Three new species of Pleurothallis (Orchidaceae) from Costa Rica and Panama, with a note on asexual reproduction by prolification in Pleurothallidinae

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9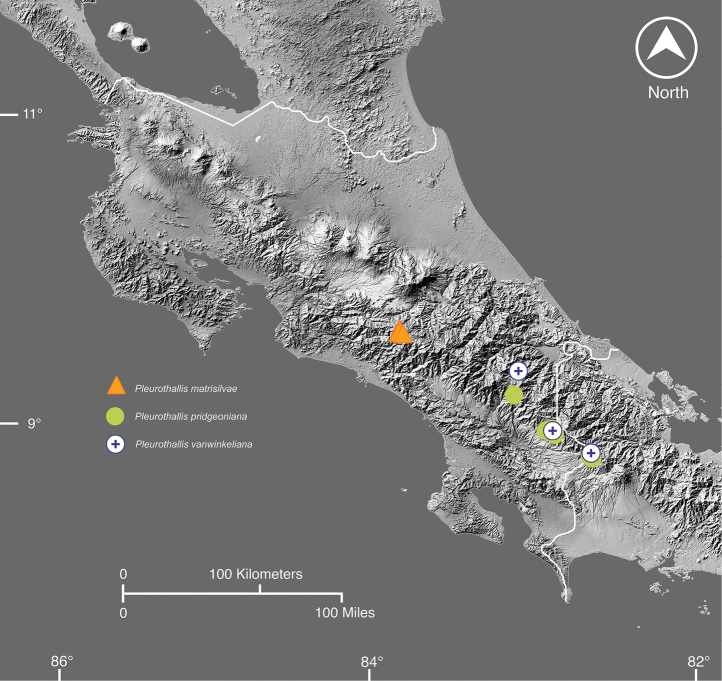 Figure 10
Figure 10Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and animal studies · Botany, Ecology, and Taxonomy Studies · Plant and Fungal Species Descriptions
Introduction
Pleurothallis R.Br. (Aiton 1813: 211), currently includes around 550 accepted species, being surpassed only by Lepanthes Sw. (Swartz 1799a: 85), Masdevallia Ruiz & Pavón (Ruiz and Pavón 1794: 122), and Stelis Sw. (Swartz 1799b: 239) as the most species-rich genus in the Pleurothallidinae (Karremans 2016; Karremans and Vieira-Uribe 2020). They are distributed from Mexico through Central America and the Antilles to the Andean and Guayanan countries of South America, where they reach their highest diversity (Luer 2005). In Costa Rica, Pleurothallis is the fourth most species-rich orchid genus with 63 species and four natural hybrids recognized by Pupulin et al. (2023). Although some highly morphologically distinctive Pleurothallis species were recently described from Costa Rica (e.g. Karremans and Bogarín 2011), most novelties are recognized when species complexes are carefully monographed, as it happened in the PleurothallisbothrosLuer (1996: 70), P.cardiothallis Rchb.f. (Reichenbach 1857: 158) and P.phyllocardia Rchb.f. (Reichenbach 1866: 97) groups (Pupulin et al. 2017a, 2017b, 2021; Karremans and Jiménez 2018). Despite thorough studies, Pupulin et al. (2023) noted that the number of Pleurothallis species in Costa Rica with a small habit and flowers, colloquially known as ‘frogs’, will likely quadruple in the near future. One such species was described as P.trigyna Pupulin (Belfort-Oconitrillo et al. 2024: 173), and here we describe three additional Pleurothallis species from Costa Rica and western Panama.
