NtANTL2 overexpression regulates starch-related and nitrogen metabolism in tobacco plants
Bo Lei, Fei Wang, Huina Zhao, Benbo Xu, Le Xu, Lingli Xie

TL;DR
Overexpression of the NtANTL2 gene in tobacco plants improves nitrogen tolerance and affects carbon and nitrogen metabolism.
Contribution
This study reveals the regulatory role of NtANTL2 in balancing carbon and nitrogen metabolism through enzyme activity and gene expression changes.
Findings
NtANTL2 overexpression increases tolerance to high nitrogen and enhances growth in tobacco plants.
Overexpression alters key enzyme activities and gene expressions related to starch and nitrogen metabolism.
Increased amino acid levels and enzyme activities like GDH demonstrate NtANTL2's regulatory role.
Abstract
The function of the aromatic and neutral amino acid transporter gene ANT1-like (NtANTL2) was investigated. NtANTL2-3 over-expressor plants displayed greater tolerance to high nitrogen application compared to WT plants, characterized by better growth. As for carbon metabolism, the activity of sucrose phosphate synthetase in overexpression lines was 1.5 times higher than that in wild-type plants at 0.1% nitrogen, while sucrose synthase activity was significantly lower under all nitrogen levels. These changes in enzyme activities were closely associated with the differential gene expressions of GBSSI (granule—bound starcsssh synthase I), SUS (sucrose synthase), INV (invertase), and SPS (sucrose phosphate synthase). The activities and gene expressions of key enzymes in nitrogen metabolism like nitrate reductase, glutamine synthetase, glutamate synthase, and glutamate dehydrogenase were also…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1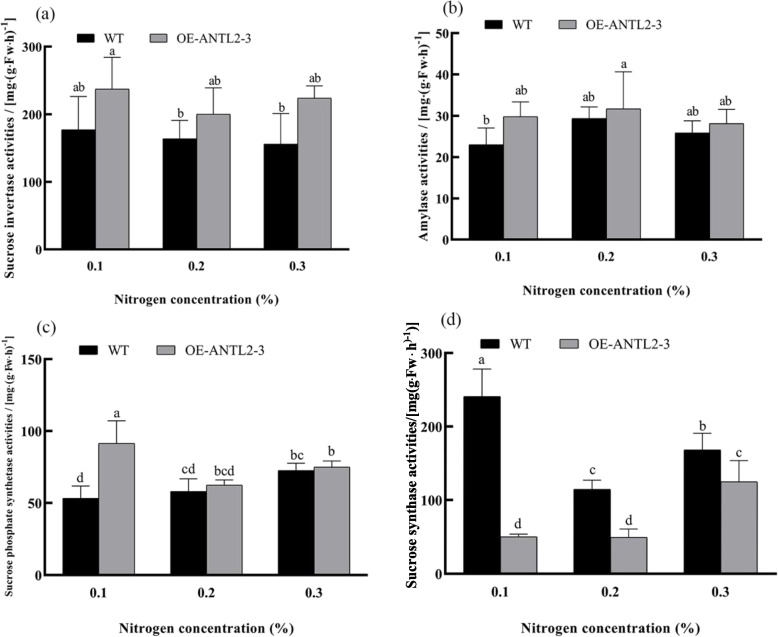 Figure 2
