vietnam |
| 2022 |
| 11_220528_YenBai |
| Van Yen District, Yen Bai Province, Vietnam |
| 2022 |
| 12_220528_YenBai | |
| Quang Hoa District, Cao Bang Province, Vietnam |
| 2023 |
| 29_230414_CaoBang | |
| Nguyen Binh District, Cao Bang Province, Vietnam |
| 2023 |
| 30_230414_CaoBang | |
| Thuan Chau District, Son La Province, Vietnam |
| 2023 |
| 52_231028_SonLa | |
| Van Ho District, Son La Province, Vietnam |
| 2023 |
| 53_231030_SonLa | |
| Quang Hoa District, Cao Bang Province, Vietnam |
| 2024 |
| 57_240402_CaoBang | |
| Quang Hoa District, Cao Bang Province, Vietnam |
| 2024 |
| 58_240402_CaoBang | |
| Quang Hoa District, Cao Bang Province, Vietnam |
| 2024 |
| 59_240402_CaoBang | |
| Van Yen District, Yen Bai Province, Vietnam |
| 2024 |
| 60_240404_YenBai | |
| Hahé Province, Seychelles |
| 1983 | - | MNHN_P00904869 | |
| Hahé Province, Seychelles |
| 1983 | - | MNHN_P00904870 | |
| Jardin botanique Mahé, Hahé Province, Seychelles |
| 1982 | - | MNHN_P01033130 | |
| Laos |
| 1925 | - | MNHN_P04783895 | |
| West flank of Piton du Fouge, Mauritius |
| 1975 | - | MNHN_P04868852 | |
| Cuc Phuong National Park, Nho Quan District, Ninh Binh Province, Vietnam |
| 2002 |
| MNHN_P05474376 | |
| East of Urugala, Kandy District, Central Province, Sri Lanka |
| 1973 | - | MNHN_P05598587 | |
| La Grande Chaloupe, La Possession, La Réunion |
| 2023 |
| 35_Reunion_2303 | |
| Bassin Boeuf, Sainte-Suzanne, La Réunion |
| 2021 |
| REU_BassinBoeuf_1 | |
| Bassin Boeuf, Sainte-Suzanne, La Réunion |
| 2021 |
| REU_BassinBoeuf_1bis | |
| Bassin Boeuf, Sainte-Suzanne, La Réunion |
| 2021 |
| REU_BassinBoeuf_2 | |
| La Grande Chaloupe, La Possession, La Réunion |
| 2021 |
| REU_GrandeChaloupe_1 | |
| La Grande Chaloupe, La Possession, La Réunion |
| 2021 |
| REU_GrandeChaloupe_2 | |
| La Grande Chaloupe, La Possession, La Réunion |
| 2021 |
| REU_GrandeChaloupe_3 | |
| Le Petit Serré, Saint-Louis, La Réunion |
| 2021 |
| REU_PetitSerre_1 | |
| Le Petit Serré, Saint-Louis, La Réunion |
| 2021 |
| REU_PetitSerre_2 | |
|
| Ba Thuoc District, Thanh Hoa Province, Vietnam |
| 2023 |
| 31_230414_ThanhHoa |
| Chu Mon Ray National Park, Sa Thay District, Kon Tum Province, Vietnam |
| 2023 |
| 33_230414_KonTum | |
| Cam Lam District, Khanh Hoa Province, Vietnam |
| 2023 |
| 36_230624_KhanhHoa | |
| Ninh Hai District, Ninh Thuan Province, Vietnam |
| 2023 |
| 39_230928_NinhThuan | |
| Ninh Hai District, Ninh Thuan Province, Vietnam |
| 2023 |
| 42_230925_NinhThuan | |
| Ninh Hai District, Ninh Thuan Province, Vietnam |
| 2023 |
| 43_230929_NinhThuan | |
| Ninh Hai District, Ninh Thuan Province, Vietnam |
| 2023 |
| 44_230929_NinhThuan | |
| Ninh Hai District, Ninh Thuan Province, Vietnam |
| 2023 |
| 45_230929_NinhThuan | |
|
| Cam Lam District, Khanh Hoa Province, Vietnam |
| 2023 |
| 54_231228_KhanhHoa |
| Huong Hoa District, Quang Tri Province, Vietnam |
| 2024 |
| 66_240702_QuangTri | |
| Huong Hoa District, Quang Tri Province, Vietnam |
| 2024 |
| 68_240702_QuangTri | |
| Huong Hoa District, Quang Tri Province, Vietnam |
| 2024 |
| 69_240702_QuangTri | |
| Huong Hoa District, Quang Tri Province, Vietnam |
| 2024 |
| 70_240702_QuangTri | |
| Chu Mon Ray National Park, Sa Thay District, Kon Tum Province, Vietnam |
| 2024 |
| 78_240704_KonTum | |
| Chu Mon Ray National Park, Sa Thay District, Kon Tum Province, Vietnam |
| 2024 |
| 79_240704_KonTum | |
| Muong Hoong commune, Dak Glei District, Kon Tum Province, Vietnam |
| 1995 | - | MNHN_P05598601 | |
| Huong Hoa District, Quang Tri Province, Vietnam |
| 2022 |
| 02_220404_QuangTri | |
| Ba Thuoc District, Thanh Hoa Province, Vietnam |
| 2022 |
| 16_220815_ThanhHoa | |
| My Duc District, Ha Noi Capital, Vietnam |
| 2023 |
| 49_231102_HaNoi | |
| My Duc District, Ha Noi Capital, Vietnam |
| 2023 |
| 50_231102_HaNoi | |
| My Duc District, Ha Noi Capital, Vietnam |
| 2023 |
| 51_231102_HaNoi | |
| Cuc Phuong National Park, Nho Quan District, Ninh Binh Province, Vietnam |
| 2004 |
| MNHN_P00819738 | |
