Pseudomonas lyxosi sp. nov., Pseudomonas arabinosi sp. nov. and Pseudomonas frigoris sp. nov., isolated from glaciers
Chan Zhao, Yu-Hua Xin, Qing Liu

TL;DR
This paper describes three new species of Pseudomonas bacteria found in glaciers on the Tibetan Plateau.
Contribution
The discovery and classification of three novel Pseudomonas species from glacier samples.
Findings
The new species are psychrotolerant, growing at temperatures as low as −2 °C.
Phylogenomic analysis confirmed their distinct status from other Pseudomonas species.
Each strain has unique fatty acid profiles and low DNA similarity to other species.
Abstract
Three Gram-stain-negative, rod-shaped, motile bacterial strains, each with a single polar flagellum, designated LB3P38T, LT1P18T and ZB1P45T, were isolated from glacier samples on the Tibetan Plateau, P.R. China. These strains grew at temperatures ranging from −2 to 32 °C (optimum 25–30 °C) and at pH values of 5.0–11.0 (optimum pH 7.0). The similarities of 16S rRNA gene sequences among these strains ranged from 99.67% to 99.93%. Phylogenetic analysis based on 16S rRNA gene sequences confirmed their affiliation with the genus Pseudomonas. Phylogenomic analysis positioned these strains in close relation to Pseudomonas svalbardensis PMCC 200367T and Pseudomonas frederiksbergensis LMG 19851T. Average nucleotide identity values and digital DNA–DNA hybridization values between these strains and other type strains of the genus Pseudomonas were below 94.6% and 63.4%, respectively. The…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
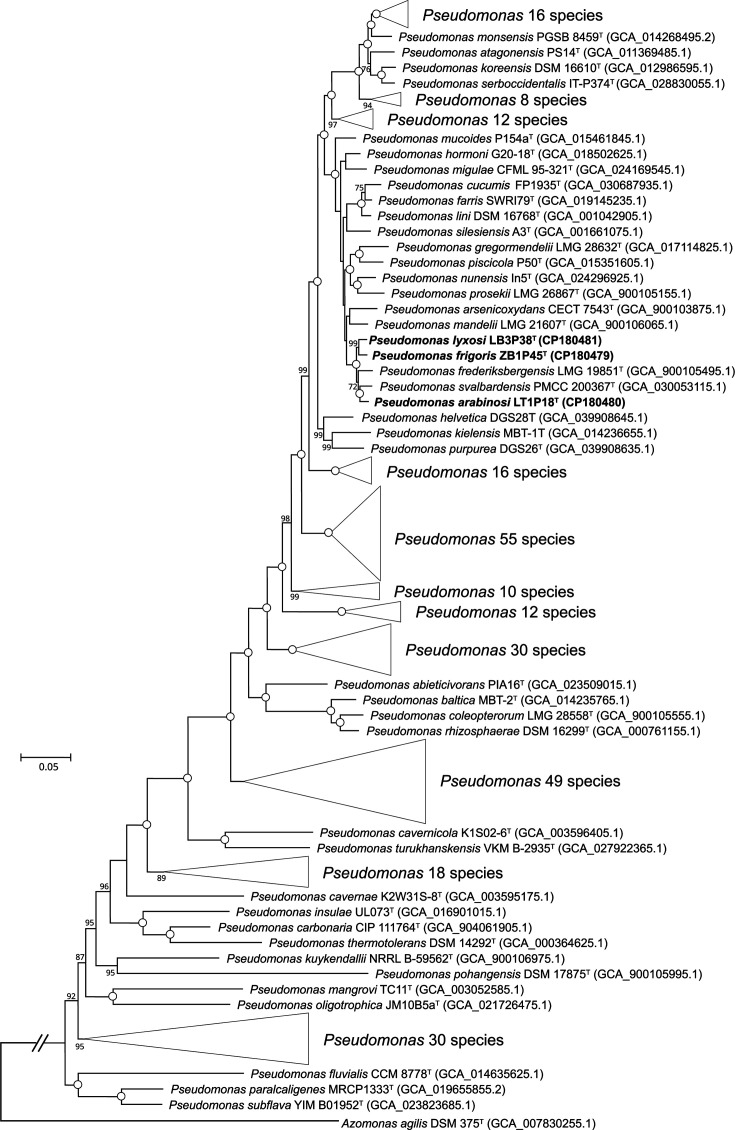 Fig. 1
Fig. 1| Characteristic | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Temperature range for growth (℃) | −2–32 | −2–32 | −2–32 | 4–30* |
| NaCl range for growth (%, w/v) | 0–5.0 | 0–5 | 0–4 | 0–7* |
| pH range for growth | 5–11 | 5–11 | 5–11 | 4–9.5* |
| Reduction of nitrates to nitrogen | − | + | − | + |
| Aesculin hydrolysis | − | − | − | + |
| Gelatin hydrolysis | + | + | − | + |
|
| ||||
| Trypsin | − | + | + | + |
| α-Chymotrypsin | − | − | + | + |
| Urease | + | − | + | + |
| Tryptophan deaminase | − | − | − | + |
|
| ||||
| | − | + | − | − |
| Sorbitol | − | − | − | + |
| | + | + | + | − |
| Esculin | − | − | − | + |
| Salicin | − | − | − | + |
| D-Cellobiose | − | − | − | + |
| D-Maltose | − | − | − | + |
| D-Melibiose | − | − | − | + |
| D-Trehalose | + | + | − | + |
| Starch | − | − | − | + |
| Glycogen | − | − | − | + |
| | + | − | − | − |
