RNA-seq identifies Amd1 as a regulator of hepatocyte proliferation via Skp2 during liver development and hepatocellular carcinoma progression in zebrafish
Ke Zhang, Botong Li, Zhiling Deng, Yong Dong, Yuanyuan Li, Bingyu Chen, Mao Lu, Liyan Wang, Xingdong Liu, Zhenhua Guo, Sizhou Huang

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
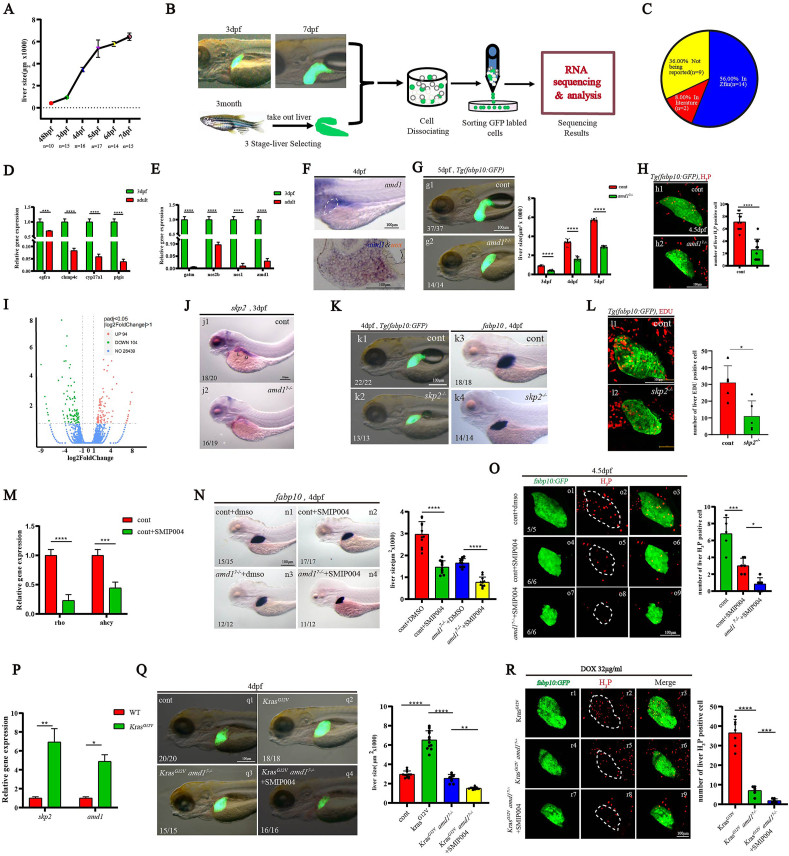 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsUbiquitin and proteasome pathways · Pancreatic function and diabetes · Cancer, Hypoxia, and Metabolism
Early studies utilized microarrays to identify liver-enriched genes, and the roles of some of these genes in liver development in mice and zebrafish were confirmed.1^,^2 However, many genes involved in liver growth regulation remained unidentified through this approach. Recently, bulk RNA sequencing or single-cell RNA sequencing has been used to study liver differentiation,3 but no study has screened genes highly expressed in hepatocytes during liver growth. Since rapid hepatocyte proliferation is a common characteristic of liver growth in early liver development and hepatocellular carcinoma (HCC) progression, identifying the genes being involved in regulating liver growth and clarifying how it works would benefit studying HCC progress.
