Life history of Papilio (Druryia) meriones Felder & Felder (Lepidoptera, Papilionidae) from Madagascar
Sixun Ge, Lili Ren, Shaoji Hu

TL;DR
This paper describes the life cycle of a swallowtail butterfly species found only in Madagascar, based on observations from collected eggs and rearing experiments.
Contribution
The study provides the first detailed life history description of Papilio meriones, a Madagascan-specific swallowtail butterfly.
Findings
Eggs of Papilio meriones were collected during an expedition to Madagascar.
A successful rearing experiment allowed the documentation of all developmental stages of P. meriones.
Abstract
The swallowtail butterflies (Lepidoptera: Papilionidae) have been researched extensively as a model species in biology, ecology, and conservation biology. During a recent expedition to Central and South Madagascar, eggs of a Madagascan-specific swallowtail butterfly, Papiliomeriones, were collected. After a successful rearing experiment, the life history of P.meriones is described with all developmental stages illustrated.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5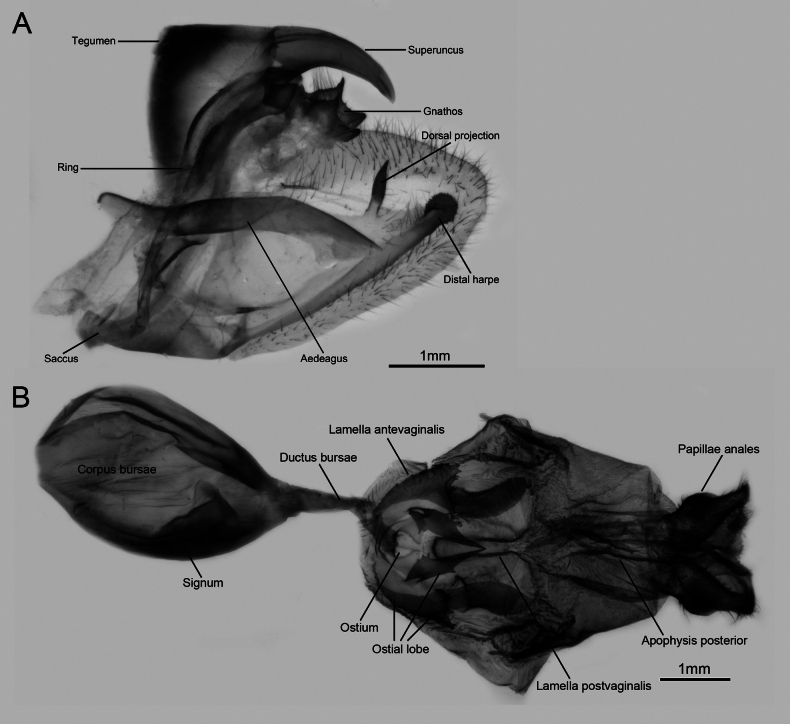 Figure 6
Figure 6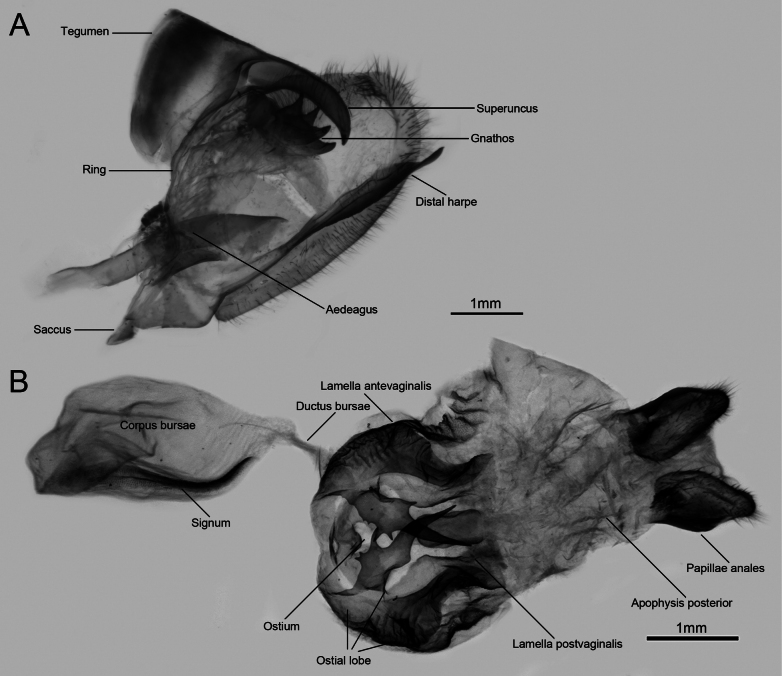 Figure 7
Figure 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLepidoptera: Biology and Taxonomy · Plant and animal studies · African Botany and Ecology Studies
Introduction
The swallowtail butterflies (Lepidoptera: Papilionidae) are one of the most famous and well-studied insect groups, with many species serving as model organisms in evolutionary biology, ecology, genomics, and conservation biology (e.g., Kunte 2009; Kunte et al. 2014; Dupuis and Sperling 2015). The genus Papilio Linnaeus, 1758, which includes over 200 known species, is considered the largest genus in the family (Zakharov et al. 2004; Condamine et al. 2012; Nakae 2021). Despite numerous studies carried out on interspecific phylogenetics within the group over the past few decades, it was only in recent years that the classification within the genus was revised (Condamine et al. 2023) due to its vast diversity.