Figure 2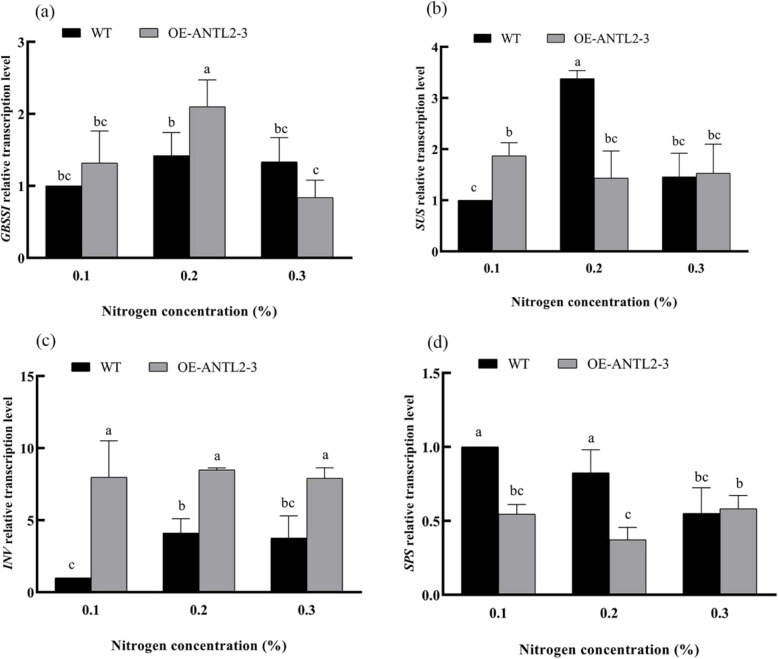 Figure 3
Figure 3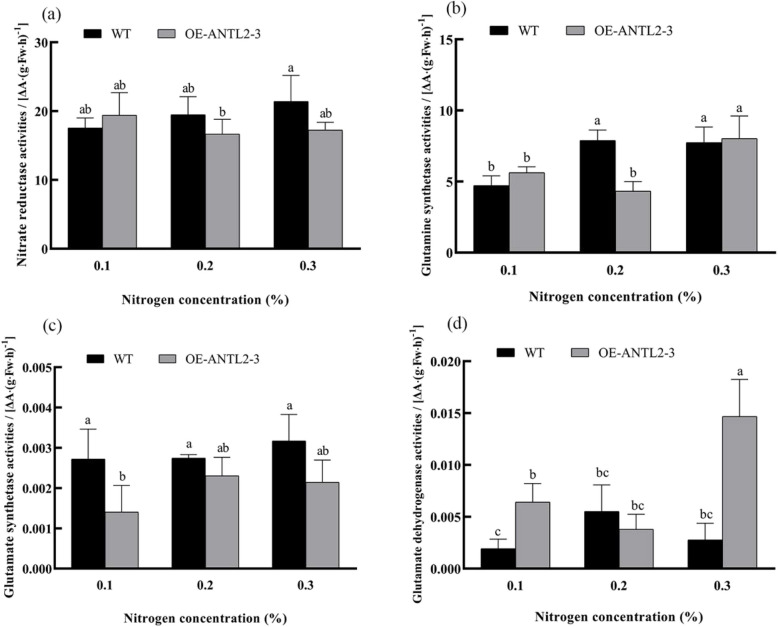 Figure 4
Figure 4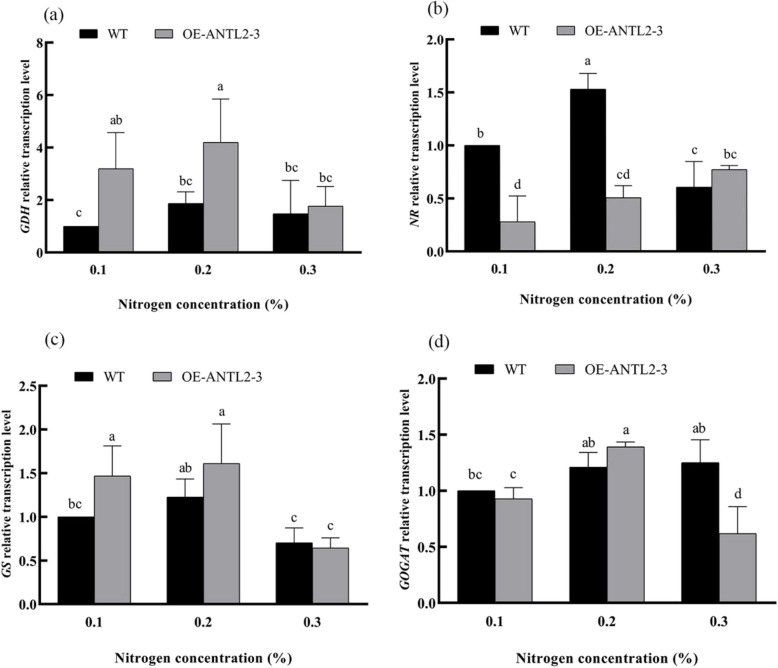 Figure 5
Figure 5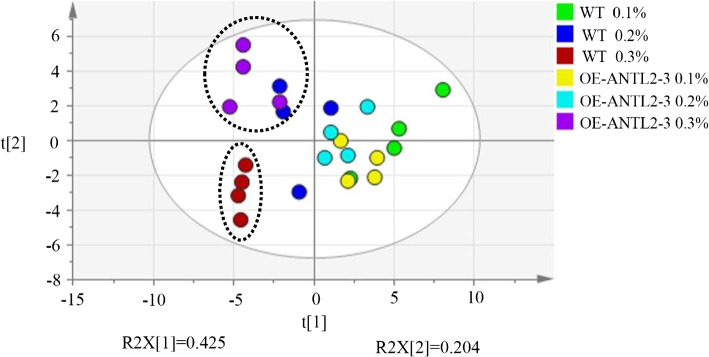 Figure 6
Figure 6- —Program of China National Tobacco Corporation
- —Program of Guizhou Branch of China National Tobacco Corporation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant nutrient uptake and metabolism · Legume Nitrogen Fixing Symbiosis · Plant Micronutrient Interactions and Effects
Introduction
Flue-cured tobacco (Nicotiana tabacum L.) is an economically significant, non-edible field crop, widely cultivated worldwide and all over China [1]. Nitrogen (N) availability is a critical limiting factor affecting biomass production and metabolic processes [2, 3]. Consequently, to achieve higher yields and improve economic efficiency, farmers often apply substantial quantities of N fertilizers [4, 5]. However, excessive nitrogen fertilizer can make the aroma become weak or be masked by a strong nicotine smell and excessive use of N fertilizers leads to environmental pollution and soil acidification [5–8]. Enhancing nitrogen use efficiency (NUE) in flue-cured tobacco plants is a crucial strategy for mitigating the environmental pressures associated with excessive N fertilizer application [9].