We observed that the zebrafish liver underwent rapid growth between 2 dpf (days post fertilization) and 5 dpf (Fig. 1A; Fig. S1) and hypothesized that certain genes regulating liver growth were likely to be highly expressed in hepatocytes during this period. To identify these genes, we performed RNA sequencing on GFP-labeled hepatocytes sorted at different developmental stages (Fig. 1B): 3 dpf, 7 dpf and adult zebrafish. The results showed that 1654 genes were enriched in the hepatocytes but not in non-hepatocytes at 3 dpf (Table S1). Among the top 25 of these genes, 14 were previously reported as liver-enriched in the Zfin database, 2 had been identified in earlier publications, and 9 were novel findings (Fig. 1C; Table S3), suggesting the identification of new liver-enriched genes. Further analysis revealed that 813 of these genes were highly expressed in 3 dpf hepatocytes but not in adult hepatocytes (Table S4). This finding was validated by quantitative reverse transcription PCR analysis of four selected genes (Fig. 1D). Since the rate of liver growth significantly decreased after 5 hpf (Fig. 1A; Fig. S1), we hypothesized that the expression of genes regulating liver growth would be down-regulated in hepatocytes at 7 dpf compared with 3 dpf. Indeed, 756 of the 813 genes were down-regulated in hepatocytes at 7 dpf compared with 3 dpf (Table. S5). Interestingly, among these 756 genes, four were associated with arginine and proline metabolism, and quantitative reverse transcription PCR confirmed their higher expression in 3 dpf hepatocytes (Fig. 1E). To these four genes, amd1, the key enzyme involved in the synthesis of polyamines and embryonic stem cell self-renewal,4 its role in liver development has not been addressed. We investigated the role of amd1 in liver growth to confirm that the genes we identified could serve as candidates for regulating liver growth.Figure 1. The amd1-skp2 cascade regulates hepatocyte proliferation during liver development and HCC progression in zebrafish. (A) Comparison of liver size from 2 dpf to 7 dpf using in situ hybridization and live Tg(fabp10:GFP) transgenic embryos. Lateral view. (B) Schedule for sorting different staged hepatocytes and bulk RNA sequencing. (C) Among the top 25 genes of liver-enriched genes at 3 dpf, 56% were reported to be enriched in the embryonic liver in the Zfin database, 8% were reported in early literature, and 36% were uncharacterized. (D) Four genes were randomly selected from those enriched and highly expressed in hepatocytes at 3 dpf. Their expression levels were higher in hepatocytes at 3 dpf compared with adult hepatocytes. (E) The relative expression of gatm (0.63%), nos2b (9.7%), nos1 (1.0%), and amd1 (3.0%) in 3 dpf hepatocytes was much higher than in adult hepatocytes. (F) Single staining (amd1 as probe) and double staining (uox for fast red, amd1 for blue) at 4 dpf. Uox and amd1 were colocalized in the liver. (G) From 2 dpf to 5 dpf, liver size was smaller in amd1^7−/−^ embryos than in controls. (H) At 4.5 dpf, the number of proliferating hepatocytes stained with H3P in amd1^7−/−^ embryos was lower than in control embryos. (I) RNA sequencing data analysis showed that in amd1^7−/−^ embryos, 94 genes were up-regulated, and 104 genes were down-regulated; there was no significant difference in the expression of 28,439 genes. **(J)**In situ experiments showed that skp2 was down-regulated in the liver and gut in amd1^7−/−^ embryos at 4 dpf. (K) Liver size in skp2 mutants (k2, k4) was smaller than in controls (k1, k3). (L) The number of proliferating hepatocytes stained with EdU in skp2 mutants (l2) is smaller than in controls (l1). (M) Quantitative reverse transcription PCR results showed that the expression of rho and ahcy was significantly down-regulated in embryos treated with 80 μM SMIP004. (N) In situ experiments showed that 100% of embryos treated with 80 μM SMIP004 (Nn2) displayed smaller livers than controls (Nn1). Inhibiting skp2 activity using SMIP004 resulted in 91.6% of amd1^7−/−^ embryos (Nn4) displaying much smaller livers than controls (Nn3). (O) Cell proliferation was evaluated using H_3_P staining. After treatment with SMIP004, the number of H_3_P-stained hepatocytes was decreased compared with controls (control, n = 5; SMIP treatment, n = 6; p = 0.0004). SMIP004 treatment further decreased the number of H_3_P-stained hepatocytes in amd1^7−/−^ embryos (n = 6; p = 