| | − | − | − | + |
| | − | + | − | + |
| 5-Ketogluconate | − | − | − | + |
| Fatty acid | 1 | 2 | 3 | 4 | 5* |
|---|---|---|---|---|---|
|
| |||||
| C12:0 | 3.1 | 2.9 | 2.7 | 5.23 | 4.7 |
| C14:0 | – | – | – |
|
|
| C16:0 | 28.9 | 29.0 | 31.6 | 32.14 | 32.0 |
| C18:0 |
|
| 1.7 |
|
|
| C17:0 cyclo | 10.0 | 17.0 | 16.2 |
| 19.7 |
|
| |||||
| C14:1
| – | – | – |
| – |
|
| |||||
| | – | – | – | 1.13 | – |
| | – |
|
|
| – |
|
| |||||
| C10:0 3-OH | 6.8 | 5.9 | 4.0 | – | 3.7 |
| C12:0 2-OH | 4.9 | 4.7 | 4.2 | 3.32 | – |
| C12:0 3-OH | 7.0 | 5.3 | 3.8 | – | 3.3 |
| C12:1 3-OH | 4.7 | – |
| – |
|
|
| |||||
| 3 | 22.3 | 17.1 | 19.2 | 38.63 | 23.2 |
| 8 | 8.5 | 13.7 | 13.0 | 9.34 | 7.0 |
- —Beijing Municipal Science & Technology Project, China
- —Biological Resources Programme, Chinese Academy of Sciences
- —http://dx.doi.org/10.13039/501100001809 National Natural Science Foundation of China
- —Strategic Priority Research Program of the Chinese Academy of Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Microbial Community Ecology and Physiology · Probiotics and Fermented Foods
The genus Pseudomonas, belonging to the family Pseudomonadaceae, was originally described by Migula [1], with Pseudomonas aeruginosa as the type species. Most Pseudomonas strains are Gram-negative, aerobic and rod-shaped. Pseudomonas species exhibit extensive genetic diversity and remarkable metabolic versatility [23]. At the time of writing, the genus Pseudomonas includes over 350 species with validly published names [4]. Hesse et al. [5] established a genomic framework for Pseudomonas evolutionary history, identifying 13 evolutionary groups by comparing genomes from type strains of 163 species and 3 subspecies. Pseudomonas is widely distributed across diverse habitats, including plants [6], soil [7], freshwater [8], marine environments [9] and glaciers [10]. Certain species, such as P. aeruginosa and Pseudomonas syringae, are known as human and plant pathogens, respectively [11]. Studies have shown that Pseudomonas is one of the most dominant bacterial genera present in glacial soils and snow [10]. In this study, three Pseudomonas strains, LB3P38^T^, LT1P18^T^ and ZB1P45^T^, were isolated from glacier samples in P.R. China and characterized using a polyphasic taxonomic approach. The results indicated that these strains represent three novel species, proposed here as Pseudomonas lyxosi sp. nov., Pseudomonas arabinosi sp. nov. and Pseudomonas frigoris sp. nov., respectively.