In most plants, nitrogen is primarily assimilated into amino acids in roots or shoots to support plant growth [10–12]. Amino acids are involved in the tricarboxylic acid cycle, generate cellular energy [13], and regulate the carbon-to-nitrogen (C/N) balance [14]. Amino acid transporters facilitate root uptake, xylem loading in roots, phloem loading in leaves, and nitrogen import into seeds. Based on sequence similarities and uptake properties, amino acid transporter families in plants are classified into two subfamilies: amino acid/auxin permeases (AAP) and amino acid polyamine and choline transporters [15–17].
Among plant amino acid transporters, lysine histidine transporter 1 (LHT1) and AAP families and their roles in nitrogen (N) and carbon (C) metabolism, growth, and development were well studied. Arabidopsis AtLHT1, highly expressed in roots, young leaves, flowers, and leaf sheaths, mediates uptake of neutral and acidic amino acids into mesophyll cells. Overexpression of AtLHT1 or cucumber homologs CsLHT1/CsLHT6 enhances amino acid uptake, while AtLHT1 loss reduces aboveground biomass and seed yield under N limitation [18–20]. In rice, OsLHT1 knockout impairs root uptake of root-synthesized amino acids, decreasing N absorption efficiency and yield but increasing seed protein content and essential free amino acids [21, 22]. NtLHT1 is a low-affinity amino acid transporter which transports neutral, acidic, non-polar, and aromatic amino acids, as well as the ethylene precursor ACC (1-aminocyclopropane-1-carboxylate), gibberellin, and indole-3-acetic acid. NtLHT1 influences key physiological processes such as cell division, elongation, and differentiation by modulating the levels of amino acids and plant hormones [23].
The AAP family exhibits functional diversity: AtAAP1, localized at root tips, epidermal cells and regulated by light, C status, and nitrate, influences Arabidopsis growth, seed development, protein storage, and yield [24, 25]. In pea (Pisum sativum), overexpression of AAP1 improved nitrogen uptake and utilization efficiency [26]. AtAAP2 in phloem facilitates N translocation from xylem to phloem, with aap2 mutants allocating more N to leaves to enhance photosynthesis and seed yield [27, 28]. AtAAP3 in root vascular tissues may mediate amino acid uptake from soil or phloem, and AtAAP5 transports cationic amino acids [29, 30]. Rice OsAAP6 overexpression increases seed storage proteins without affecting yield [27], while AtAAP8 in leaf phloem and petioles is critical for loading amino acids into phloem for seed allocation; aap8 mutants show reduced phloem loading, impaired photosynthesis, and N/C assimilation [31, 32]. Disruption of rice OsAAP3, OsAAP5, or OsAAP7—functions contrasting with LHT1—promotes tillering and increases grain yield, highlighting complementary roles in regulatory networks [33–35].
Aromatic and neutral amino acid transporters (ANTs) belong to the amino acid/auxin permease subfamily [36], with genes like ANTL2 capable of transporting diverse substrates to meet physiological demands [11]. In Arabidopsis, AtANT1 mediates the transport of aromatic and neutral amino acids, including arginine, exhibiting tissue-specific expression modulated by nutrient status and environmental cues. For instance, seedling AtANT1 expression is induced by varying nitrate concentrations, linking its activity to nitrogen signaling and amino acid homeostasis [37]. Wheat TaANT1 shows peak expression in endosperm aleurone cells 20 days post-anthesis, suggesting a role in delivering N to developing grains during the mid-filling stage. Genetic analysis in Arabidopsis reveals that the ant1 mutant accumulates more free amino acids in sieve tubes significantly than wild-type plants, indicating that ANT1 facilitates amino acid removal from the phloem, likely through involvement in phloem unloading [38, 39].
This function underscores ANT1’s role in regulating source-to-sink amino acid partitioning, where disrupted transport alters long-distance N distribution and metabolic balance. Collectively, these findings highlight ANTs as key mediators of amino acid transport, environmental responses, and nutrient allocation across plant tissues, with implications for understanding N metabolism and optimizing crop productivity. Although there are few detailed studies directly on ANT proteins in tobacco, similar regulatory mechanisms are likely present in tobacco, suggesting that these transporters are finely tuned to meet the plant's metabolic needs [40].