| Sam Mo Watt village, Kan-en District, Hainan, China |
| 1934 | - | MNHN_P04783883 | |
| Suan Dtoon Falls, Muang District, Songkla Province, Thailand |
| 1985 | - | MNHN_P05474375 | |
| Surat District, Kao Wong Province, Thailand |
| 1930 | - | MNHN_P05598593 | |
|
| Cam Lo District, Quang Tri Province, Vietnam |
| 2023 |
| 38_2307_QuangTri |
| Character |
|
|
|
|
|
|
|
|---|---|---|---|---|---|---|---|
| Leaf blade | oblong, elliptic, ovate to lanceolate, 5.5–17.5 cm long | elliptic, 9–16 cm long | ovate-lanceolate, small, 1.2–7.6 cm long | ovate, ovate-elliptic to ovate-oblong, 4.5–14.5 cm long | ovate-oblong or ovate-elliptic, lanceolate, leathery, 3–10 cm long | ovate-oblong or ovate-elliptic; leathery, thick, 9–10 cm long | oblong, eliptic-oblong, or ovate-lanceolate, 7.6–21 cm long |
| Branchelet | red brown pubescent | white to grey sericeous | puberulent | - | - | pubescent | densely yellowish brown or silver-gray pubescent |
| Leaf base | lanceolate, oblong, orbicular, ovate to rotound, cuneate | cuneate | cuneate | rounded to sub-cordate | rounded | rounded | acute to rounded, cuneate, broadly cuneate or obtuse |
| Leaf apex | accumninate or retuse | acuminate | acuminate | mucronate or acuminate, rarely obtuse | rather abruptly attenuate-obtuse | obtuse rigid tip | acuminate, rarely acute or rounded |
| Leaf basal glands | 1–3 pairs | 1 pair | 1 pair | often 1 pair | 1 pair | 1 pair | often 1 pair |
| Thyrses | axillary or terminal; 2.5–20 cm long | axillary or terminal, 3–10 cm long | axillary and terminal | 4–13 cm long | axillary, 5–10 cm long | axillary and terminal, 12 cm long | axillary or terminal, 4–35 cm long |
| Sepals | ovate, apex rounded to acute, ca. 15 mm long | elliptic, apex obtuse, 4–5 mm long | orbicular, apex obtuse, ca. 2 mm long | ovate or oblong, apex rounded, ca. 3 mm long | ovate, apex rounded, ca. 2.5 mm long | apex rounded | obtuse, broadly elliptic, ovate or oblong, 2–10 mm long |
| Calyx glands | 8 glands, small, ca. 0.5 mm long | 2 glands, sometimes with additional smaller glands, ca 1 mm long | absent | usually 2 glands on each sepal, less than 1 mm long | 2–5 glands | 1–2 small glands between each sepal | only 1 gland, very large |
| Petals | white to light pink, suborbicular, erect to forward curved, 10–14 × 8–12 mm, fimbriated | pink, suborbicular, extremely reflexed, ca. 10 mm long, clawed | white, rounded plates, erect to forward curved, fimbriated | white, suborbicular, 8 × 6 mm, clawed | white, suborbicular, erect to forward curved, 15 mm long, clawed | white or pink, erect to forward curved, 10 mm long, unfimbriated | white, ovate-oblong to suborbicular, extremely reflexed, 8–15 mm long, fimbriated, clawed |
| Fruits | wingless, hemispherical | 3-winged | 3-winged | 3-winged | - | 3-winged | 3-winged |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Diversity and Evolution · Plant and animal studies · Plant and Fungal Species Descriptions
Introduction
Hiptage Gaertn. (Hiptageae, Malpighioideae, Malpighiaceae) is a genus that gathered lianas or shrubs (de Almeida et al. 2024) from, up to now, a total of 47 accepted species (POWO 2024). All species are native to Asian countries, except H.myrtifolia A.Gray which is native to Fiji. The most recent common ancestor of Hiptage probably arose in the rainforests of Indochina ca. 23 Mya and diversified in this region (de Almeida and van den Berg 2022). The most distinctive feature of the genus is probably the three-winged mericarps whose shape helps seed dispersal through wind and water. The genus is also characterised by many-flowered thyrses, which bear mirror-image flowers (Ren et al. 2013; Qian and Ren 2016). Mirror-image flowers are the result of a kind of sexual polymorphism, known as enantiostyly, where the female reproductive organs are shifted on one side, while the large stamen is shifted to the opposite side. It promotes cross-pollination between flower types and minimises self-pollination (Barrett et al. 2000; Jesson and Barrett 2002; Ren et al. 2013).
