The genome sequence of a small dung beetle, Volinus sticticus (Panzer, 1798), formerly known as Aphodius sticticus
František Sládeček, Liam M. Crowley, Doga Cedden, Lindsey C Perkin, Hume Douglas

TL;DR
This paper provides the genome sequence of a small dung beetle, including its chromosomes and mitochondrial DNA.
Contribution
The study presents a high-quality genome assembly of the dung beetle Aphodius sticticus, including sex chromosomes and mitochondrial DNA.
Findings
The genome assembly is 628.40 megabases in total length.
98.96% of the assembly is scaffolded into 11 chromosomal pseudomolecules.
The mitochondrial genome is 20.8 kilobases in length.
Abstract
We present a genome assembly from an individual male specimen of the small dung beetle, Aphodius sticticus (Arthropoda; Insecta; Coleoptera; Scarabaeidae). The genome sequence has a total length of 628.40 megabases. Most of the assembly (98.96%) is scaffolded into 11 chromosomal pseudomolecules, including the X and Y sex chromosomes. The mitochondrial genome has also been assembled and is 20.8 kilobases in length.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1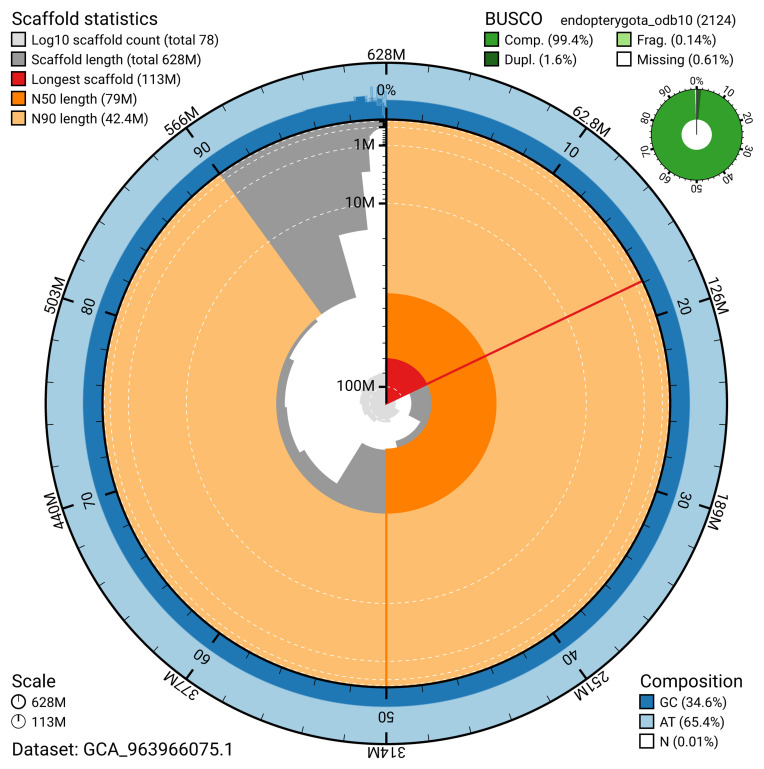 Figure 2
Figure 2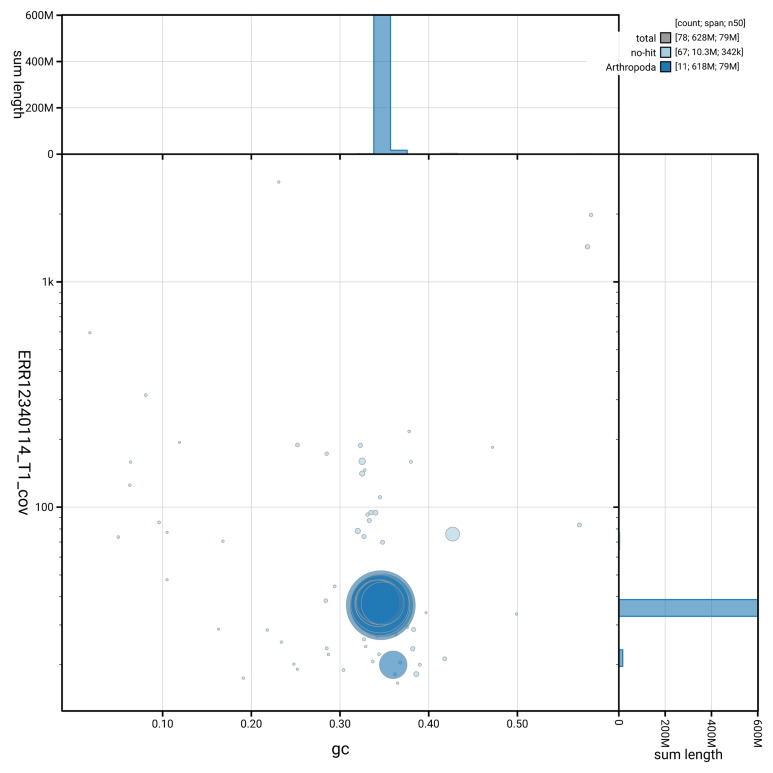 Figure 3
Figure 3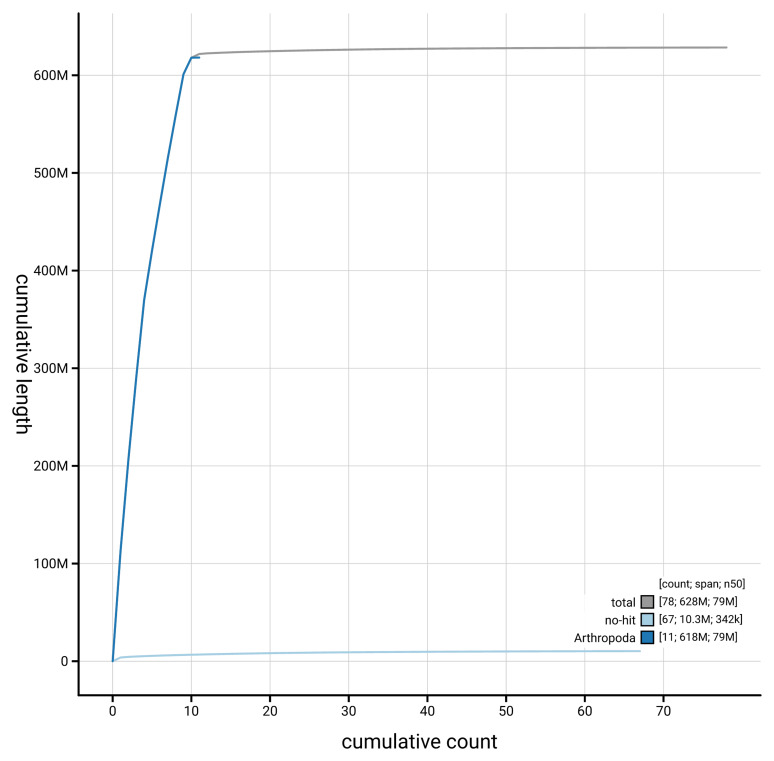 Figure 4
Figure 4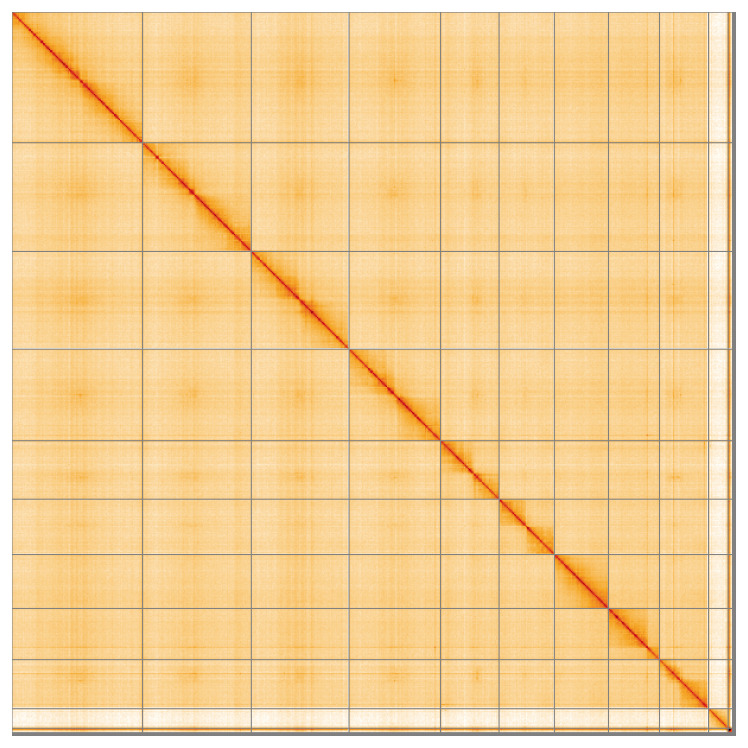 Figure 5
Figure 5| Project information | |||
|---|---|---|---|
|
| Aphodius sticticus | ||
|
| PRJEB70743 | ||
|
|
| ||
|
| SAMEA112774809 | ||
|
| 207213 | ||
| Specimen information | |||
|
|
|
|
|
|
| icAphStic4 | SAMEA112774903 | Whole organism |
|
| icAphStic2 | SAMEA112774901 | Whole organism |
| Sequencing information | |||
|
|
|
|
|
|
| ERR12342492 | 7.96e+08 | 120.19 |
|
| ERR12340114 | 2.16e+06 | 22.88 |
| Genome assembly | ||
|---|---|---|