The amount and form of available nitrogen significantly affect plant growth, photosynthesis, yield, and quality [41, 42]. It is well established that nitrogen, carbon, and water cycles are closely interrelated during plant growth [43, 44]. Nitrogen metabolism depends on carbon metabolism for carbon skeletons and energy, while carbon metabolism relies on nitrogen metabolism for enzyme production and photosynthetic pigment synthesis [44].
Previous results showed that the relative transcription level of NtANTL2 was significantly higher in mild aromatic tobacco varieties Weining and Tianzhu than in medium aromatic Kaiyang, suggesting that NtANTL2 may be associated with amino acid content and transport. However, the mechanism by which the ANT2 family participates in the uptake and long-distance transport of nitrogenous organic compounds remains unclear. This study reports the characterization of NtANTL2 gene function in tobacco growth, enzyme activity, carbon and nitrogen metabolism, and amino acid accumulation under varying nitrogen applications.
Material and methods
Plant materials and culture condition
Tobacco variety K326 was kindly provided by China National Tobacco Corporation. Plants were cultivated in 500 mL pots containing a soil mixture (50% vermiculite) at 25 °C under a 16 h light/8 h dark photoperiod, with a light intensity of 52 μmol·m⁻^2^·s⁻^1^ and relative humidity of 80%. Urea was added to achieve final nitrogen concentrations of 0.1%, 0.2%, and 0.3%.
NtANTL2 transgenic plant vector construction and genetic transformation
The pCAMBIA2301G vector, Agrobacterium tumefaciens strain LBA4404, and PEASY-Blunt vector were provided by the College of Life Science, Yangtze University. The pCAMBIA2301G vector contains the CaMV 35S promoter, which is a constitutive promoter known for driving gene expression in tobacco. RNA was extracted from tobacco K326 plants and reverse-transcribed to synthesize complementary DNA (cDNA). The NtANTL2 gene was amplified using a high—fidelity Pfu DNA polymerase, and the resulting fragment was ligated downstream of the CaMV 35S promoter in the pCAMBIA2301G expression vector to ensure its expression in transgenic plants. Escherichia coli DH5α was transformed with the recombinant pCAMBIA2301G-NtANTL2 plasmid. Positive clones were identified, plasmid DNA was extracted, and transformed into Agrobacterium tumefaciens LBA4404. Selected colonies were cultured and stored at − 80 °C. Tobacco was transformed using the Agrobacterium-mediated method with K326 leaf discs as explants to generate T3 NtANTL2-overexpressing transgenic plants. RT-qPCR analysis confirmed the overexpression of NtANTL2 in transgenic lines. After 30 days of growth, both control and transgenic tobacco plants were harvested for further analysis. All the primers used were listed in Supplemental Table S1.
RNA extraction and qPCR analysis
OE-ANTL2-3 and WT tobacco plants were cultivated to the six-leaf stage, and plants with similar growth were transplanted to soil with nitrogen levels of 0.1%, 0.2%, and 0.3%. Total RNA was extracted from approximately 100 mg of tobacco leaves using TRIzol reagent, following the manufacturer’s instructions (Invitrogen, USA). One microgram of total RNA was reverse-transcribed using the PrimeScript™ II 1 st Strand cDNA Synthesis Kit (TaKaRa, Japan). The diluted cDNA (fivefold) was used as the template for RT-qPCR analysis using a CFX Connect Real-Time System (BIO-RAD). Relative transcription levels of NtANTL2-3 genes, carbon metabolism-related genes (GBSSI, SUS, INV, SPS), and nitrogen metabolism-related genes (GDH, NR, GS, GOGAT) were analyzed. Relative expression levels were calculated using the 2⁻ΔΔCT method [45]. All the primers used for RT-qPCR were listed in Supplemental Table S1. All experiments were repeated at least three times with similar results.
Morphological and physiological measurements of tobacco seedlings
Uniformly grown seedlings at the six-leaf stage were planted in pots with varying nitrogen concentrations. Plant height and root length were measured after 30 days, and fresh/dry weights of stems, leaves, and roots were recorded. Chlorophyll content were analyzed by spectrophotometer method [46]. Soluble carbohydrate and protein contents were measured following established protocols [47, 48]. Enzyme activities, including sucrose invertase, sucrose synthase, sucrose phosphate synthase, nitrate reductase, glutamine synthetase, and glutamate synthase, were measured as described previously [15, 16, 49, 50]. Four biological replicates with five plants per replicate were used.
Nitrogen treatment and phenotypic analysis
Full and pest-free K326 (Wester, WT) seeds and T3 generation of overexpressing NtANTL2 transgenic (OE-ANTL2) seeds were disinfected in 75% alcohol for 30 s and then washed with sterile water for 2–3 times. After disinfection, K326 seeds were inoculated on MS medium, and OE-ANTL2 seeds were inoculated on 100 mg L^−1^ kanamycin + MS medium. Plants were cultivated under dark conditions 2 days, and then transferred to conditions with a photoperiod of 16 h light/8 h darkness at 25 ℃ for about 30 days.