0.171). (P) Compared with normal liver, the expression of amd1 (5.39-fold increase; p = 0.0162) and skp2 (7.93-fold increase; p = 0.0035) was significantly increased. (Q) Dox-induced overexpression of Kras increased liver growth in the HCC model (Qq2; p ≤ 0.0001), amd1 loss of function decreased the size of the liver in the HCC model (Qq3; p ≤ 0.0001), and simultaneous inhibition of skp2 and amd1 resulted in much smaller livers in the HCC model (Qq4; p = 0.0012). (R) Compared with controls, amd1 loss of function decreased the number of H_3_P-stained hepatocytes (Rr5; p ≤ 0.0001), and simultaneous inhibition of skp2 and amd1 further decreased the number of H_3_P-stained hepatocytes (Rr8; p = 0.0005). Values are reported as mean ± standard error of the mean. ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001, ∗∗∗∗p < 0.0001. HCC, hepatocellular carcinoma; dpf, days post fertilization.Figure 1
First, in situ experiments showed that amd1 was a maternal gene and was highly expressed in the liver at 3 dpf and 4 dpf (Fig. 1F; Fig. S2A, B). The expression of amd1 in hepatocytes at 3 dpf was higher than at 4 dpf (Fig. S2C), coinciding with the most rapid liver growth occurring between 3 dpf and 4 dpf (Fig. 1A; Fig. S1). These data suggest a potential role for amd1 in liver growth. Next, we generated two amd1 mutant lines (amd1^168^ and amd1^7^; Fig. S3A–D) to investigate this possibility. Although no distinct external phenotype was observed during early development in amd1^7−/−^ or amd1^168−/−^ embryos (Fig. S3E, F), and liver specification was not disturbed (Fig. S4), the livers in these two mutant lines were smaller than those of controls from 3 dpf to 5 dpf (Fig. 1G; Fig. S5). These results indicate that amd1 mutation specifically inhibits liver growth. Furthermore, immunostaining and TUNEL assays showed that hepatocyte proliferation, but not apoptosis, was decreased in amd1^7−/−^ embryos (Fig. 1H; Fig. S6B–E). Additionally, proliferation-related markers cdk1, cdk4, chk1 and mcm5 were down-regulated in amd1 mutants (Fig. S6A). These findings demonstrate that amd1 is essential for hepatocyte proliferation.
To elucidate how amd1 regulates liver growth, we analyzed gene expression in amd1^7−/−^ embryos at 4 dpf using RNA sequencing. The results showed that 94 genes were up-regulated and 104 genes were down-regulated in amd1^7−/−^ embryos (Fig. 1I). Among the down-regulated genes, four were in the mTOR signaling pathway, including skp2 (Fig. S7A, B). Skp2, a key component of the SKP1-cullin 1-F-box (SCF) complex, primarily functions as an oncoprotein.5 Skp2 was expressed ubiquitously before 24 hpf and then restricted to the liver, gut, eyes and boundary of the hindbrain and midbrain at 3 dpf (Fig. S7C). Quantitative reverse transcription PCR (Fig. S7D) and in situ hybridization (Fig. 1J) showed that skp2 was down-regulated in amd1^7−/−^ embryos at 3 dpf, particularly in the liver and gut. These results suggest that skp2 may be required for amd1 to regulate liver growth. Next, we generated a skp2 mutant line (Fig. S8A–D) and found that both liver size and hepatocyte proliferation were reduced (Fig. 1K and L). Finally, we found that injection of skp2 mRNA could restore liver development in amd1 mutants (Fig. S8E). These data suggest that skp2 is required for liver development and is also essential for amd1 to regulate liver development.
Skp2 was ubiquitously expressed at early stages (Fig. S7C), and the expression of prox1 and hhex was decreased in skp2 mutants at 48 hpf (Fig. S8H, I). This result suggests the possibility that the liver phenotype is a secondary effect of early developmental defects in skp2 mutants. Treatment with 80 μM SMIP004 (skp2 inhibitor) reduced Skp2 activity, leading to decreased expression of the downstream genes rho and ahcy (Fig. 1M; Fig. S9A–D). To observe the direct role of skp2 during liver growth, embryos were treated with 80 μM SMIP004 from 48 hpf to 4 dpf, and liver size was analyzed (Fig. S9E, F). The data showed that skp2 inhibition resulted in smaller liver at 4 dpf (Fig. 1. Nn2; Fig. S9Ff2) and decreased hepatocyte proliferation (Fig. 1. Oo6), demonstrating that skp2 plays a vital role in liver growth. Additionally, treatment with SMIP004 also led to a much smaller liver and significantly reduced hepatocyte proliferation in amd1^7−/−^ embryos (Fig. 1. Nn4, Oo9), implying that skp2 is required for amd1 to regulate liver growth.