Papilio (Druryia) meriones C. Felder & R. Felder, 1864 is a distinct species found in Madagascar. It was formerly a subspecies of the African mocker swallowtail, P.dardanus Yeats, 1776, a well-known polymorphic Batesian mimic that served as a model organism in evolutionary biology (Clarke and Sheppard 1960; Timmermans et al. 2014, 2020). Intriguingly, the closely related species, P.meriones has females with “male-like” wing patterns that are monomorphic and non-mimetic. The evolution of the “male-like” female pattern of P.dardanus and its two closest relatives, P.humbloti Oberthür, 1888 and P.meriones, has been a source of speculation, discussion, and ongoing investigation for over 150 years (e.g., Trimen 1869; Poulton 1924; Ford 1936; Bernardi 1963; Vane-Wright 1978; Clarke et al. 1985; Vane-Wright and Smith 1991; Timmermans et al. 2017, 2020). Despite previous studies into the evolution and mimicry of these species, our understanding of their immature stages remains limited. The eggs and larvae of P.meriones were obtained during a recent expedition to Central and South Madagascar. After a successful rearing experiment, the morphological characters and duration of each immature stage are reported herein.
Material and methods
Eggs and larvae of P.meriones were collected in the forest near Tobindranon’i Mantasoa (18.998455°N, 47.874170°E). Shoots and leaves of the host plant (Citrus sp.) bearing eggs or larvae were clipped from the host plant and put into plastic boxes. Branches of the host plant with healthy leaves were also collected and sealed in plastic bags.
The rearing experiment was performed without using a climate chamber, and the ambient temperature was approximately 25 °C, with a humidity of 80%. Each individual was observed and recorded each day. Waste and leaf residue were removed from the boxes on a daily basis to keep the environment clean. The size of eggs and body length of each instar larva, prepupa, and pupa were measured.
The immature stages were photographed using an interchangeable-lens digital camera. Measurements and figures of larval head capsules were made using a ZEISS AxioZoom V16 stereomicroscope with a ZEISS Axiocam 503 color camera. Photographs were combined using ZEN 2.3 (blue edition). Final plates were prepared in Adobe Photoshop CC (adobe.com).
The specimen collection has been approved by the government of Madagascar, with the authorization permit number: N°11019/FAC.Sci/D. Specimens used in this study have been deposited in Beijing Forestry University and are available for re-examination by any researcher on request to Si-Xun Ge.
Results
Developmental stages
Eggs. Female adults laid eggs singly on the upper side or underside of leaves (Fig. 1); sometimes eggs were even laid on the new shoots at the bottom of the plant or on old branches. Eggs spherical, 1.00–1.05 mm in diameter. Surface smooth with small dents, glossy yellowish with pearl sheen (Fig. 1A), gradually turning dark brownish as they develop (Fig. 1B, C). Duration of eggs about 4–6 d.
Eggs of P.merionesA freshly laid egg B about 2-day-old egg C egg about to hatch.
Larva. 1^st^ instar larva (Fig. 2A, B): Maximum width of head capsule 0.92–0.97 mm (Fig. 3A). Body length 5–6 mm. Body olive green with olive or whitish verrucae. Head capsule glossy black. Thoracic verrucae whitish, with the verruca on the 1^st^ thoracic segment extremely elongated. Abdominal verrucae variable, with those on the 1^st^–3^rd^ and 6^th^–8^th^ abdominal segments darkish olive, while those on the 4^th^–5^th^ and 8^th^–9^th^ whitish, verrucae elongated in the last two abdominal segments. All verrucae with long setae. Duration of 1^st^ instar larvae about 4–6 d.