| Assembly name | icAphStic4.1 | |
| Assembly accession | GCA_963966075.1 | |
|
|
| |
| Span (Mb) | 628.40 | |
| Number of contigs | 361 | |
| Number of scaffolds | 77 | |
| Longest scaffold (Mb) | 112.87 | |
| Assembly metrics
|
| |
| Contig N50 length (Mb) | 4.8 |
|
| Scaffold N50 length (Mb) | 79.0 |
|
| Consensus quality (QV) | Primary: 60.6; alternate: 59.6;
|
|
|
| Primary: 80.97%; alternate:
|
|
| BUSCO
| C:99.3%[S:97.7%,D:1.6%],
|
|
| Percentage of assembly
| 98.96% |
|
| Sex chromosomes | XY |
|
| Organelles | Mitochondrial genome: 20.8 kb |
|
| INSDC accession | Name | Length (Mb) | GC% |
|---|---|---|---|
| 1 | 112.75 | 34.5 | |
| 2 | 93.81 | 34.5 | |
| 3 | 84.26 | 34.5 | |
| 4 | 79.04 | 34.5 | |
| 5 | 50.49 | 34.5 | |
| 6 | 47.68 | 34.5 | |
| 7 | 46.68 | 34.5 | |
| 8 | 44.06 | 34.5 | |
| 9 | 42.37 | 35.0 | |
| X | 16.86 | 36.0 | |
| Y | 3.8 | 42.5 | |
| MT | 0.02 | 23.0 |
| Software tool | Version | Source |
|---|---|---|
| BEDTools | 2.30.0 |
|
| BLAST | 2.14.0 |
|
| BlobToolKit | 4.3.7 |
|
| BUSCO | 5.4.3 and 5.5.0 |
|
| bwa-mem2 | 2.2.1 |
|
| Cooler | 0.8.11 |
|
| DIAMOND | 2.1.8 |
|
| fasta_windows | 0.2.4 |
|
| FastK | 427104ea91c78c3b8b8b49f1a7d6bbeaa869ba1c |
|
| Gfastats | 1.3.6 |
|
| GoaT CLI | 0.2.5 |
|
| Hifiasm | 0.19.8-r587 |
|
| HiGlass | 44086069ee7d4d3f6f3f0012569789ec138f42b84
|
|
| Merqury.FK | d00d98157618f4e8d1a9190026b19b471055b
|
|
| MitoHiFi | 3 |
|
| MultiQC | 1.14, 1.17, and 1.18 |
|
| NCBI Datasets | 15.12.0 |
|
| Nextflow | 23.04.0-5857 |
|
| PretextView | 0.2.5 |
|
| purge_dups | 1.2.5 |
|
| samtools | 1.16.1, 1.17, and 1.18 |
|
| sanger-tol/ascc | - |
|
| sanger-tol/blobtoolkit | 0.6.0 |
|
| Seqtk | 1.3 |
|
| Singularity | 3.9.0 |
|
| TreeVal | 1.0.0 |
|
| YaHS | 1.2a.2 |
|
- —Wellcome Trust