The current classification (Arènes 1954) is based on the presence and number of sepal glands that secrete oil as a reward to pollinators, with H.sect.Hiptage (one sepal nectar gland), H.sect.Archihiptage (two to ten sepal nectar glands) and H.sect.Metahiptage (no sepal gland). However, the presence and number of sepal glands are not synapomorphic characters and should, therefore, be used with caution (de Almeida and van den Berg 2022) and morphological analysis should be combined with molecular analysis to improve the reliability of classification.
Hiptagebenghalensis (L.) Kurz is the most studied Hiptage species native to India and Southeast Asia (POWO 2024). This species rapidly became invasive after its introduction in La Réunion and Mauritius (Friedmann 2011), Florida and Hawaii (Randall 2002; Starr et al. 2003) and northern Australia (Queensland) (Csurhes 2016). Invasions of H.benghalensis result in severe negative impacts on the biodiversity of the new areas and the plant is recognised by the IUCN as one of the world’s 100 worst invasive species (Lowe et al. 2000).
Vietnam is recognised as a major centre of the diversity for the Hiptage genus (Ren 2015), with 17 species recorded (Pham 2003; POWO 2024). While there are botanical descriptions from French botanists dating back to the mid-19^th^ century (e.g. Pierre 1892) and more recent studies focusing primarily on H.benghalensis (Nguyen 2003; Do et al. 2006; Chau et al. 2022), there remains a significant knowledge gap with no genetic information currently available for any specimens sampled in the country. This lack of data not only hinders the characterisation of the rich species diversity in the country, but also limits our knowledge of the genus’s history. Here, in order to obtain a better understanding of the characteristics and distributions of Hiptage species in Vietnam, we used ecological niche modelling to predict the potential genus distribution areas and support field surveys and specimen collections. Then, using a combination of morphological and molecular analyses, we determine the diversity of Hiptage species in Vietnam and retrace the distribution history of H.benghalensis. Our study also revealed a new species of Hiptage that is characterised by a wingless fruit, which has not been recorded in all the known Hiptage species (Pham 2003; POWO 2024).
Materials and methods
Hiptage niche modelling
To provide information for the field sampling, areas where plants of the genus Hiptage are likely to grow were defined using the niche modelling approaches implemented in Biomod2 (Thuiller et al. 2009). Occurrences of Hiptage individuals were obtained from GBIF on 27 June 2024 (GBIF.org 2024). It was filtered to keep individuals observed in Indo-Asia, the native area of the genus and those for which full GPS coordinates were available. The filtered dataset comprised 1,613 occurrences from 27 species. Then, using these occurrences along with climatic data from Worldclim 2 (Fick and Hijmans 2017), individual GLM, SRE, RF and MAXNET models were constructed using the SRE pseudo-absence strategy. The final ensemble model was obtained using the weighted mean method and the ROC and TSS metrics. Briefly, it implies that the better an individual model is at predicting the observed distribution, the more importance it has in the ensemble. All other parameters were kept at default or adjusted according to the Biomod2 recommendations available online (https://CRAN.R-project.org/package=biomod2). The model was projected onto Vietnam to identify the most likely locations for Hiptage.
Sample collection
Silica-dried materials were used in this study from: (i) 44 samples collected during field surveys in Vietnam from 2022 to 2024, (ii) nine samples collected in La Réunion from 2021 to 2023 and (iii) twelve herbarium specimens kept at the Muséum national d’Histoire naturelle de Paris (MNHN) which were collected between 1925 and 2004 from the following countries: China (n = 1), Laos (n = 1), Mauritius (n = 1), Seychelles (n = 3), Sri Lanka (n = 1), Thailand (n = 2) and Vietnam (n = 3). Details on all the samples are provided in Table 1.