Larvae of P.merionesA 1st instar larva, dorsal view B 1st instar larva, head capsule C 2nd instar larva, dorsal view D 2nd instar larva, head capsule E 3rd instar larva, dorsal view F 3rd instar larva, head capsule G 4th instar larva, dorsal view H 4th instar larva, head capsule I 5th instar larva (greenish form), dorsal view J 5th instar larva (greenish form), head capsule K 5th instar larva (whitish-spots form), dorsal view L 5th instar larva (whitish-spots form), head capsule.
Head capsules of P.meriones larvae A 1st instar larva B 2nd instar larva C 3rd instar larva D 4th instar larva E 5th instar larva.
2^nd^ instar larva (Fig. 2C, D): Maximum width of head capsule 1.33–1.39 mm (Fig. 3B). Body length 8–10 mm. Body olive yellow with whitish bands, verrucae olive yellow or whitish. Head capsule glossy black. Thoracic segments and verrucae largely whitish; verruca on the 1^st^ thoracic segment robust and elongated, while those on the 2^nd^–3^rd^ thoracic segments short. The 3^rd^, 8^th^ and 9^th^ abdominal segments whitish. Abdominal verrucae largely small and short, except those on 9^th^ and 10^th^ abdominal segments, which are elongate. All verrucae with long setae. Duration of 2^nd^ instar 5–6 d.
3^rd^ instar larva (Fig. 2 E, F): Maximum width of head capsule 1.98–2.09 mm (Fig. 3C). Body length 9–13 mm. Body olive green to olive yellow with whitish markings. Head capsule glossy dark olive with clypeus area paler. The 1^st^ thoracic verrucae large, elongate into a horn-like shape. Verrucae on 8^th^ and 9^th^ abdominal segments elongate. All elongate verrucae whitish, with rather short setae. Verrucae on other segments rather weak, with a pair of whitish spots on the basal part. The 3^rd^ abdominal segments with a pair of large sub-rhombus whitish spots. In this instar, the larvae evert dark reddish brown osmeteria when disturbed. Duration of 3^rd^ instar 5–6 d.
4^th^ instar larva (Fig. 2G, H): Maximum width of head capsule 2.7–2.87 mm (Fig. 3D). Body length 13–16 mm. Body olive green to olive yellow with whitish markings, whitish spots near the weak verrucae with slightly blueish tone. Head capsule olive yellow on frons area, while olive green on clypeus area. The 1^st^ thoracic verrucae large, elongate into a horn-like shape. Verrucae on the 8^th^ abdominal segment reduced, while on 9^th^ abdominal segment elongate. All elongated verrucae whitish, with setae rather short. The 3^rd^ abdominal segment with a pair of large, irregular whitish spots extending to the 2^nd^ and 4^th^ abdominal segments. Duration of 4^th^ instar 6–8 d.
5^th^ (final) instar larva (Fig. 2I–L): Maximum width of head capsule 4.18–4.45 mm (Fig. 3E). Body length 18.5–45 mm. Body colour largely green with densely scattered pale whitish spots (sometimes with whitish markings), whitish spots near the weak verrucae with blueish tone. Head capsule completely greenish. The first thoracic verrucae on the pronotum reduced, short, and blunt without setae. Verrucae on 8^th^ abdominal segment absent; while on 9^th^ abdominal segment weakened, short and robust. In this instar, the larvae were observed with two different colour forms. The whitish-spots form has a pair of large irregular whitish spots on the 3^rd^ abdominal segment, extending to the 4^th^ abdominal segment (Fig. 2K, L); while the greenish form lacks whitish spots but has a slightly paler 3^rd^ abdominal segment (Fig. 2I, J). Duration of 5^th^ instar 14–16 d.
Prepupa. 38–45 mm in length. Body colour light emerald green with semi-transparent tone (Fig. 4A), similar to the 5^th^ instar larva. Duration of prepupa 1–2 d.
Prepupa and pupa of P.merionesA prepupa B greenish form pupa, dorsal view C greenish form pupa, ventral view D greenish form pupa, lateral view E whitish spot form pupa, lateral view.