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect Resistance and Genetics · Coleoptera Taxonomy and Distribution · Insect symbiosis and bacterial influences
Species taxonomy
Eukaryota; Opisthokonta; Metazoa; Eumetazoa; Bilateria; Protostomia; Ecdysozoa; Panarthropoda; Arthropoda; Mandibulata; Pancrustacea; Hexapoda; Insecta; Dicondylia; Pterygota; Neoptera; Endopterygota; Coleoptera; Polyphaga; Scarabaeiformia; Scarabaeoidea; Scarabaeidae; Aphodiinae; Aphodius; Volinus; Aphodius sticticus (Panzer, 1798) (NCBI:txid207213)
Background
The subfamily Aphodiinae is a diverse group of beetles within the scarab beetle family, Scarabaeidae, and has a global distribution. While many aphodiines are dung beetles, some species are associated with decaying vegetable matter or burrows of other beetles, such as Geotrupidae ( British Scarabs, 2025). In Britain and Ireland, 46 aphodiine species have been recorded ( Duff, 2018). Different species of Aphodiinae commonly coexist within dung piles by occupying distinct ecological niches, including preferences for dung type, age, soil conditions, or exposure to sunlight, which helps to minimise competition among species ( British Scarabs, 2025). Recent taxonomic revisions have reclassified many former members of the genus Aphodius into separate genera, reflecting advances in our understanding of their evolutionary relationships ( British Scarabs, 2025). The species formerly known as Aphodius sticticus is now placed within the genus Volinus.
Volinus sticticus is a small dung beetle, measuring approximately 4.5–5.0 mm in length, with distinctive black streaks on yellow-brown elytra. It is considered a “dweller” spending its entire life cycle within dung ( British Scarabs, 2025). It shows a preference for horse and sheep dung in shaded habitats.