Soil samples were collected from the field, air-dried, and mixed with a matrix at a 1:1 volume ratio. Initial soil N content was 0.097%, measured via the Kjeldahl method: samples were digested with concentrated sulfuric acid and a catalyst to convert N into ammonium, which was then alkalized, distilled, and absorbed by boric acid before titration with standard acid to calculate total N. Urea was added to adjust mixed substrate N levels to 0.1% (control), 0.15%, 0.2%, 0.3%, 0.4%, and 0.5%. After transplanting tobacco seedlings into these substrates, high-N treatments (0.4% and 0.5%) caused chlorosis or plant death within 20 days, while plants under 0.1%, and 0.15% N application showed no significant differences. Thus, 0.1%, 0.2%, and 0.3% N concentrations were selected for further experiments. Four biological replicates were performed, each containing five plants.
Amino acids analysis
Metabolomic analysis of amino acids was performed by Wuhan Metware Biotechnology Co., Ltd. Samples were lyophilized, crushed into powder, and extracted with 70% aqueous methanol. The extracts were analyzed using an ultra-performance liquid chromatography–tandem mass spectrometry (UPLC–MS/MS) system. Amino acids with a variable importance in projection ≥ 1 and an absolute log_2_ fold change (log_2_FC) ≥ 1 were considered differentially regulated. Three biological replicates were conducted, with four plants included in each replicate.
Statistical analysis
One-way two-factor ANOVA followed by Fisher’s protected LSD multiple comparison was used to evaluate differences among conditions. Statistical analyses were performed using SPSS 20.0 (Chicago, USA), and significance was indicated by different letters (a, b, c) for p < 0.05.
Results
NtANTL2 overexpression enhances growth under high nitrogen conditions
The NtANTL2 gene family in tobacco comprises five copies (NtANTL2-1 to NtANTL2-5), with tissue-specific expression patterns suggesting functional specialization across tissues. Quantitative analysis revealed that NtANTL2-1, NtANTL2-2, NtANTL2-4, and NtANTL2-5 exhibited the highest transcript levels in leaves, followed by stems, with minimal expression in roots. In contrast, NtANTL2-3 displayed preferential expression in stems (Figure S1). Five stable overexpression lines (OE-ANTL2-1 to OE-ANTL2-5) were generated, and OE-ANTL2-3 exhibited the highest NtANTL2 expression among transgenic lines and it was selected for detailed phenotypic and molecular characterization. Relative expression analysis of NtANTL2-3 in transgenic and wild-type (WT) plants under varying nitrogen regimes is presented in Figure S2.
Tobacco seedlings (OE-ANTL2-3 and WT) were cultivated to the six-leaf stage before transplantation into substrates amended with 0.1%, 0.2%, or 0.3% nitrogen. After 30 days of treatment, OE-ANTL2-3 plants exhibited significantly enhanced agronomic traits compared to WT plants, including increased leaf length, leaf width, and plant height (Fig. 1), indicating improved growth performance under tested N conditions.Fig. 1. Growth of wide type and OE-ANTL2-3 plants under different nitrogen conditions
At a nitrogen level of 0.1%, WT plants exhibited greater plant height than OE-ANTL2-3 plants, but other traits showed no significant differences. At 0.2% nitrogen, OE-ANTL2-3 plants demonstrated significantly higher plant height, shoot fresh weight, and root length than WT plants. At 0.3% nitrogen, OE-ANTL2-3 plants showed remarkable increases in plant height (2.31 times), shoot fresh weight (3.23 times), root length (1.47 times), root fresh weight (6.06 times), and root dry weight (5.67 times) compared to WT plants. These results suggest that OE-ANTL2-3 plants have enhanced tolerance to high nitrogen levels (Table 1). Table 1. Investigation on agronomic