Morphological analysis
Observations of morphological characters of the species collected in Vietnam were based on either fresh or herbarium specimens depending on their availability. For the new species, each morphological character was annotated during fieldwork and measured in detail. The macro-morphological features were studied, based on the notes in the field and micro-morphological observations were analysed and photographed using an Olympus Stereomicroscope SZX7. The morphological characters of studied specimens were compared with type specimens and protologues of the known Hiptage species (at the Herbarium Vietnam National Museum of Nature – VNMN or consulted online on Muséum national d’Histoire naturelle de Paris – MNHN online database, JSTOR Global Plants database; http://plants.jstor.org and the Chinese Virtual Herbarium; http://www.cvh.ac.cn). Taxonomic literature and morphological terminology followed Kurz (1874), Hooker (1875), Arènes (1943, 1954), Jacobs (1955), Sirirugsa (1991), Hajra et al. (1997), Pham (2003), Chen and Funston (2008), Ren et al. (2013), Ren (2015), Yang et al. (2018), Tan et al. (2019), Dong et al. (2020) and Zhang et al. (2023). The conservation status assessment of the new species was based on the International Union for Conservation of Nature guidelines (IUCN 2024).
Molecular analysis
For the samples collected during field surveys in Vietnam, total DNA was extracted from dried leaf samples using the CTAB method (Doyle and Doyle 1990). The DNA quantity and quality were measured using an Eppendorf BioPhotometer D.30 spectrophotometer (Eppendorf, Germany). Samples with DNA concentrations above 10 ng/µl and A_260 nm_/A_280 nm_ ratio in the range of 1.8 to 2.0 were used for further analyses. DNA extracts were stored at -20 °C before use. Amplicons of the nuclear ribosomal Internal Transcribed Spacer (ITS) were then obtained using the primers ITS5A (Stanford et al. 2000) and ITS4 (White et al. 1990) using either MyTaq DNA Polymerase (Bioline, Meridian Life Science) or 2X Taq Master Mix (dye plus) (Vazyme Biotech Co.). The PCR reactions were established following the manufacturer’s recommendations. PCR cycles were initial denaturation for 5 min at 95 °C, followed with 40 cycles with 95 °C for 30 sec, 58 °C for 30 sec and 72 °C for 30 sec before a final elongation for 5 min at 72 °C. In order to confirm the presence of amplicons of the expected size (~ 650 bp), PCR products were visualised using UV light after agarose gel electrophoresis and ethidium bromide staining. PCR amplicons were sent to Macrogen Company (South Korea) and Phusa Company (Vietnam) for direct sequencing in both directions. Chromatograms were then assembled using DNA Baser Sequence Assembler v.5.21.0 (Heracle BioSoft, Romania). All ambiguous positions were converted into IUPAC degenerate nucleotide codes.
For the herbarium samples, total plant DNA was extracted from 20 mg of dried leaves with DNeasy Plant Mini Kits (Qiagen). The DNA isolates were assessed by electrophoresis and their concentrations were measured by Qubit^TM^ dsDNA High Sensitivity Kits (Invitrogen). All twelve herbarium DNAs were purified with the Nucleomag NGS Clean-up and Size Select Kit (Macherey-Nagel). ITS amplification was performed using the primers ITS1P (Ridgway et al. 2003) and ITS4 (White et al. 1990) and LongAmp® Taq DNA Polymerase (New England Biolabs). Samples underwent PCR as follows: denaturation for 30 sec at 94 °C, 35 cycles with 30 sec at 94 °C, 55 sec at 50 °C, 1 min at 65 °C, then 15 minutes at 65 °C. PCR amplification was controlled by electrophoresis, then PCR amplicons were sent to Eurofins (France) for Sanger sequencing in both directions.
Phylogenetic analysis
A total of 52 ITS sequences from 21 Hiptage species and five ITS sequences of Chlorohiptage, Callaeum, Heteropterys and Niedenzuella genera, used as outgroups, were obtained from NCBI in January 2024 (Suppl. material 1). These 57 sequences, along with the 65 new sequences obtained in this study, were aligned using MAFFT v.7.520 (Katoh and Standley 2013). A phylogenetic tree was inferred with 1,000 bootstrap replicates using IQ-TREE v.2.2.6 (Minh et al. 2020). Using the automatic model selection procedure implemented in IQ-TREE, the TNe+R2 model was selected as the best-fitted following the Bayesian Information Criterion. The ML tree was visualised and annotated using the ‘plot.phylo’ function from the ‘ape’ R package (Paradis and Schliep 2019).