Volinus sticticus is distributed across parts of the Palaearctic region, with records primarily from Europe. Most occurrences are concentrated in western and central Europe, with additional records in countries such as Turkey and Albania ( GBIF Secretariat, 2023). It is fairly common in England and Wales and can occur in numbers where it is found ( NBN Atlas Partnership, 2024).
We present a chromosome-level genome sequence for Aphodius sticticus, based on a male specimen from Blackdown Farm, Gloucestershire, United Kingdom ( Figure 1).
Photograph of the Aphodius sticticus (icAphStic4) specimen used for genome sequencing.
Genome sequence report
The genome of Aphodius sticticus ( Figure 1) was sequenced using Pacific Biosciences single-molecule HiFi long reads, generating a total of 22.88 Gb (gigabases) from 2.16 million reads, providing an estimated 36-fold coverage. Chromosome conformation Hi-C sequencing produced 120.19 Gb from 795.96 million reads. Specimen and sequencing details are summarised in Table 1.
Table 1.: Specimen and sequencing data for Aphodius sticticus.
Assembly errors were corrected by manual curation, including 54 missing joins or mis-joins and 5 haplotypic duplications. This reduced the scaffold number by 9.3%. The final assembly has a total length of 628.40 Mb in 77 sequence scaffolds, with 283 gaps, and a scaffold N50 of 79.0 Mb ( Table 2).
Table 2.: Genome assembly data for Aphodius sticticus, icAphStic4.1.
The snail plot in Figure 2 provides a summary of the assembly statistics, indicating the distribution of scaffold lengths and other assembly metrics. Figure 3 shows the distribution of scaffolds by GC proportion and coverage. Figure 4 presents a cumulative assembly plot, with separate curves representing different scaffold subsets assigned to various phyla, illustrating the completeness of the assembly.