characters of transgenic plants and wild type plants under different nitrogen levelsNitrogen %MaterialPlant height (cm)Shoot FW (g)Shoot DW (g)Root length (cm)Root FW (g)Root DW (g)0.1WT63.33 ± 1.20a165.67 ± 6.39a14.81 ± 1.24a11.27 ± 0.15a7.00 ± 0.58a1.18 ± 0.11aOE-ANTL2-349.33 ± 1.76b148.33 ± 10.53ab12.85 ± 1.66ab10.00 ± 0.58ab6.50 ± 0.29a0.79 ± 0.21ab0.2WT33.33 ± 1.76c113.00 + 3.00b6.03 ± 0.32c7.67 ± 0.17c2.00 ± 0.00 cd0.30 ± 0.02 cdOE-ANTL2-346.33 ± 1.86b163.33 ± 16.76a10.15 ± 1.41b9.33 ± 0.44b3.33 ± 0.67bc0.70 ± 0.19bc0.3WT19.33 ± 3.18 d44.00 ± 21.39c2.39 ± 1.26c6.33 ± 0.33c0.77 ± 0.15 d0.12 ± 0.03 dOE-ANTL2-344.67 ± 3.93b142 ± 9.64ab10.17 ± 1.41b9.33 ± 0.67b4.67 ± 0.88b0.68 ± 0.11bcThe data in the table are mean ± standard error. Different lowercase letters in the same column indicated a difference of 0.05 levels
Effects of NtANTL2 overexpression on carbon metabolism
Carbon metabolism is a fundamental process in tobacco leaves, and its regulation is closely linked to the activity of associated enzymes. Enzyme activities of sucrose invertase (EC 3.2.1.26), α-amylase (EC 3.2.1.1), sucrose phosphate synthetase (2.4.1.14), and sucrose synthase (EC 2.4.1.13) were measured in WT and OE-ANTL2-3 plants under varying nitrogen levels. Sucrose invertase and amylase activities in OE-ANTL2-3 plants were similar to those in WT plants across all three nitrogen levels (Fig. 2a and b). Sucrose phosphate synthetase activity in OE-ANTL2-3 plants was 1.5 times higher than in WT plants at 0.1% nitrogen but showed no significant differences at 0.2% and 0.3% nitrogen levels (Fig. 2c). Conversely, sucrose synthase activity in OE-ANTL2-3 plants was significantly lower than in WT plants under all nitrogen levels, decreasing by 79.23% and 57.03% at 0.1% and 0.2%, respectively. At 0.3% nitrogen, OE-ANTL2-3 plants exhibited higher sucrose synthase activity than at lower nitrogen levels (Fig. 2d).Fig. 2. Comparison of carbon metabolism related enzyme activities between transgenic plants and wild plants under different nitrogen levels
Expression levels of carbon metabolism-related genes were also analyzed under different nitrogen levels. In OE-ANTL2-3 plants, the relative transcription levels of GBSSI, SUS, INV, and SPS genes varied significantly. For example, GBSSI expression was 47.82% higher at 0.2% nitrogen, and INV expression was consistently higher than WT across all nitrogen levels (Fig. 3). Regarding the discrepancies between gene expression and enzyme activity, potential post—transcriptional and post—translational regulation mechanisms could cause the differences.Fig. 3. Comparison of transcription levels of carbon metabolism related genes between transgenic plants and wild plants under different nitrogen levels
Effects of NtANTL2 overexpression on nitrogen metabolism
Key nitrogen metabolism enzymes, including nitrate reductase (NR, EC 1.6.6.1), glutamine synthetase (GS, EC 6.3.1.2), glutamate synthase (GOGAT, EC 1.4.1.14), and glutamate dehydrogenase (GDH, EC 1.4.1.2), were analyzed. GDH activity in OE-ANTL2-3 plants was significantly higher than in WT plants at 0.1% and 0.3% nitrogen, increasing by 236.84% and 425.00%, respectively. The relative transcription level of GDH was also elevated in OE-ANTL2-3 plants at lower nitrogen levels.
Glutamine (Gln) is the first amino acid synthesized in the nitrogen assimilation process in plants. GS catalyzes the conversion of glutamate (Glu) and NH4^+^ into glutamine at the expense of ATP. Tobacco plants primarily absorb nitrate (NO3^−^) and ammonium (NH4^+^) from the soil through their roots. Nitrate must be reduced by NR and nitrite reductase (NiR, EC 1.7.7.1) to ammonia, which is then assimilated into amino acids through the GS and GOGAT system, a crucial step in nitrogen metabolism.