Results
Assessment of the Hiptage distributions in Vietnam
Whereas the Hiptage genus has a large geographical distribution, proper knowledge of the distribution of that genus in Vietnam was lacking. In order to assess the areas of Vietnam, a country that spans more than 14° of latitude, which may be most suitable for plants of Hiptage, we conducted niche modelling analyses. The analyses demonstrated that most of the Vietnamese regions (except for the Mekong Delta and some local areas) have very favourable climatic conditions for Hiptage (see the yellowish colour on the background of Fig. 1), so that a pan-country distribution of the genus could be expected. Accordingly, Hiptage samples were collected in forest ecosystems from montane and coastal areas or growing as wild or ornamental plants in urban areas from eleven provinces across Vietnam.
Vietnam map showing locations of the Hiptage samples collected during this study. Samples are coloured according to the species, as described on the legend on top-right. Probabilities of the occurrence of Hiptage in Vietnam, inferred using Biomod2 (Thuiller et al. 2023) and WorldClim 2 bioclimatic variables (Fick and Hijmans 2017), are represented on the background of the map and are coloured according to the legend on the left of the map. The yellowish areas indicate climatic conditions highly favourable for Hiptage.
As most Hiptage species only flower seasonally (Pham 2003; Chen and Funston 2008), typically in March–April each year, we could not gather all the necessary morphological data to identify the species. Indeed, out of the 44 samples collected in Vietnam during the survey, complete morphological characteristics were obtained for 14 samples. Our identification would, thus, rely on molecular analysis. In total, including samples collected during this study in Vietnam, La Réunion and the herbarium specimens, we obtained 65 ITS sequences. All the details of these samples and the accession numbers of the ITS sequences are available in Table 1.
Sequences obtained from the herbaria samples
Twelve sequences were obtained from herbaria samples labelled as H.benghalensis and deposited at MNHN (Table 1). These samples were collected from 1925 to 2004 from seven countries. Congruent with their labelling in the collection, seven of the samples were clearly clustering within the H.benghalensis clade with sequences from Mauritius, Laos, Seychelles, Sri Lanka and Vietnam. All these sequences were collected between 1925 and 2002. Interestingly, the other five sequences were grouping with other Hiptage species (Fig. 2). Amongst these, one sequence (PQ449765) from a sample collected in Vietnam in 1995 was clustering with H.marginata. Conversely, the four other sequences appeared as outgroups of currently identified species: one sequence (PQ449754) collected in Vietnam in 2004 was very close to the sequences of H.tianyangensis (MK967960) which is known endemically in China. Two sequences (PQ449764; PQ449761), obtained from Thailand and collected in 1930 and 1985, were forming a sister clade of H.multiflora and H.tianyangensis. The last sequence (PQ449758), obtained from a sample collected in China in 1934, was more closely related to three other sequences (MG730326–MG730328) also obtained from China and identified in GenBank as H.benghalensis. However, as these sequences clearly fall outside the H.benghalensis clade, they would most probably belong to another species.
Phylogenetic relationships of Hiptage, based on ITS sequences. Colours represent the origins of the sequences according to the legend shown at the top left of the tree. The presence and number of the calyx glands is indicated on the right of the tree tips according to the legend shown at the bottom left of the tree. Sequences obtained in this study are in bold and sequences from herbaria are indicated with a star (). Sequences from this study with full morphological characteristics are underlined. Open and closed circles on nodes indicate bootstrap support for the branches to their left, superior to 70% and to 90%, respectively.*
Sequences of H.benghalensis in the world
Of all the 26 H.benghalensis sequences analysed in this study, four sequence groups were identified. Whereas all the groups gathered sequences from Southeast Asia, all the sequences that were obtained from the Mascarenes and Seychelles archipelagos (reddish-coloured tips on Fig. 2), one of the main invasive areas, were grouped into a single clade. This clade gathered other sequences from samples collected in Laos and Sri Lanka, both from herbaria. These sequences were indeed 100% identical to one another. The H.benghalensis sequences from Vietnam were found in three groups: one that included additional samples from China, another that encompassed sequences from Thailand and Singapore and the third group consisting of sequences from Vietnam (including one from herbaria) alongside a sequence from India; none was associated with samples from the invasive area.
Hiptageaptera sp. nov. – a new species with wingless fruits
Sequences obtained from six samples collected in three localities in Vietnam exhibited clear genetic differences from the known Hiptage species (Fig. 2). These sequences form a distinct clade, supported by 100% of bootstrap replicates. Amongst the species and specimens analysed, H.minor shows the closest phylogenetic distance to the proposed new species and is also closely related to H.lushuiensis, H.bullata, H.subglabra, H.lucida and other unknown species. This new species exhibits diverse leaf shapes, similar to those of the comparison species. All possess basal leaf glands and several small calyx glands. However, they differ in the number of calyx glands and H.minor lacks calyx glands entirely. In addition, the sample of this new species were clearly distinguished from the related species by their unique wingless fruits and were, thus, proposed as belonging to a new species, namely Hiptageaptera M.T.X.Lam & T.V.Do, sp. nov.. The distinguishing features are presented in Table 2.