Isolation and ecology
Strains LB3P38^T^ and LT1P18^T^ were isolated from ice and cryoconite samples, respectively, collected from the Laigu Glacier (29.3087826° N, 96.8186951° E), and strain ZB1P45^T^ was isolated from an ice sample from the Zepu Glacier (30.276556° N, 95.2508392°E), both on the Tibetan Plateau, P.R. China, in October 2016. The samples were homogenized, serially diluted with sterile water and plated onto peptone, yeast extract and glucose agar (PYG, 0.5% bacto peptone (Difco), 0.02% yeast extract, 0.5% glucose, 0.3% beef extract, 0.05% NaCl, 0.15% MgSO_4_·7H_2_O, pH adjusted to 7.0). Following incubation at 14 °C for 15 days, strains LB3P38^T^, LT1P18^T^ and ZB1P45^T^ were isolated and purified by repeated streaking on PYG agar. Strains were routinely cultured on Nutrient Agar (NA) at 14 °C and preserved in 10% (v/v) glycerol suspensions in a liquid nitrogen storage tank. Strain Pseudomonas svalbardensis CCTCC AB 2023225^T^, which is most closely related to the three strains obtained from the China Center for Type Culture Collection, was used as an experimental control.
16s rRNA phylogeny
Genomic DNA was extracted using the TaKaRa MiniBEST Bacteria Genomic DNA Extraction Kit Ver. 3.0 (TaKaRa, Dalian, China) following the manufacturer’s instructions. The 16S rRNA gene was amplified and sequenced using the bacterial universal primers 27F and 1492R [11]. Complete 16S rRNA gene sequences were also extracted from the genome assemblies of the three strains. These sequences were compared using the EzBioCloud database [12] and BLAST +programme V2.14 [13]. 16S rRNA gene sequences of closely related taxa were retrieved for phylogenetic analysis. Multiple sequence alignments were performed using the clustal_W program implemented in mega7 [14]. Phylogenetic trees were reconstructed using the neighbour-joining (NJ) [15] method in mega7, with genetic distances calculated using Kimura’s two-parameter model [16]. Tree topologies were evaluated with 1,000 bootstrap replicates [17].
Nearly complete 16S rRNA gene sequences of strains LB3P38^T^ (1,412 bp), LT1P18^T^ (1,328 bp) and ZB1P45^T^ (1,374 bp) were identical to the corresponding complete sequences (1,534 bp) retrieved from their genomic sequence. Sequence similarities among the three strains ranged from 99.67% to 99.93%. Blastn and EzBioCloud analyses confirmed that the three strains belong to the genus Pseudomonas. Type strains with 16S rRNA gene sequence similarities >98.6% to the three strains are listed in Table S1, available in the online Supplementary Material. Strains LB3P38^T^ and LT1P18^T^ shared the highest sequence similarity with * P. nunensis* In5T (99.66%), followed by P. silesiensis A3^T^ (99.52%), P. piscicola P50^T^ (99.51%) and P. mandelii NBRC 103147^T^ (99.45%). Strain ZB1P45^T^ showed the highest sequence similarity with P. nunensis In5^T^ (99.93%), followed by P. silesiensis A3^T^ (99.79%), * P. mandelii* NBRC 103147^T^ (99.73%), P. farris SWRI79^T^ (99.73%), P. svalbardensis S025^T^ (99.73%) and P. frederiksbergensis JAJ28^T^ (99.73%). The NJ tree showed that strains LB3P38^T^ and LT1P18^T^ formed a distinct clade, separated from other species with 99% bootstrap support, whereas strain ZB1P45^T^ clustered with P. svalbardensis S025^T^ and P. nunensis In5^T^ with less than 50% bootstrap support (Fig. S1).