Pupa. 36–42 mm in length. Body emerald green (Fig. 4B–E). Pupa mimics a curved leaf. Head angular with a pair of emerald green processes, with its apical part close together. Pupae were observed as shown in two different colour forms, corresponding to the colour form of 5^th^ instar larvae. Pupae of the whitish-spots form larvae have a sub-triangular whitish spot near the basal part of the wing, while the greenish form lacks the whitish spot. Duration of pupa 13–16 d.
Adults. Male (Fig. 5A, B): Length of fore-wing about 50 mm. Upperside (Fig. 5A): Fore- and hind-wings largely whitish. Fore-wing with outer margin blackish, broadening at the apex. Sub-apical area with an oval whitish spot. Hind-wing with blackish outer margin, extending from space 5 to space 8. Spaces 2–4 with crescent-shaped blackish spots near the outer margin. Spaces 1b, 2, 4, 6 and 7 with sub-rectangular blackish spots. Tornus with a blackish spot. Tails blackish, with tips pale creamy whitish. Underside similar to the upperside (Fig. 5B), but with all blackish markings pale brownish. Hind-wing with brownish lines well-developed in all spaces, forming a radial pattern.
Adults of P.merionesA male upperside B male underside C female upperside D female underside.
Male genitalia (Fig. 6A): Highly sclerotised. Ring slightly wavy in the upper part, with distinctly developed bicornate gnathos on the tip; tegumen broad; superuncus broad at the base, slightly narrowed towards the tip and curved ventrally in lateral view; saccus flat, more or less reduced. Valve sub-triangular shaped, slightly elongate with its distal part blunt; the distal harpe strongly sclerotised, toothed disc-shaped; the dorsal projection digit-shaped with an acutely pointed tip. Aedeagus robust, more or less wavy.
Genitalia of P.merionesA male genitalia B female genitalia.
Female (Fig. 5C, D): Similar to male, but fore-wing with a blackish marking in the discal cell. On the upperside, it extends from the basal of vein Sc to the sub-apical part of the discal cell (Fig. 5C), while on the underside it is developed as an oblique sub-rectangular dark brownish spot in the median part of the discal cell (Fig. 5D).
Female genitalia (Fig. 6B): Apophysis posterior short and slender; lamella postvaginalis lobe-shaped with inner distal edge extending to the centre. Ostial lobe strongly sclerotised; inner part of the ostial lobe multi-toothed sub-triangular or sub-quadrangular shaped, with its apical tip rather elongate; outer part of the ostial lobe flange-shaped, with its outer margin multi-dentate and with a prominent protrusion in the sub-median part.
Discussion
The African mocker swallowtail, Papiliodardanus, is a well-known example of a polymorphic Batesian mimic. It has played an important part in the debate over the evolution of phenotypic variation. Studies on the mimicry of the species date back over a century (Trimen 1869; Poulton 1924; Ford 1936). However, in terms of systematics and taxonomy, the first molecular approach involving the subgenus Druryia was only recently carried out (Condamine et al. 2023); it supported P.meriones as a distinct species rather than a subspecies of P.dardanus.
Morphological data from immature stages are extremely important in taxonomy, systematics, and evolutionary biology in Lepidoptera. Our study describes the life history of a Madagascan endemic species, P.meriones, with a focus on the morphology of immature stages. As closely related species, P.dardanus, P.meriones, and P.humbloti exhibit distinct differences in their male genitalia, particularly in the dorsal projection on the inner side of the valve (Carpenter 1948; this study). Papiliomeriones possesses a strongly sclerotized distal harpe, apically toothed and disc-shaped, with a sclerotized digit-shaped dorsal projection (Fig. 6A). In contrast, the distal harpe of P.d.dardanus is highly elongated beyond the valve (Fig. 7A), laminar in structure, and lacks a dorsal projection. Specimens from the eastern Madagascan populations (including P.d.tibullus, P.d.antinorii, P.d.byatti, and P.meriones) always display a digit-shaped dorsal projection with an acutely pointed tip. In contrast, western specimens (P.d.dardanus, P.d.meseres and P.humbloti) show a highly reduced or vestigial dorsal projection. In the female genitalia, P.meriones and P.d.dardanus also exhibit distinct differences (Figs 6B, 7B), primarily manifested in the morphology of the ostial lobe.
Genitalia of P.d.dardanusA male genitalia B female genitalia.