Hiptage
aptera
Taxon classificationPlantaeMalpighialesMalpighiaceae
M.T.X.Lam & T.V.Do sp. nov.
220B8B1A-C370-58DD-8FAB-C601BE32710C
urn:lsid:ipni.org:names:77362501-1
Type.
Vietnam: • Ba Ria–Vung Tau Province, Dat Do District, 10.427317, 107.254949, 39 m elev., 16 May 2022, Nguyen Hoang Nam 08_240118_VungTau (holotype: VNMN; isotype: VNMN).
Diagnosis.
This species shares similarities with H.lushuiensis, H.minor, H.bullata, H.subglabra and H.lucida such as leaf shape, basal leaf glands and petal colour. However, H.aptera differs from these related species in petal size and calyx gland presence. Its petals are significantly larger than those of H.lushuiensis, H.minor, H.bullata and H.subglabra and are erect to forward-curved with fimbriate margins, unlike the strongly reflexed petals in H.lushuiensis and the unfimbriate petals in H.lucida. Additionally, the absence of calyx glands distinguishes H.aptera from H.minor. This new species is distinctly different from H.benghalensis, one of the most studied species in the genus, which has extremely reflexed petals and a large calyx gland. A notable feature of H.aptera is its wingless, hemispherical mericarps (fruits), which are relatively large, measuring 2.4–4.5 cm in diameter. This unique fruit shape represents the first recorded occurrence within the Hiptage genus.
Description.
Woody shrubs or lianas, young branches densely red-brown pubescent, hairs appressed; older twigs glabrous, with white and small lenticels, rounded, coarse, stem wart-like structures. Leaves opposite; stipules present, linear, 1–2 mm, located at the base of the petiole, densely red-brown pubescent; petioles ca. 6 mm long, densely red-brown pubescent, adaxially canaliculate; leaf blade oblong, elliptic, ovate to lanceolate, 5.5–17.5 × 2–7 cm, coriaceous; young leaves usually red, white or red-brown pubescent on abaxial surface, densely so along mid-rib, adaxially glabrous; mature leaves green, glabrous on both surfaces, base cuneate, apex acuminate or retuse, margin entire, abaxially often with 1–3 pairs marginal gland near the base; lateral veins 5–7 pairs, prominent on both surfaces. Thyrses: terminal or axillary; main axis 2.5–20 cm long, red-brown pubescent; peduncle 2–17 mm long, covered red-brown pubescent; bracts lanceolate, 9–12 mm long, green; bracteoles lanceolate, 2–3 mm long, red-brown. Flowers: white to slightly pink; pedicels 16–22 mm long, spread red-brown pubescent. Calyx with five sepals basally connate, ovate to round, apex rounded to acute, ca. 3 × 15 mm, densely light brown pubescent abaxially, glabrous adaxially. Calyx glands: eight prominent, round, small, green, not decurrent to the pedicel, often connate at the junction between two sepals. Petals: five, suborbicular, 11–14 × 8–12 mm, white to light pink, erect to forward curved, apex rounded, margin fimbriate, claw short, ca. 0.2 mm length, ca. 0.3 mm width, basally yellow, yellow maculations, abaxially red brown pubescent, densely so near base, abaxially glabrous. Stamens ten, glabrous, unequal in size; filaments white, circinate, 9–11 mm long in the longest stamen and 2–5 mm long in the remaining nine; anthers oblong; pollen sacs yellow, rimose (longitudinal dehiscence). Ovary: ca. 25 mm, ovoid, densely light red pubescent; style 1, yellowish-green, 7.5–12 mm long, slightly curved upwards, deflected either to the left or right side, glabrous; stigma apical. Fruits: wingless, 1–3 mericarps, hemispherical, spread red brown pubescent, young fruit green, red-brown pubescent on both surfaces; mature fruit rusty to reddish-brown, glabrous, 2.4–4.5 cm in diam., adaxially with longitudinal groove, abaxially bulging. Seeds: ovoid, ca. 1.7 cm long, dark yellow or brown (Fig. 3).
Morphological characteristics of Hiptageaptera M.T.X.Lam & T.V.Do, sp. nov. with wingless fruits A, B adaxial and abaxial surface of leaf C two basal glands of leaf D flowering branch E bud of flower F flower G calyx glands H–J wingless fruits. Photos by Lam Thi Xuan Mai and Nguyen Hoang Nam, edited by Rafael Felipe de Almeida.