Genome features
Genomic DNA was sequenced using a combination of long-read shotgun sequencing with the PacBio Sequel II (Pacific Biosciences, CA, USA) and short-read sequencing via Illumina sequencing platforms. Short-reads were generated using the Illumina Hiseq 4000 platform (Illumina, San Diego, CA, USA) with 150 bp paired-end reads according to the manufacturer’s protocols. Complete genome sequences were assembled and polished using Unicycler v0.5.1, integrating both short-reads and long-reads [18]. Genome sequence quality was evaluated based on completeness and contamination rates using CheckM2 v1.0.1 [19] and QUAST v5.2 [20]. The genomes of the three strains were annotated using Bakta version 1.7.0 [21]. Maximum-likelihood (ML) phylogenomic trees were constructed with the IQ-TREE program v2.0.7 [22] using concatenated alignment of 81 core genes extracted via the UBCG2 pipeline [23]. The best-fit nucleotide substitution model (GTR+F+I+R9) was applied, with topologies supported by 1,000 bootstrap replicates. Average nucleotide identity (ANI) values were calculated using the FastANI program [24]. Digital DNA–DNA hybridization (dDDH) values were determined using the Type (Strain) Genome Server [25].
Assembled genome information of the three strains is presented in Table S2. The genomes of strains LB3P38^T^, LT1P18^T^ and ZB1P45^T^ were circular and complete, with CheckM2 confirming 100% completeness and contamination levels of 1.38%, 1.63% and 1.33%, respectively. The G+C content was 58.9% for LB3P38^T^, 58.8% for LT1P18^T^ and 58.7% for ZB1P45^T^. To determine their taxonomic status, 295 genomic sequences of Pseudomonas species with validly published names were downloaded for phylogenomic analysis. The phylogenomic tree, constructed from core gene alignments, showed that strains LT1P18^T^, LB3P38^T^, ZB1P45^T^, P. svalbardensis PMCC 200367^T^ and P. frederiksbergensis LMG 19851^T^ formed a distinct lineage with 100% bootstrap support. Within this lineage, LB3P38^T^ and ZB1P45^T^ clustered together, while LT1P18^T^ grouped with P. svalbardensis PMCC 200367^T^ (Fig. 1). Based on the taxonomic framework established by Garrido-Sanz et al. [26] and expanded by Hesse et al. [5], these strains were classified within the P. fluorescens group, specifically the P. mandelii subgroup, as supported by their close phylogenetic relationship with P. mandelii LMG 21607^T^ in the phylogenomic tree.
Phylogenetic tree of strains LB3P38T, LT1P18T, ZB1P45T and related taxa inferred using ML algorithms in IQ-TREE based on the concatenated alignment of 81 core genes. Bootstrap values (>70%) based on 1,000 replicates are shown at the branch nodes. Open circles indicate nodes with 100% bootstrap support. Azomonas agilis DSM 375T was used as an outgroup. Scale bar, 0.05 substitutions per nucleotide position.
The ANI and dDDH values between strains LT1P18^T^, LB3P38^T^, ZB1P45^T^, P. svalbardensis PMCC 200367^T^ and P. frederiksbergensis LMG 19851^T^ were 93.0–94.6% and 52.1–63.4%, respectively (Table S3). Additionally, ANI and dDDH values between the three strains and other Pseudomonas species with validly published names, for which genome sequences were available from NCBI, were also calculated. All resulting ANI and dDDH values were below 89.4% and 37.8%, respectively. These values align with standard criteria for delineating distinct species (ANI <95–96% and dDDH <70%) [2728], supporting the classification of strains LT1P18^T^, LB3P38^T^ and ZB1P45^T^ as three novel species within the genus Pseudomonas.