In terms of immature stages, these closely related species exhibit subtle yet diagnostically significant divergences (Williams 2023, this study): The 1^st^ instar larvae: (1) P.dardanuscenea Stoll, 1790 presents a predominantly blackish-brown coloration, contrasting with much paler P.meriones larvae; (2) the W-shaped marking on the 6^th^ segment (3^rd^ abdominal segment) of P.dardanuscenea larvae is absent in P.meriones; and(3) P.meriones larvae have the 4^th^ and 5^th^ abdominal segments whitish, while in P.dardanuscenea uniformly darkish. The 2^nd^ instar larvae show no discernible interspecific differentiation, while divergences re-emerge in the 3^rd^ instar: (1) P.dardanuscenea larvae develop double rows of small blue tubercles restricted to the 3^rd^ thoracic and 1^st^, 4^th^, 6^th^ abdominal segments, whereas P.meriones exhibit continuous tubercle rows from the 2^nd^ thoracic segment to the 7^th^ abdominal segment (excluding the 3^rd^ abdominal segment); (2) The white marking on the 3^rd^ abdominal segment is expanded in P.dardanuscenea larvae, while reduced to the segment interior in P.meriones; and (3) The head capsule of P.dardanuscenea larvae is greenish, whereas that of P.meriones is glossy dark olive with pale clypeus. Later instars and pupae demonstrate morphological convergence between species; however, the whitish spot form of 5^th^ larvae and pupa characterized in P.meriones are not reported in P.dardanuscenea descriptions.
Madagascar is a hotspot for evolutionary biological research, and its fauna is widely regarded as unique. Intriguingly, the closely related species of the model of mimetic P.dardanus were also found in Madagascar (e.g., Amaurisnossima (Ward, 1870)), but P.meriones did not evolve Batesian mimicry in the same way that P.dardanus did. In addition, P.dardanusantinorii Oberthür, 1883 from Ethiopia, as well as another closely related species, P.humbloti endemic in Comoro Is, are also non-mimetic.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Benardi G (1963) Quelques aspects zoogéographiques du mimétisme chez les Lépidoptères. Proceedings of the 16th International Congress of Zoology 4: 161–166.
- 2Carpenter GDH (1948) Notes on the males of Papiliodardanus Brown (Lep.). with the definition of a new transitional race and a redescription of a P.dardanusochracea Poulton. Proceedings of the Royal Entomological Society of London.Series B, Taxonomy 17(1–2): 11–17. 10.1111/j.1365-3113.1948.tb 00876.x · doi ↗
- 3Clarke CA Sheppard PM (1960) The genetics of Papiliodardanus Brown. II. Races dardanus, polytrophus, meseres, and tibullus. Genetics 45(4): 439–456. 10.1093/genetics/45.4.439PMC 121006117247935 · doi ↗ · pubmed ↗
- 4Clarke CA Clarke LF Collins SC Gill AC Turner JR (1985) Male-like females, mimicry and transvestism in butterflies (Lepidoptera: Papilionidae).Systematic Entomology 10(3): 257–283. 10.1111/j.1365-3113.1985.tb 00137.x · doi ↗
- 5Condamine FL Sperling FAH Wahlberg N Rasplus JY Kergoat GJ (2012) What caused the latitudinal gradient of species diversity in swallowtail butterflies? Ecology Letters 15: 267–277. 10.1111/j.1461-0248.2011.01737.x 22251895 · doi ↗ · pubmed ↗
- 6Condamine FL Allio R Reboud EL Dupuis JR Toussaint EFA Mazet N Hu SJ Lewis DS Kunte K Cotton AM Sperling FAH (2023) A comprehensive phylogeny and revised taxonomy illuminate the origin and diversification of the global radiation of Papilio (Lepidoptera: Papilionidae). Molecular Phylogenetics and Evolution 183: 107758. 10.1016/j.ympev.2023.10775836907224 · doi ↗ · pubmed ↗
- 7Dupuis JR Sperling FAH (2015) Repeated reticulate evolution in North American Papiliomachaon group swallowtail butterflies. P Lo S One 10(10): e 0141882. 10.1371/journal.pone.0141882 PMC 462782826517268 · doi ↗ · pubmed ↗
- 8Ford EB (1936) The genetics of Papiliodardanus Brown (Lep).Transactions of the Royal Entomological Society of London 85(19): 435–466. 10.1111/j.1365-2311.1936.tb 00238.x · doi ↗