Phenology.
Flowering in March to June and fruiting from May to August.
Etymology.
The specific epithet reflects the unusual fruit morphology compared to other Hiptage species.
Vernacular name.
Vietnamese: Dùi đục không cánh. “Dùi đục” is the popular Vietnamese name of Hiptage genus, “không cánh” is wingless.
English name.
Wingless Hiptage.
Habitat and distribution.
Hiptageaptera is found in two Provinces: Ba Ria–Vung Tau (southern Vietnam) and Quang Tri (central Vietnam). It grows in areas around the base of mountains, on slopes and along mountain paths under evergreen forests and near residential areas, at elevations of 28–433 m.
Conservation status
(and study). Since its identification, we have found this species in various locations within Ba Ria–Vung Tau Province, where it maintains a relatively large population. It is commonly observed as shrubs or lianas covering certain areas, co-existing with other species in the same habitat. We propose that the new species H.aptera should be classified as Least Concern (LC) according to the current IUCN Red List Categories and Criteria (IUCN 2024). However, continuous monitoring of its population status and habitat conditions is crucial for making accurate assessments in the future.
Additional specimens examined
(paratypes). Vietnam: • Dat Do, Ba Ria–Vung Tau, 10.4064358, 107.2631240, 194 m elev., 8 Apr 2024, Lam Thi Xuan Mai, original ID 61_240408_VungTau; • Dat Do, Ba Ria–Vung Tau, 10.406327, 107.263000, 190 m elev., 01 Jun 2024, Lam Thi Xuan Mai, original ID 62_240601_VungTau; • Tan Thanh, Ba Ria–Vung Tau, 10.545323, 107.147117, 333 m elev., 23 Jun 2024, Nguyen Hoang Nam, original ID 64_240623_VungTau; • Dat Do, Ba Ria–Vung Tau, 10.433306, 107.254722, 28 m elev., 23 Jun 2024, Nguyen Hoang Nam, original ID 65_240623_Vung Tau; • Da Krong, Quang Tri, 16.328694, 106.997167, 433 m elev., 01 Dec 2022, Tran Dang Khoa, original ID 19_221201_QuangTri.
Discussion
Presence, diversity and distribution of Hiptage genus in Vietnam
This study represents the first attempt at molecular characterisation of Hiptage species in Vietnam. From our results, four species that have already been known elsewhere and a new species were identified, these Hiptage populations being distributed across various localities in Vietnam. Besides, our study also included six unidentified samples. Based on the molecular analysis and despite the absence of morphological description, these samples (sequences PQ449708; PQ449717; PQ449734–PQ449736 and PQ449754; Hiptage spp.) were most likely belonging to two unidentified species. It is important to note that Vietnam is home to 17 Hiptage species, twelve of which are endemic to the country (POWO 2024). This study updated the count to 18 species, with 13 species currently known exclusively from Vietnam. Whereas our analyses revealed that most of Vietnam presents climatic conditions favourable to plants from the Hiptage genus in general, it would now be important from a botanical and conservation perspective to gather more knowledge on these 18 species and their distinct distribution in the country. Resampling during the flowering and fruiting season will be essential for further characterisation. Confronting the morphological reports of these species would reveal how the newly-uncovered clades match or complement the known diversity of Hiptage in Vietnam. The presence of known species, a new species and unidentified specimens further supports the notion that the Indochina Peninsula is a hotspot for the genus and Vietnam is a centre of its diversity (Ren 2015; de Almeida and van den Berg 2022).
Potential migration route to the South West Indian Ocean Islands
Based on the phylogenetic relatedness of the 13 H.benghalensis samples collected in the secondary area, it is interesting to notice first that all the samples fall in the same clade, suggestive of a single introduction event of the plant into the region or all the introductions in all the islands were from the same area. Indeed, based on historical reports (Céré 1785; M.-S. Jagou and F. Martos, pers. comm.), it is thought that the liana was introduced in Mauritius in the late 18^th^ century and later spread to the surrounding islands. Nevertheless, although our analysis did support that hypothesis, it remains difficult to retrace the introduction route to any of the native areas. One sample from Laos and another from Sri Lanka are, indeed, genetically similar to the introduced ones. However, no clear-cut conclusion could be drawn, as determining the origin of any invasive populations often requires more abundant and more intensive sampling from their native range (Hagenblad et al. 2015) and more diverse genetic markers would certainly be required. In addition, even though all the samples collected in Vietnam appeared in other phylogenetic groups, it does not absolutely rule out the possibility that the introduced populations were from that area. It is very likely that other populations with different genetic profiles are present in Vietnam, but were not sampled here. It is also entirely possible that some populations existed in Vietnam at some point, but are now extinct.