Genome assembly of Aphodius sticticus, icAphStic4.1: metrics.The BlobToolKit snail plot provides an overview of assembly metrics and BUSCO gene completeness. The circumference represents the length of the whole genome sequence, and the main plot is divided into 1,000 bins around the circumference. The outermost blue tracks display the distribution of GC, AT, and N percentages across the bins. Scaffolds are arranged clockwise from longest to shortest and are depicted in dark grey. The longest scaffold is indicated by the red arc, and the deeper orange and pale orange arcs represent the N50 and N90 lengths. A light grey spiral at the centre shows the cumulative scaffold count on a logarithmic scale. A summary of complete, fragmented, duplicated, and missing BUSCO genes in the endopterygota_odb10 set is presented at the top right. An interactive version of this figure is available at https://blobtoolkit.genomehubs.org/view/GCA_963966075.1/dataset/GCA_963966075.1/snail.
Genome assembly of Aphodius sticticus, icAphStic4.1: BlobToolKit GC-coverage plot showing sequence coverage (vertical axis) and GC content (horizontal axis).The circles represent scaffolds, with the size proportional to scaffold length and the colour representing phylum membership. The histograms along the axes display the total length of sequences distributed across different levels of coverage and GC content. An interactive version of this figure is available at https://blobtoolkit.genomehubs.org/view/GCA_963966075.1/dataset/GCA_963966075.1/blob.
Genome assembly of Aphodius sticticus icAphStic4.1: BlobToolKit cumulative sequence plot.The grey line shows cumulative length for all scaffolds. Coloured lines show cumulative lengths of scaffolds assigned to each phylum using the buscogenes taxrule. An interactive version of this figure is available at https://blobtoolkit.genomehubs.org/view/GCA_963966075.1/dataset/GCA_963966075.1/cumulative.
Most of the assembly sequence (98.96%) was assigned to 11 chromosomal-level scaffolds, representing 9 autosomes and the X and Y sex chromosomes. These chromosome-level scaffolds, confirmed by the Hi-C data, are named in order of size ( Figure 5; Table 3). During manual curation, chromosomes X and Y were assigned based on read coverage statistics.
Genome assembly of Aphodius sticticus icAphStic4.1: Hi-C contact map of the icAphStic4.1 assembly, visualised using HiGlass.Chromosomes are shown in order of size from left to right and top to bottom. An interactive version of this figure may be viewed at https://genome-note-higlass.tol.sanger.ac.uk/l/?d=QScANppxQV2p1Yw8VLrZvA.
Table 3.: Chromosomal pseudomolecules in the genome assembly of Aphodius sticticus, icAphStic4.
While not fully phased, the assembly deposited is of one haplotype. Contigs corresponding to an alternate haplotype have also been deposited. The mitochondrial genome was also assembled and can be found as a contig within the multifasta file of the genome submission, and as a separate fasta file.
Assembly quality metrics
The estimated Quality Value (QV) and k-mer completeness metrics, along with BUSCO completeness scores, were calculated for each haplotype and the combined assembly. The QV reflects the base-level accuracy of the assembly, while k-mer completeness indicates the proportion of expected k-mers identified in the assembly. BUSCO scores provide a measure of completeness based on benchmarking universal single-copy orthologues.
The primary haplotype has a QV of 60.6, and the combined primary and alternate assemblies achieve an estimated QV of 60.1. The k-mer completeness for the primary haplotype is 80.97%, and for the alternate haplotype it is 77.35%, while the combined primary and alternate assemblies achieve a k-mer completeness of 99.07%. BUSCO analysis using the endopterygota_odb10 reference set ( n = 2,124) indicated a completeness score of 99.4% (single = 97.8%, duplicated = 1.6%).
Table 2 provides assembly metric benchmarks adapted from Rhie et al. (2021) and the Earth BioGenome Project Report on Assembly Standards September 2024. The assembly achieves the EBP reference standard of 6.C.Q60.
Methods
Sample acquisition and DNA barcoding
Specimens of Aphodius sticticus were collected from Blackdown Farm, Gloucestershire, United Kingdom (latitude 52.04, longitude –1.70) on 2022-08-25 by potting. The specimens were collected by František Sládeček (University of South Bohemia) and Liam Crowley (University of Oxford), identified by František Sládeček, and then preserved on dry ice. The specimen used for PacBio HiFi sequencing was specimen ID Ox002960 (ToLID icAphStic4), while another specimen (specimen ID Ox002958, ToLID icAphStic2) was used for Hi-C scaffolding of the assembly.