Physiology and chemotaxonomy
Colony morphology of the three novel strains was observed on NA plates. Gram staining was performed as described previously [29]. Cell morphology was examined using a JEM-1400 transmission electron microscopy (JEOL Ltd., Tokyo, Japan). Motility was observed by oil-immersion phase-contrast microscopy using the hanging drop method. Growth at various temperatures (−2–42 °C) was evaluated in Nutrient Broth (NB) medium. The pH range for growth was assessed in NB medium adjusted to pH 4.0–11.0 (at 1.0 pH unit intervals), using appropriate biological buffers (0.2 M Na_2_HPO_4_/NaH_2_PO_4_·2H_2_O for pH 5–8 and 0.2 M Na_2_CO_3_/NaHCO_3_ for pH 9–10). NaCl tolerance (0–6%, w/v, at 0.5% intervals) was tested in NB medium. Anaerobic growth was evaluated in NB supplemented with 1% (w/v) KNO₃ in anaerobic culture tubes. The capacity to hydrolyse starch, casein, tributyrin and Tween 80 was tested following Smibert and Krieg using NA plate as the basal medium [30]. Catalase activity was determined using bubble production in 3% (v/v) H_2_O_2_, while cytochrome oxidase activity was assessed using 1% (w/v) tetramethyl-p-phenylenediamine. Carbon source utilization was tested using API 50 CH strips (bioMérieux) with a basal medium (0.2% (NH_4_)2_SO_4, 0.05% NaH_2_PO4·H_2_O, 0.05% K_2_HPO4, 0.02% MgSO_4_·7H_2_O and 0.01 % CaCl_2_·2H_2_O). Additional biochemical tests were performed using API 20NE, API 20E and API ZYM strips (bioMérieux) according to the manufacturer’s instructions. For cellular fatty acids analysis, cell masses were harvested from the NA plate after incubation at their optimal temperature of 28 °C for 1 day. Fatty acids were extracted following the standard protocol of the MIDI 6.0 system (MIDI Inc., Newark, DE, USA) and analysed using the 6890 N Gas Chromatograph (Agilent Technologies, Santa Clara, CA, USA) with the Sherlock Microbial Identification System version 6.0 [31].
Strains LB3P38^T^, LT1P18^T^ and ZB1P45^T^ were Gram-stain-negative, rod-shaped bacteria, each possessing a single flagellum (Fig. S2). These strains grew at −2–32 °C, pH 5.0–11.0. All strains were positive for oxidase and catalase activities. Strains LB3P38^T^ and LT1P18^T^ grew at NaCl concentrations of 0–3.5% (w/v), exhibited weak growth at 4.0 and 5.0% NaCl (w/v), but failed to grow at 6.0% NaCl (w/v). Strain ZB1P45^T^ grew at 0–3.5% NaCl (w/v), showed weak growth at 4.0% (w/v), but could not grow at 5.0% (w/v). Nitrate reduction was positive for all three strains; strains LB3P38^T^ and ZB1P45^T^ reduced nitrates to nitrites, while strain LT1P18^T^ reduced nitrates to nitrogen. Detailed phenotypic characteristics of the three novel strains are provided in the species description. Distinguished phenotypic traits between strains LB3P38^T^, LT1P18^T^, ZB1P45^T^ and the reference strain are listed in Table 1.
The cellular fatty acid profiles of three novel strains were analysed and compared with those of the reference strains P. svalbardensis CCTCC AB 2023225^T^ and P. frederiksbergensis LMG 19851^T^, as shown in Table 2. All four strains contained C_16:0_ and summed feature 3 (C_16:1_ ω7c and/or C_16:1_ ω6c) as their predominant fatty acid components. However, distinct variations were observed in the proportions of specific fatty acids across the strains. The novel strains and P. frederiksbergensis LMG 19851^T^ displayed significantly higher levels of C_17:0_ cyclo, ranging from 10.0% to 19.7%, compared with P. svalbardensis CCTCC AB 2023225^T^ (<1.0%). Additionally, hydroxylated fatty acids were more abundant in the three novel strains than in P. svalbardensis CCTCC AB 2023225^T^. In particular, C_10:0_ 2-OH and C_12:0_ 3-OH were detected at 3.8–7.0% in the novel strains but were absent in P. svalbardensis CCTCC AB 2023225^T^. Moreover, strain LB3P38^T^ was uniquely distinguished by the presence of C_12:1_ 3-OH at 4.7%, a fatty acid not observed in the other strains.