The NR activity in OE-ANTL2-3 and WT plants is comparable (Fig. 4). The GS activity in OE-ANTL2-3 plants is lower than that in WT plants at a 0.2% nitrogen level, with a decrease of 45.20%. At other nitrogen levels, no difference is found between OE-ANTL2-3 plants and WT plants. The GOGAT activity of OE-ANTL2-3 plants is lower than that of WT plants at a 0.1% nitrogen level, showing a reduction of 48.15%. The GDH activity of OE-ANTL2-3 plants at 0.1% and 0.3% nitrogen levels is significantly higher than that of WT plants, increasing by 236.84% and 425.00% respectively. The GDH activity of WT plants remains unchanged among all nitrogen levels. The GDH activity of OE-ANTL2-3 plants at a 0.3% nitrogen level is significantly higher than that at 0.1% and 0.2% nitrogen levels, with increases of 129.69% and 286.84%, respectively.Fig. 4. Comparison of nitrogen metabolism related enzyme activities between transgenic plants and wild plants under different nitrogen levels
The gene expression levels of GDH, NR, GS, and GOGAT were analyzed under three different nitrogen levels. The relative transcription level of the GDH gene in OE-ANTL2-3 plants is higher than that in WT plants at 0.1% and 0.2% nitrogen levels, with increases of 219.22% and 124.07% respectively. At 0.1% and 0.2% nitrogen levels, the relative transcription level of the NR gene is significantly lower than that of WT plants, decreasing by 72.02% and 66.91% respectively. Under a 0.1% nitrogen level, the relative transcription level of the GS gene is significantly higher than that of WT plants, increasing by 46.66%. Under 0.3% nitrogen level, the relative transcription level of the GOGAT gene is significantly lower than that of WT plants, decreasing by 46.21%. The enzyme activity and gene expression of GS and GDH were consistent. In OE plants, GS activity was lower than wide type plants (Figs. 4 and 5).Fig. 5. Comparison of transcription levels of nitrogen metabolism related genes between transgenic plants and wild plants under different nitrogen levels
According to the principal component analysis of nitrogen compounds content in OE-ANTL2-3 plants and WT plants (Fig. 6), the contents of nitrogen substances in tobacco leaves of OE-ANTL2-3 plants and WT plants were different under different nitrogen levels, among which the contents of nitrogen substances in tobacco leaves of OE-ANTL2-3 plants and WT plants were the most different under 0.3% nitrogen level. They were obviously divided into two categories. Therefore, the subsequent analysis focused on the difference of nitrogen content in tobacco leaves between OE-ANTL2-3 plants and WT plants at 0.3% nitrogen level.
Fig. 6. Scatter plots (PCA plots) of principal component analysis of nitrogen containing substances in transgenic plants and wild plants under different nitrogen levels. Three biological replicates were conducted, with four plants included in each replicate. Unit: ug/g dry weight
Effects of NtANTL2 Overexpression on amino acid metabolism
As can be seen from Table 2, the metabolites with significant differences (P < 0.05) in nitrogen content of tobacco leaves between OE-ANTL2-3 plants and WT plants include: aspartic acid, glycine, threonine, arginine, gamma-aminobutyric acid, tyrosine, valine, histidine, methionine, leucine, phenylalanine, ornithine, lysine, tyramine, putrescine, phenylethylamine, and isoamylamine. At 0.3% nitrogen level, the content of aspartic acid in OE-ANTL2-3 plants was significantly lower than that in WT plants, and the contents of glycine, threonine, and tyrosine were significantly increased, while the contents of histidine, arginine, gamma-aminobutyric acid, valine, methionine, leucine, phenylalanine, ornithine, lysine, tyramine, putputylamine, phenethylamine and isoamylamine were significantly increased. Table 2. Multiple comparisons of nitrogenous substances in tobacco leaves of transgenic plants and wild plants under 0.3% nitrogen level (unit: ug/g dry weight)ParametersWT 0.3%OE-ANTL2-3 0.3%Sigaspartic acid1261.6 ± 223.91816.02 ± 92.830.010histidine174.04 ± 15.65306.4 ± 79.620.020glycine50.78 ± 4.3575.03 ± 5.530.004threonine418.68 ± 34.8519.76 ± 27.260.009arginine175.89 ± 26.98343.33 ± 82.260.011γ-aminobutyric acid212.9 ± 27.3270.94 ± 28.140.025alanine373.37 ± 32.77389.09 ± 30.050.506proline247.45 ± 46.65329.6 ± 46.410.069tyrosine111.56 ± 23.6221.07 ± 41.290.006valine180.29 ± 14.73267.92 ± 48.270.017methionine34.64 ± 4.641.94 ± 1.570.049tryptophan129.49 ± 19.06208.66 ± 61.540.055leucine193.22 ± 13.43286.57 ± 62.610.031phenylalanine172.54 ± 13.59239.39 ± 38.520.022ornithine3.50 ± 0.835.8 ± 0.730.013lysine147.49 ± 27.78261.9 ± 48.010.010tyramine124.71 ± 33.69214.32 ± 34.710.018tetrametnylened iamine149.26 ± 33.31310.92 ± 83.510.011phenylethylamine45.36 ± 10.3470.54 ± 15.020.033isoamylamine584.15 ± 62.711227.7 ± 260.860.014
Meanwhile, the chlorophyll, soluble sugar, soluble protein, total nitrogen, and total protein contents in transgenic and wild plants under different nitrogen levels were analyzed. Results showed no differences in these parameters between the two plant types, indicating genetic modification did not affect their physiological substance levels in response to varying nitrogen conditions (Fig. S3, S4, and S5).
Discussion
Nitrogen, a critical macronutrient for plant growth, often constrains crop yield due to its limited availability and the intricate balance required for optimal carbon -nitrogen metabolism. Efficient N use hinges on dynamic partitioning between source (e.g., leaves) and sink (e.g., roots, reproductive organs) tissues, which integrates N uptake, photosynthetic enzyme activity (e.g., Rubisco, phosphoenolpyruvate carboxylase), and phloem-mediated amino acid transport [51, 52]. Disruptions in this balance—such as excessive N fertilization—can lead to soil salinization, nitrate accumulation, and stunted growth, underscoring the need to develop crops with improved N tolerance and utilization [3, 53].