Since hepatocytes in developing liver and HCC are characterized by rapid cell proliferation, we generated an HCC model in zebrafish larvae (Fig. S10A–C) to examine whether amd1 and skp2 played critical roles during zebrafish HCC progression. The data showed that the expression of amd1 and skp2 was up-regulated in the sorted HCC cells (Fig. 1P), and the liver size and hepatocyte proliferation in amd1^−/−^ embryos were decreased compared with controls (Fig. 1. Qq3, Rr6). Additionally, simultaneous inhibition of amd1 and skp2 further decreased liver size and hepatocyte proliferation in the zebrafish HCC model (Fig. 1. Qq4, Rr9). These data demonstrate that the Amd1-Skp2 cascade was also involved in HCC progression in zebrafish.
In conclusion, our study identified some uncharacterized genes enriched in hepatocytes at 3 dpf. Among these genes, amd1 was identified as playing a crucial role in regulating hepatocyte proliferation via skp2 during normal liver development and HCC progression. These results imply that the identified genes may be candidates for regulating hepatocyte proliferation in normal liver development and HCC progression.
CRediT authorship contribution statement
Ke Zhang: Validation, Software, Methodology, Investigation, Formal analysis, Data curation. Botong Li: Methodology, Investigation, Data curation. Zhiling Deng: Methodology, Investigation. Yong Dong: Software, Methodology, Investigation. Yuanyuan Li: Methodology, Investigation. Bingyu Chen: Investigation. Mao Lu: Writing – review & editing, Methodology. Liyan Wang: Writing – review & editing, Methodology. Xingdong Liu: Writing – review & editing, Conceptualization. Zhenhua Guo: Writing – review & editing, Writing – original draft, Visualization, Validation, Resources, Methodology, Investigation, Formal analysis, Conceptualization. Sizhou Huang: Writing – review & editing, Writing – original draft, Supervision, Methodology, Investigation, Funding acquisition, Conceptualization.
Ethics declaration
All experimental methods and protocols were approved by Chengdu Medical College (Sichuan, China). Zebrafish were maintained following the Guidelines of the Animal Care Committee of Chengdu Medical College, Sichuan, China.
Funding
This work was supported by the 10.13039/501100001809National Natural Science Foundation of China (No. 32070805), the 10.13039/501100004829Science and Technology Department of Sichuan Province, China (No. 2024NSFSC2093); National Clinical Research Center for Child Health and Disorders, Children’s Hospital of Chongqing Medical University (NCRCCHD-2022-GP-04).
Conflict of interests
The authors declared no conflict of interests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cheng W.Guo L.Zhang Z.HNF factors form a network to regulate liver-enriched genes in zebrafish Dev Biol 294220064824961663115810.1016/j.ydbio.2006.03.018 · doi ↗ · pubmed ↗
- 2Jochheim-Richter A.Rüdrich U.Koczan D.Gene expression analysis identifies novel genes participating in early murine liver development and adult liver regeneration Differentiation 74420061671731668398710.1111/j.1432-0436.2006.00066.x · doi ↗ · pubmed ↗
- 3Yang L.Wang W.H.Qiu W.L.Guo Z.Bi E.Xu C.R.A single-cell transcriptomic analysis reveals precise pathways and regulatory mechanisms underlying hepatoblast differentiation Hepatology 6652017138714012868148410.1002/hep.29353 PMC 5650503 · doi ↗ · pubmed ↗
- 4Zhang D.Zhao T.Ang H.S.AMD 1 is essential for ESC self-renewal and is translationally down-regulated on differentiation to neural precursor cells Genes Dev 26520124614732239144910.1101/gad.182998.111PMC 3305984 · doi ↗ · pubmed ↗
- 5Cai Z.Moten A.Peng D.The Skp 2 pathway: a critical target for cancer therapy Semin Cancer Biol 67Pt 2202016333201460810.1016/j.semcancer.2020.01.013PMC 9201937 · doi ↗ · pubmed ↗