The initial identification was verified by an additional DNA barcoding process according to the framework developed by Twyford et al. (2024). A small sample was dissected from the specimens and stored in ethanol, while the remaining parts were shipped on dry ice to the Wellcome Sanger Institute (WSI). The tissue was lysed, the COI marker region was amplified by PCR, and amplicons were sequenced and compared to the BOLD database, confirming the species identification ( Crowley et al., 2023). Following whole genome sequence generation, the relevant DNA barcode region was also used alongside the initial barcoding data for sample tracking at the WSI ( Twyford et al., 2024). The standard operating procedures for Darwin Tree of Life barcoding have been deposited on protocols.io ( Beasley et al., 2023).
Nucleic acid extraction
The workflow for high molecular weight (HMW) DNA extraction at the WSI Tree of Life Core Laboratory includes a sequence of procedures: sample preparation and homogenisation, DNA extraction, fragmentation and purification. Detailed protocols are available on protocols.io ( Denton et al., 2023b). The icAphStic4 sample was prepared for DNA extraction by weighing and dissecting it on dry ice ( Jay et al., 2023). Tissue from the whole organism was homogenised using a PowerMasher II tissue disruptor ( Denton et al., 2023a). HMW DNA was extracted using the Automated MagAttract v2 protocol ( Oatley et al., 2023a). DNA was sheared into an average fragment size of 12–20 kb in a Megaruptor 3 system ( Bates et al., 2023). Sheared DNA was purified by solid-phase reversible immobilisation, using AMPure PB beads to eliminate shorter fragments and concentrate the DNA ( Oatley et al., 2023b). The concentration of the sheared and purified DNA was assessed using a Nanodrop spectrophotometer and Qubit Fluorometer using the Qubit dsDNA High Sensitivity Assay kit. Fragment size distribution was evaluated by running the sample on the FemtoPulse system.
Hi-C sample preparation
Tissue from the whole organism of the icAphStic2 sample was processed at the WSI Scientific Operations core, using the Arima-HiC v2 kit. Tissue (stored at –80 °C) was fixed, and the DNA crosslinked using a TC buffer with 22% formaldehyde, final concentration 2%. After crosslinking, the tissue was homogenised using the Diagnocine Power Masher-II and BioMasher-II tubes and pestles. Following the kit manufacturer's instructions, crosslinked DNA was digested using a restriction enzyme master mix. The 5’-overhangs were then filled in and labelled with biotinylated nucleotides and proximally ligated. An overnight incubation was carried out for enzymes to digest remaining proteins and for crosslinks to reverse. A clean up was performed with SPRIselect beads prior to library preparation.
** PacBio HiFi **
At a minimum, samples were required to have an average fragment size exceeding 8 kb and a total mass over 400 ng to proceed to the low input SMRTbell Prep Kit 3.0 protocol (Pacific Biosciences, California, USA), depending on genome size and sequencing depth required. Libraries were prepared using the SMRTbell Prep Kit 3.0 (Pacific Biosciences, California, USA) as per the manufacturer's instructions. The kit includes the reagents required for end repair/A-tailing, adapter ligation, post-ligation SMRTbell bead cleanup, and nuclease treatment. Following the manufacturer’s instructions, size selection and clean up was carried out using diluted AMPure PB beads (Pacific Biosciences, California, USA). DNA concentration was quantified using the Qubit Fluorometer v4.0 (Thermo Fisher Scientific) with Qubit 1X dsDNA HS assay kit and the final library fragment size analysis was carried out using the Agilent Femto Pulse Automated Pulsed Field CE Instrument (Agilent Technologies) and gDNA 55kb BAC analysis kit.
Samples were sequenced on a Revio instrument (Pacific Biosciences, California, USA). Prepared libraries were normalised to 2 nM, and 15 μL was used for making complexes. Primers were annealed and polymerases were hybridised to create circularised complexes according to manufacturer’s instructions. The complexes were purified with the 1.2X clean up with SMRTbell beads. The purified complexes were then diluted to the Revio loading concentration (in the range 200–300 pM), and spiked with a Revio sequencing internal control. Samples were sequenced on Revio 25M SMRT cells (Pacific Biosciences, California, USA). The SMRT link software, a PacBio web-based end-to-end workflow manager, was used to set-up and monitor the run, as well as perform primary and secondary analysis of the data upon completion.