Based on their phylogenetic, physiological, chemotaxonomic and genotypic features, strains LB3P38^T^, LT1P18^T^ and ZB1P45^T^ are proposed to represent three novel species of the genus Pseudomonas, for which the names P. lyxosi (LB3P38^T^=CGMCC 1.11284^T^=JCM 37126^T^) sp. nov., P. arabinosi (LT1P18^T^=CGMCC 1.11310^T^=JCM 37131^T^) sp. nov. and P. frigoris sp. nov. (ZB1P45^T^ = CGMCC 1.23235^T^ = JCM 37141^T^) are proposed.
Description of Pseudomonas lyxosi sp. nov.
Pseudomonas lyxosi (ly.xo’si. N.L. gen. n. lyxosi, of lyxose).
Cells are Gram-stain-negative, aerobic, motile by means of a monopolar flagellum. 1.1–1.7 µm in length and 0.8–1.0 µm in width. Colonies are 1–3 mm in diameter, round and pale yellow after 1–2 days of incubation at 28 °C on NA plate. Growth occurs at −2–32 °C (optimum, 25–30 °C), pH 5.0–11.0 (optimum, pH 7.0) and 0–5.0% (w/v) NaCl (optimum, 1%). Positive for oxidase and catalase. Hydrolyse casein, gelatin, starch and tributyrin, but not esculin and Tween 80. Positive for reduction of nitrates to nitrites, citrate utilization, urease, Voges–Proskauer test, arginine dihydrolase, lysine decarboxylase, ornithine decarboxylase, alkaline phosphatase, esterase (C4), esterase lipase (C8), lipase (C14), leucine arylamidase, valine arylamidase, naphthol-AS-BI-phosphohydrolase and acid phosphatase. Negative for H_2_S production, indole production, fermentation of glucose, β-galactosidase, cystine arylamidase, trypsin, α-chymotrypsin, α-galactosidase, β-glucuronidase, α-glucosidase, β-glucosidase, N-acetyl-β-glucosaminidase, α-mannosidase and α-fucosidase. Acids are produced from d-glucose, l-rhamnose, sucrose, melibiose, amygdalin and l-arabinose. No acids are produced from d-manitol, inositol and sorbitol. Utilizes the following carbohydrate as the sole carbon source: glycerol, l-arabinose, d-ribose, d-galactose, d-glucose, d-fructose, d-mannitol, inositol, d-mannitol, N-acetylglucosamine, sucrose, trehalose, d-lyxose, d-arabitol, gluconate and 2-ketogluconate. Cannot utilize the following carbohydrates: erythritol, d-arabinose, d-xylose, l-xylose, d-adonitol, methyl-β-d-xylopyranoside, l-sorbose, l-rhamnose, dulcitol, d-sorbitol, methyl-α-d-mannopyranoside, methyl-α-d-glucopyranoside, amygdalin, arbutin, esculin, salicin, cellobiose, maltose, lactose, melibiose, inulin, melezitose, raffinose, starch, glycogen, xylitol, gentiobiose, turanose, d-tagatose, d-fucose, l-fucose, l-arabitol and 5-ketogluconate. The major cellular fatty acids are C_16:0_, summed feature 3 (C_16:1_ ω7c and/or C_16:1_ ω6c) and C_17:0_ cyclo. The genomic DNA G+C content is 58.9 mol%.
The type strain LB3P38^T^ (=CGMCC 1.11284^T^=JCM 37126^T^) was isolated from an ice sample collected from the Laigu Glacier on the Tibetan Plateau, P.R. China. The GenBank accession no. for the 16S rRNA gene sequence reported in this paper is PQ844665. The genome sequence has been deposited at DDBJ/ENA/GenBank under the accession no. CP180481.
Description of Pseudomonas arabinosi sp. nov.
Pseudomonas arabinosi (a.ra.bi.no’si. N.L. gen. n. arabinosi, pertaining to arabinose.)