Collecting additional samples in Vietnam and from other Southeast Asian countries and the Indian Peninsula would help clarify the migration routes of H.benghalensis to the South West Indian Ocean Islands. Our study also points to the requirement for a more discriminant genetic method that would help refine our understanding of sample relationships. Such an initiative would provide invaluable clues on how H.benghalensis may have disseminated to the world, where it has become invasive and caused ecological consequences in the introduced areas. It is noteworthy that, in the process of achieving this feat, populations of previously unknown Hiptage species were discovered, putting this genus at both extremes of the conservation spectrum.
Wingless fruits of Hiptageaptera – A morphological innovation in Hiptage
The newly-identified species from Vietnam, named Hiptageaptera, derives its name from the Greek term “aptera,” which refers to the absence of wings on the fruit. This species presents eight calyx glands, a clear distinctive character from the closest related species in the ITS phylogeny, H.minor with no calyx gland or other closely-related species, H.lushuiensis, H.bullata, H.subglabra and H.lucida, which differ in the number of calyx glands present. It must be noted, however, that this character is homoplastic in the Hiptage genus and can be problematic for taxonomic classification (de Almeida and Van den Berg 2022). Nevertheless, the phylogenetic tree provides strong support for distinguishing this species. The species produces large, hemispherical, wingless fruits known as smooth mericarps. Whereas it represents a first for the Hiptage genus, the observation of a mixed winged- and wingless-fruit was also observed in other genera within the family, such as Camarea, Mascagnia and several other Neotropical genera (Davis and Anderson 2010). Understanding the factors associated with this change would be important. The realistic observation that the fruit was floating on the water points to a possible dispersal through water (hydrochory), which is a very common strategy in Amazonian Malpighiaceae growing near large bodies of water (de Almeida et al. 2024). The transition from winged to wingless fruits with hard pericarps may also be related to a “defence scenario” proposed by Mack (2000). This change enhances the protective ability to defend seeds and secondarily evolves into structures that promote seed dispersal.
Conclusion
Our study highlights Vietnam as a significant centre of diversity for the Hiptage genus, with 18 species to date, including both previously-recorded species and the newly-described Hiptageaptera. The new species is distinct with its unique morphological trait – wingless fruit, contrary to all known Hiptage species. Furthermore, while the origin of H.benghalensis in the Mascarene Islands remains unclear, our genetic analysis indicated a probable single introduction event, aligning with historical records from the 18^th^ century.
Supplementary Material
XML Treatment for Hiptage aptera
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arènes J (1943) Contribution à l’étude des Malpighiacées.Notulae Systematicae 11(2): 62–80. https://www.biodiversitylibrary.org/part/324501
- 2Arènes J (1954) Florae Malesianae precusores-VI. Malpighiaceae malesianae novae.Reinwardtia 3(1): 67–74. 10.55981/reinwardtia.1954.960 · doi ↗
- 3Barrett SCH Jesson LK Baker AM (2000) The evolution and function of stylar polymorphisms in flowering plants.Annals of Botany 85: 253–265. 10.1006/anbo.1999.1067 · doi ↗
- 4CéréN (1785) Recensement de tout ce que renferme le Jardin du Roi, le Monplaisir, Isle de France [Census of everything contained in the Jardin du Roi, Monplaisir, French Island]. National Archives, A.N. Col C/4/66, f°92–103 and MNHN Archives. Ms: 303.
- 5Chau DV Nam PV Chung NT (2022) Đa dạng nguồn gen cây thuốc của dân tộc mường, thu thập lưu giữ và bảo tồn tại tỉnh thanh hóa năm 2021 [Diversity of genetic resources of medicine traditional medicine collected, stored and conserved in Thanh Hoa province in 2021]. Tạp chí Khoa học Trường Đại học Hồng Đức (62): 17–26. https://hdujs.edu.vn/index.php/hdujs/article/view/2/2
- 6Chen S Funston AM (2008) Malpighiaceae. In: Wu ZY Raven PH Hong DY (Eds) Flora of China 11 (Oxalidaceae through Aceraceae).Science Press, Beijing & Missouri Botanical Garden Press, 135–136.
- 7Csurhes S (2016) Invasive plant risk assessment: Hiptagebenghalensis. The State of Queensland, Department of Employment, Economic Development and Innovation, 1–11. https://www.daf.qld.gov.au/__data/assets/pdf_file/0006/69540/IPA-Hiptage-Risk-Assessment.pdf
- 8Davis CC Anderson WR (2010) A complete generic phylogeny of Malpighiaceae inferred from nucleotide sequence data and morphology.American Journal of Botany 97(12): 2031–2048. 10.3732/ajb.100014621616850 · doi ↗ · pubmed ↗