Vegetative morphology in Pleurothallis is somewhat uniform, members of this genus usually form caespitose plants with ramicauls emerging from the rhizome. But the three species treated here stand out for exhibiting a prolific growth habit. Prolification refers to the phenomenon in certain plants where the floral meristem develops a vegetative bud from the axil of a bract (Bell and Bryan 2008). In orchids, these clones, named keikis in horticulture, are usually associated with adventitious roots and can grow independently if shed or removed from the inflorescence axis, resulting in an asexual reproduction method (Rojas-Alvarado et al. 2021; Rojas-Alvarado and Karremans 2024). Prolification is frequent across Pleurothallidinae, where it often involves the formation of a ramicaul, on the apex of a previous ramicaul, occasionally forming chains (Figs 1, 2). In some Pleurothallidinae, prolification is a constitutive feature, representing the standard condition for the species or group of species. This is true for a few species in Pleurothallis and Stelis, to all species of MyoxanthusPoepp. & Endl.sect.Scandentia Luer (Luer 1992: 5), and Karma Karremans (Karremans 2023: 62), and to most species of Andinia (Luer) Luer (Luer 2000: 5), among others, which form pendent chains of superposed stems. The vast majority of pleurothallids, however, exhibit facultative prolification, occurring only occasionally, mostly under stress-induced, non-optimal, growing conditions (Rojas-Alvarado et al. 2021; Rojas-Alvarado and Karremans 2024). Most constitutively prolific Pleurothallidinae form long chains of ramicauls that lack roots and do not naturally detach from the plant (Figs 1A, E–G, K). In contrast, the prolific growth of certain Pleurothallis species produces multiple roots and ramicauls that can naturally sever from the plant with age. Strictly prolific species of Pleurothallis include P.killipii Garay (Garay 1956: 254), P.lopezii Luer & R.Escobar (Luer and Escobar 1998: 102), P.palliolata Ames (Ames 1922: 86) (Fig. 2F) and P.vinealis Luer & R.Escobar (Luer and Escobar 1986: 30). These are all high elevation species, often growing at around 2000 m, and up to 3000 m in elevation, where they are found in humid, dense, mossy vegetations. In the field, large plants with such a habit are typically found surrounded by independently growing plantlets, which arise by prolification as a means of asexual reproduction (Rojas-Alvarado and Karremans 2024).
Prolific ramicauls in species across PleurothallidinaeAZootrophionmachaqwayi (JBL-4585) BZ.vulturiceps (Pupulin 3960) CZ.gracilentus (Pupulin 5220) DLepanthopsisprolifera (JBL-458091) EKarmachaetoglossa (JBL-45847) FKarma sp. (Álvarez 1444) GMyoxanthusscandens (Rojas-Alvarado 1621) HDresslerellapilosissima (Pupulin 8070) ITrichosalpinx sp. (Bogarín 11972) JTrichosalpinxblaisdellii (Fernández 123) KAndinia sp. (CIOA-001590) LRestrepiatrichoglossa (Pupulin 7889) MLepanthes sp. (Álvarez 1466) NEchinosepalauncinata (Chinchilla 1026) OE.longipedunculata (Rojas-Alvarado 570). Photographs and digital composition by FP.
Prolific ramicauls in species of Pleurothallis. Macrophyllae-fasciculate group AP.matrisilvae (Pupulin 8951) BP.phyllocardioides (Chinchilla 4380) CP.homalantha (Pupulin 9040) DPleurothallis sp. (Fernández 250) EP.winkeliana (Chinchilla 3408) FP.palliolata (Bogarín 9101) G, HP.pridgeoniana (Chinchilla 2865). Macrophyllae-racemosae group. Ancipitae group IP.eumecocaulon (Rojas-Alvarado 1556) IP.crocodiliceps (Pupulin 5783). Pleurothallis group JP.ruscifolia (Pupulin 7013) KPleurothalliscf.ruscifolia (Smith 261) LPleurothallis sp. (Bogarín 5304) MP.rowleei (Pupulin 4352). Photographs and digital composition by FP and APK (G, H).
While preparing a monograph of the genus Pleurothallis for Costa Rica, we found three unidentified taxa with a prolific habit growing at high-elevations in the Talamanca mountain range. These novelties belong to Pleurothallissubsect.Macrophyllae-Fasciculatae [= Acroniasect.Macrophyllae-Fasciculatae (Lindl.) Luer] as defined by Luer (Luer 1988; 2005), but may not be closely related. Unfortunately, no broad phylogeny of Pleurothallis is currently available. Studies including only a small sampling fraction of the genus show that Pleurothallis is monophyletic as currently defined (Karremans 2016), but sections and subsections are not monophyletic, including the paraphyletic Pleurothallissubsect.Macrophyllae-Fasciculatae (Karremans et al. 2013; Pérez-Escobar et al. 2017; Chumová et al. 2021). How these species groups interrelate remains to be shown. Although we can only speculate about their phylogenetic relationships, we are confident the taxa described here represent previously unknown entities. Therefore, we formally describe and illustrate them, based on field-collected and cultivated specimens.
Methods
Plant collection and documentation followed the methods described by Pupulin et al. (2021). Living plants intended for this study were collected between 2010 and 2024 and cultivated at Lankester Botanical Garden (JBL, its acronym in Spanish), University of Costa Rica. At JBL plants were individually labelled and cultivated, and field data was recorded and stored in the general databases. Living plants from Costa Rica and Panama were photographed using Nikon 7100, 810, and 850 cameras, and the herbarium specimens were prepared from living plants and deposited in the liquid or dried collections at JBL and USJ (acronyms following Thiers 2025, continuously updated). Images were optimized through post-processing with Adobe Photoshop 2024, and the Lankester Composite Digital Plates (LCDP) and other comparative plates of flowers were prepared using the same software. The cited georeferenced occurrences were plotted onto a shaded relief base map using the free and open-source QGIS 3.34.