In this study, overexpression of NtANTL2, a tobacco amino acid transporter gene, demonstrated enhanced tolerance to high N conditions, primarily through coordinated regulation of C-N metabolism. Transgenic plants exhibited superior agronomic traits, including increased biomass and leaf dimensions, even at elevated N levels (Table 1, Fig. 1), aligning with the pivotal role of amino acid transporters in mediating source-sink nutrient allocation [54, 55]. Maintaining stable chlorophyll content under high N stress suggests preserved photosynthetic capacity, a critical factor for sustaining C assimilation and NUE [56, 57].
Nitrate reductase (NR), a key enzyme in N assimilation, showed increased activity in NtANTL2-overexpressing plants, indicating enhanced conversion of nitrate to nitrite and subsequent amino acid biosynthesis [58, 59]. This was corroborated by elevated levels of specific amino acids (e.g., histidine, arginine, γ-aminobutyric acid), which serve as both N storage molecules and C skeletons for energy metabolism [60, 61]. Conversely, reduced aspartic acid content in transgenic lines may reflect reallocation of C skeletons toward more N-efficient metabolic pathways, illustrating the role of NtANTL2 in fine-tuning C-N balance [62, 63].
The tissue-specific expression of NtANTL2 family members—with most copies preferentially expressed in leaves and stems, and NtANTL2-3 enriched in stems—suggests functional specialization in organ-specific N transport (Fig. S1). This aligns with previous findings where amino acid transporters participate in phloem loading/unloading and long-distance N translocation, with mutants like ant1 showing disrupted sieve tube amino acid dynamics [11, 38]. By enhancing amino acid allocation to sinks and optimizing N metabolism, NtANTL2 overexpression likely mitigates the negative effects of high N, such as reduced photosynthesis and metabolic imbalance [28, 31].
Amino acid transporters are central to integrating environmental cues (e.g., nitrate availability) with metabolic responses, as evidenced by nitrate-inducible expression of homologous ANT genes in Arabidopsis and wheat [37]. Here, NtANTL2’s role in upregulating N assimilation genes and improving N-containing compound levels under high N stress highlights its potential as a molecular target for breeding nitrogen-efficient tobacco cultivars. Such strategies are vital for sustainable agriculture, addressing both yield stability and environmental concerns associated with excessive fertilizer use [64, 65].
While this study establishes NtANTL2 as a key regulator of N tolerance and C-N metabolism, unresolved questions remain about the precise signaling pathways and interacting proteins mediating its effects. Elucidating these mechanisms will deepen our understanding of amino acid transporter multifunctionality—including roles in stress resistance and pathogen defense—and enable targeted manipulation of metabolic networks to enhance crop productivity under challenging N regimes [33, 66].
In conclusion, NtANTL2 overexpression improves high N tolerance in tobacco by optimizing amino acid transport, enhancing N assimilation, and balancing C-N metabolism. These findings provide a foundation for developing molecular breeding strategies that reconcile agricultural productivity with environmental sustainability, underscoring the importance of amino acid transporters in shaping crop responses to nutrient stress.
Conclusion
Overexpression of NtANTL2 significantly improves plant nitrogen uptake and utilization under high nitrogen conditions. Under normal nitrogen conditions, WT plants exhibit better growth than NtANTL2-overexpressing plants; however, NtANTL2-overexpressing plants demonstrate superior tolerance to high nitrogen stress. These plants show increased levels of histidine, arginine, gamma-aminobutyric acid, valine, methionine, leucine, phenylalanine, tryptophan, ornithine, lysine, tyramine, putrescine, phenylethylamine, and isovaleric acid, while aspartic acid levels are reduced. Based on previous findings that NtANTL2 inhibition significantly increases aspartic acid content (data not shown), it can be concluded that NtANTL2 regulates nitrogen uptake and influences amino acid composition in tobacco, ultimately affecting the plant's carbon and nitrogen metabolism. These findings provide a molecular basis for breeding high-efficiency nitrogen use tobacco varieties.
Supplementary Information
Supplementary Material 1: Fig. S1 Identification of the relative transcription levels of NtANTL2-3 in transgenic plants and wild-type plants. Fig. S2 Identification of the relative transcription levels of NtANTL2-3 in transgenic plants and wild type plant. Fig. S3 Comparison of chlorophyll contents between transgenic plants and wild plants under different nitrogen levels. Fig. S4 Comparison of soluble sugar and soluble protein contents between transgenic plants and wild plants under different nitrogen levels. Fig. S5 Comparison of total nitrogen contents and total protein contents between transgenic plants and wild plants under different nitrogen levels.Supplementary Material 2: Supplementary Table 1. Primer names and sequences of clone NtANTL2.