Cells are Gram-stain-negative, aerobic, motile by means of a monopolar flagellum. 1.7–2.4 µm in length and 0.9–1.1 µm in width. Colonies are 1–2 mm in diameter, round and pale yellow after 1–2 days of incubation at 28 °C on NA plate. Growth occurs at −2–32 °C (optimum, 25–30 °C), pH 5.0–11.0 (optimum, pH 7.0) and 0–5.0% (w/v) NaCl (optimum, 1%). Positive for oxidase and catalase. Hydrolyse casein, gelatin, starch and tributyrin, but not esculin and Tween 80. Positive for reduction of nitrates to nitrogen, citrate utilization, Voges–Proskauer test, arginine dihydrolase, lysine decarboxylase, ornithine decarboxylase, alkaline phosphatase, esterase (C4), esterase lipase (C8), lipase (C14), leucine arylamidase, valine arylamidase, cystine arylamidase, naphthol-AS-BI-phosphohydrolase and acid phosphatase. Negative for H_2_S production, indole production, fermentation of glucose, β-galactosidase, trypsin, α-chymotrypsin, α-galactosidase, β-glucuronidase, α-glucosidase, β-glucosidase, N-acetyl-β-glucosaminidase, α-mannosidase and α-fucosidase. Acids are produced from l-rhamnose, sucrose, melibiose, amygdalin, and l-arabinose. No acids are produced from d-glucose, d-manitol, inositol and sorbitol. Utilizes the following carbohydrate as the sole carbon source: glycerol, d-arabinose, l-arabinose, d-ribose, d-galactose, d-glucose, d-fructose, d-mannose, inositol, d-mannitol, N-acetylglucosamine, sucrose, trehalose, l-fucose, d-arabitol, gluconate and 2-ketogluconate. Cannot utilize the following carbohydrates: erythritol, d-xylose, l-xylose, d-adonitol, methyl-β-d-xylopyranoside, l-sorbose, l-rhamnose, dulcitol, d-sorbitol, methyl-α-d-mannopyranoside, methyl-α-d-glucopyranoside, amygdaline, arbutin, esculin, salicin, cellobiose, maltose, lactose, melibiose, inulin, melezitose, raffinose, starch, glycogene, xylitol, gentiobiose, turanose, d-lyxose, d-tagatose, d-fucose, l-arabitol and 5-ketogluconate. The major cellular fatty acids are C_16:0_, summed feature 3 (C_16:1_ ω7c and/or C_16:1_ ω6c), C_17:0_ cyclo and summed feature 8 (C_18:1_ ω7c and/or C_18:1_ ω6c). The genomic DNA G+C content is 58.8 mol%.
The type strain LT1P18^T^ (=CGMCC 1.11310^T^=JCM 37131^T^) was isolated from a cryoconite sample collected from the Laigu Glacier on the Tibetan Plateau in the southwest of China. The GenBank accession number for the 16S rRNA gene sequence reported in this paper is PQ844666. The genome sequence has been deposited at DDBJ/ENA/GenBank under the accession no. CP180480.
Description of Pseudomonas frigoris sp. nov.
Pseudomonas frigoris (fri’go.ris. L. gen. n. frigoris, of the cold).
Cells are Gram-stain-negative, facultatively anaerobic, motile by means of a monopolar flagellum. 1.4–3.0 µm in length and 0.9–1.2 µm in width. Colonies are 1–3 mm in diameter, round and pale yellow after 1–2 days of incubation at 28 °C on NA plate. Growth occurs at −2–32 °C (optimum, 25–30 °C), at pH 5.0–11.0 (optimum, pH 7.0) and 0–4.0% (w/v) NaCl (optimum, 1%). Positive for oxidase and catalase. Hydrolyse casein, gelatin, starch and tributyrin, but not esculin and Tween 80. Positive for reduction of nitrates to nitrites, citrate utilization, urease, Voges–Proskauer test, arginine dihydrolase, lysine decarboxylase, ornithine decarboxylase, alkaline phosphatase, esterase (C4), esterase lipase (C8), lipase (C14), leucine arylamidase, valine arylamidase, cystine arylamidase, trypsin, naphthol-AS-BI-phosphohydrolase and acid phosphatase. Negative for H_2_S production, indole production, fermentation