** Hi-C **
For Hi-C library preparation, DNA was fragmented using the Covaris E220 sonicator (Covaris) and size selected using SPRISelect beads to 400 to 600 bp. The DNA was then enriched using the Arima-HiC v2 kit Enrichment beads. Using the NEBNext Ultra II DNA Library Prep Kit (New England Biolabs) for end repair, A-tailing, and adapter ligation. This uses a custom protocol which resembles the standard NEBNext Ultra II DNA Library Prep protocol but where library preparation occurs while DNA is bound to the Enrichment beads. For library amplification, 10 to 16 PCR cycles were required, determined by the sample biotinylation percentage. The Hi-C sequencing was performed using paired-end sequencing with a read length of 150 bp on an Illumina NovaSeq 6000 instrument.
Genome assembly, curation and evaluation
** Assembly **
The HiFi reads were assembled using Hifiasm ( Cheng et al., 2021) with the --primary option. Haplotypic duplications were identified and removed using purge_dups ( Guan et al., 2020). The Hi-C reads were mapped to the primary contigs using bwa-mem2 ( Vasimuddin et al., 2019). The contigs were further scaffolded using the provided Hi-C data ( Rao et al., 2014) in YaHS ( Zhou et al., 2023) using the --break option for handling potential misassemblies. The scaffolded assemblies were evaluated using Gfastats ( Formenti et al., 2022), BUSCO ( Manni et al., 2021) and MERQURY.FK ( Rhie et al., 2020).
The mitochondrial genome was assembled using MitoHiFi ( Uliano-Silva et al., 2023), which runs MitoFinder ( Allio et al., 2020) and uses these annotations to select the final mitochondrial contig and to ensure the general quality of the sequence.
** Assembly curation **
The assembly was decontaminated using the Assembly Screen for Cobionts and Contaminants (ASCC) pipeline (article in preparation). Flat files and maps used in curation were generated in TreeVal ( Pointon et al., 2023). Manual curation was primarily conducted using PretextView ( Harry, 2022), with additional insights provided by JBrowse2 ( Diesh et al., 2023) and HiGlass ( Kerpedjiev et al., 2018). Scaffolds were visually inspected and corrected as described by Howe et al. (2021). Any identified contamination, missed joins, and mis-joins were corrected, and duplicate sequences were tagged and removed. The curation process is documented at https://gitlab.com/wtsi-grit/rapid-curation (article in preparation).
** Assembly quality assessment **
The Merqury.FK tool ( Rhie et al., 2020), run in a Singularity container ( Kurtzer et al., 2017), was used to evaluate k-mer completeness and assembly quality for the primary and alternate haplotypes using the k-mer databases ( k = 31) that were computed prior to genome assembly. The analysis outputs included assembly QV scores and completeness statistics.
A Hi-C contact map was produced for the final version of the assembly. Hi-C reads were aligned using bwa-mem2 ( Vasimuddin et al., 2019) and the alignment files were combined using SAMtools ( Danecek et al., 2021). The Hi-C alignments were converted into a contact map using BEDTools ( Quinlan & Hall, 2010) and the Cooler tool suite ( Abdennur & Mirny, 2020). The contact map was visualised in HiGlass ( Kerpedjiev et al., 2018).
The blobtoolkit pipeline is a Nextflow port of the previous Snakemake Blobtoolkit pipeline ( Challis et al., 2020). It aligns the PacBio reads in SAMtools and minimap2 ( Li, 2018) and generates coverage tracks for regions of fixed size. In parallel, it queries the GoaT database ( Challis et al., 2023) to identify all matching BUSCO lineages to run BUSCO ( Manni et al., 2021). For the three domain-level BUSCO lineages, the pipeline aligns the BUSCO genes to the UniProt Reference Proteomes database ( Bateman et al., 2023) with DIAMOND blastp ( Buchfink et al., 2021). The genome is also divided into chunks according to the density of the BUSCO genes from the closest taxonomic lineage, and each chunk is aligned to the UniProt Reference Proteomes database using DIAMOND blastx. Genome sequences without a hit are chunked using seqtk and aligned to the NT database with blastn ( Altschul et al., 1990). The blobtools suite combines all these outputs into a blobdir for visualisation.
The blobtoolkit pipeline was developed using nf-core tooling ( Ewels et al., 2020) and MultiQC ( Ewels et al., 2016), relying on the Conda package manager, the Bioconda initiative ( Grüning et al., 2018), the Biocontainers infrastructure ( da Veiga Leprevost et al., 2017), as well as the Docker ( Merkel, 2014) and Singularity ( Kurtzer et al., 2017) containerisation solutions.
Table 4 contains a list of relevant software tool versions and sources.
Wellcome Sanger Institute – Legal and Governance
The materials that have contributed to this genome note have been supplied by a Darwin Tree of Life Partner. The submission of materials by a Darwin Tree of Life Partner is subject to the ‘Darwin Tree of Life Project Sampling Code of Practice’, which can be found in full on the Darwin Tree of Life website here. By agreeing with and signing up to the Sampling Code of Practice, the Darwin Tree of Life Partner agrees they will meet the legal and ethical requirements and standards set out within this document in respect of all samples acquired for, and supplied to, the Darwin Tree of Life Project.
Further, the Wellcome Sanger Institute employs a process whereby due diligence is carried out proportionate to the nature of the materials themselves, and the circumstances under which they have been/are to be collected and provided for use. The purpose of this is to address and mitigate any potential legal and/or ethical implications of receipt and use of the materials as part of the research project, and to ensure that in doing so we align with best practice wherever possible. The overarching areas of consideration are:
• Ethical review of provenance and sourcing of the material
• Legality of collection, transfer and use (national and international)
Each transfer of samples is further undertaken according to a Research Collaboration Agreement or Material Transfer Agreement entered into by the Darwin Tree of Life Partner, Genome Research Limited (operating as the Wellcome Sanger Institute), and in some circumstances other Darwin Tree of Life collaborators.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abdennur N Mirny LA : Cooler: scalable storage for Hi-C data and other genomically labeled arrays. Bioinformatics. 2020;36(1):311–316. 10.1093/bioinformatics/btz 540 31290943 PMC 8205516 · doi ↗ · pubmed ↗
- 2Allio R Schomaker-Bastos A Romiguier J : Mito Finder: efficient automated large-scale extraction of mitogenomic data in target enrichment phylogenomics. Mol Ecol Resour. 2020;20(4):892–905. 10.1111/1755-0998.13160 32243090 PMC 7497042 · doi ↗ · pubmed ↗
- 3Altschul SF Gish W Miller W : Basic local alignment search tool. J Mol Biol. 1990;215(3):403–410. 10.1016/S 0022-2836(05)80360-2 2231712 · doi ↗ · pubmed ↗
- 4Bateman A Martin MJ Orchard S : Uni Prot: the universal protein knowledgebase in 2023. Nucleic Acids Res. 2023;51(D 1):D 523–D 531. 10.1093/nar/gkac 1052 36408920 PMC 9825514 · doi ↗ · pubmed ↗
- 5Bates A Clayton-Lucey I Howard C : Sanger Tree of Life HMW DNA fragmentation: diagenode Megaruptor ®3 for LI Pac Bio. protocols.io. 2023. 10.17504/protocols.io.81wgbxzq 3lpk/v 1 · doi ↗
- 6Beasley J Uhl R Forrest LL : DNA barcoding SO Ps for the Darwin Tree of Life project. protocols.io. 2023; [Accessed 25 June 2024]. 10.17504/protocols.io.261ged 91jv 47/v 1 · doi ↗
- 7British Scarabs: Dwellers – Aphodiines.2025; [Accessed 28 January 2025]. Reference Source
- 8Buchfink B Reuter K Drost HG : Sensitive protein alignments at Tree-of-Life scale using DIAMOND. Nat Methods. 2021;18(4):366–368. 10.1038/s 41592-021-01101-x 33828273 PMC 8026399 · doi ↗ · pubmed ↗