of glucose, β-galactosidase, α-chymotrypsin, α-galactosidase, β-glucuronidase, α-glucosidase, β-glucosidase, N-acetyl-β-glucosaminidase, α-mannosidase and α-fucosidase. Acids are produced from l-rhamnose and melibiose. No acids are produced from d-glucose, d-manitol, inositol, sorbitol, sucrose, amygdalin and l-arabinose. Utilizes the following carbohydrate as the sole carbon source: glycerol, l-arabinose, d-ribose, d-galactose, d-glucose, d-fructose, d-mannose, inositol, d-mannitol, N-acetylglucosamine, sucrose, d-arabitol, gluconate and 2-ketogluconate. Cannot utilize the following carbohydrates: erythritol, d-arabinose, d-xylose, l-xylose, d-adonitol, methyl-β-d-xylopyranoside, l-sorbose, l-rhamnose, dulcitol, d-sorbitol, methyl-α-d-mannopyranoside, methyl-α-d-glucopyranoside, amygdaline, arbutin, esculin, salicin, cellobiose, maltose, lactose, melibiose, trehalose, inulin, melezitose, raffinose, starch, glycogene, xylitol, gentiobiose, turanose, d-lyxose, d-tagatose, d-fucose, l-fucose, l-arabitol and 5-ketogluconate. The major cellular fatty acids are C_16:0_, summed feature 3 (comprising C_16:1_ ω7c and/or C_16:1_ ω6c), C_17:0_ cyclo and summed feature 8 (comprising C_18:1_ ω7c and/or C_18:1_ ω6c). The genomic DNA G+C content is 58.7 mol%.
The type strain (strain ZB1P45^T^=CGMCC 1.23235^T^=JCM 37141^T^) was isolated from an ice sample of Zepu Glacier in Tibet Autonomous Region, China (30.276556 N, 95.2508392 E). The GenBank accession number for the 16S rRNA gene sequence of strain ZB1P45^T^ is PQ844667. The genome sequence of strain ZB1P45^T^ has been deposited at DDBJ/ENA/GenBank under the accession no. CP180479.
Supplementary material
10.1099/ijsem.0.006799Uncited Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Migula W Über ein neues system der bakterien Arb Bakteriol Inst Karlsruhe 18941235328
- 2Lalucat J Mulet M Gomila M García-Valdés EJG Genomics in bacterial taxonomy: impact on the genus Pseudomonas Genes 20201113910.3390/genes 1102013932013079 PMC 7074058 · doi ↗ · pubmed ↗
- 3Hameed A Shahina M Lin S-Y Liu Y-C Young C-C Pseudomonas hussainii sp. nov., isolated from droppings of a seashore bird, and emended descriptions of Pseudomonas pohangensis, Pseudomonas benzenivorans and Pseudomonas segetis Int J Syst Evol Microbiol 2014642330233710.1099/ijs.0.060319-024744016 · doi ↗ · pubmed ↗
- 4Parte AC Sardà Carbasse J Meier-Kolthoff JP Reimer LC Göker M List of prokaryotic names with standing in nomenclature (LPSN) moves to the DSMZ Int J Syst Evol Microbiol 2020705607561210.1099/ijsem.0.00433232701423 PMC 7723251 · doi ↗ · pubmed ↗
- 5Hesse C Schulz F Bull CT Shaffer BT Yan Q et al Genome-based evolutionary history of Pseudomonas spp Environ Microbiol 2018202142215910.1111/1462-2920.1413029633519 · doi ↗ · pubmed ↗
- 6Qiu M Li Y Liu Q Zhang X Huang Y et al Pseudomonas machongensis sp. nov. and Stenotrophomonas capsici sp. nov., isolated from wilted pepper plants Int J Syst Evol Microbiol 20247400655010.1099/ijsem.0.00655039589767 · doi ↗ · pubmed ↗
- 7Ge H-Y Zhang Y-H Hu Y-Q Li H-R Han W et al Pseudomonas paeninsulae sp. nov. and Pseudomonas svalbardensis sp. nov., isolated from Antarctic intertidal sediment and Arctic soil, respectively Int J Syst Evol Microbiol 20247400646610.1099/ijsem.0.00646639073408 · doi ↗ · pubmed ↗
- 8Morris CE Sands DC Vinatzer BA Glaux C Guilbaud C et al The life history of the plant pathogen Pseudomonas syringae is linked to the water cycle ISME J 2008232133410.1038/ismej.2007.11318185595 · doi ↗ · pubmed ↗