Standard characters states in Pleurothallis, such as green leaves, glabrous perianth parts, or regular and straight margins, are not mentioned expressly in the description. Terminology for the description of the inflorescences follows Rojas-Alvarado and Karremans (2024). Descriptions are based on the specimens indicated at the end of each description account. For consistency, we adopt the term ramicaul proposed by Stern and Pridgeon (1984), to refer to the leaf-bearing stem in the Pleurothallidinae. Although we acknowledge the concerns raised by Rasmussen (1985), who argued that this structure is not a true branch in the botanical sense (from Latin ramus = branch, caulis = stem), but rather a continuation of the main axis in sympodial orchids, the term ramicaul has been widely adopted in most of the taxonomic literature on Pleurothallidinae, particularly by Luer (1988, 1992, 1996, 2000, 2005) and subsequent contemporary works (Pupulin et al. 2017a, 2017b, 2021; Karremans and Jiménez 2018; Sierra-Ariza et al. 2022; Moreno et al. 2023). Additionally, we find the term useful for distinguishing different elements within the prolific growth system of Pleurothallidinae species. This should not be confused with the terms primary ramicaul and secondary ramicaul, which are used here to differentiate the elements that compose a prolific growth system. As used here, a primary ramicaul refers to the shoot that originates directly from the primary stem (the rhizome), while a secondary ramicaul refers to a stem that develops from the apical meristem of a ramicaul.
Taxonomic treatment
Pleurothallis
matrisilvae
Taxon classificationPlantaeAsparagalesOrchidaceae
Karremans, J.Gange & Pupulin sp. nov.
C0944F9F-B518-5453-9722-7835ABC5B2ED
urn:lsid:ipni.org:names:77362475-1
Type.
Costa Rica. • Cartago: El Guarco, San Isidro, Madreselva, Tres de Junio, Carretera Interamericana Sur, 9°40'31"N, 83°53'33"W, 2530 m, bosque pluvial montano, en bosque secundario de robles a orillas de la carretera, 5 March 2022, fl. in cult. 8 March 2022, A.P. Karremans & J. Gange 9036 (holotype: JBL-spirit, M0058!; Fig. 3).
Pleurothallismatrisilvae Karremans, J.Gange & Pupulin A habit, showing some ramicauls bearing flowers and others bearing new growths B single leaf with flower C flower D dissected perianth (dorsal sepal, two lateral petals, lateral sepals fused into a synsepal and two views of the lip) E column and lip, lateral view F column and ovary, lateral and ventral views G two views of the anther cap and pollinarium. LCDP prepared by APK based on Karremans 9036.
Diagnosis.
The flower is superficially similar to that of Pleurothallisbothros, however the new species may be distinguished by the prolific habit (vs. non-prolific), ramicauls linear, very thin throughout (vs. ramicauls subclavate, notably thickened apically), bearing 1–4 open flowers (vs. many, usually +10, simultaneous flowers), the longer flower segments (dorsal sepal 7.4–7.6 vs. 6.5–6.6 mm, lateral sepals 6.6 vs. 6.0 mm, petals 5.6–5.7 vs. 3.8–4.0 mm, lip 4.1–4.3 vs. 3.1–3.5 mm), the lanceolate petals (vs. oblong) and the black flecks on the pedicel, ovary, and external surface of the sepals (vs. no flecks).
Description.
Epiphytic, caespitose, constitutively prolific, suberect to erect herbs, up to 30 cm tall. Roots flexuous, thin, ca. 1 mm in diameter, densely spaced, appearing fasciculate. Ramicauls erect to suberect, slender, up to 28 cm long, covered by tubular sheaths close to the base, tightly adpressed, up to 5 cm long. Leaves spreading, glossy, dark green, coriaceous, sessile, ovate-cordate with downturned margins, acuminate, blades of the primary ramicaul 4.0–7.5 × 2.0–3.8 cm. Inflorescences persistent, forming few to several successive multi-flowered coflorescences, each producing a single open flower, up to 4 different coflorescences bearing simultaneous single flowers, typically 1 or 2, subtended by a nearly prostrate or suberect spathe which appears deeply torn during and after anthesis, ca. 1 cm long; pedicels terete, pale gray-yellow, with irregular black flecks. Ovary clavate, 4.5–4.7 mm long, gray-green and suffused with yellow, similarly marked with black. Flowers spreading, yellow. Dorsal sepal elliptic-ovate, slightly concave, obtuse, 3-veined, with irregular black flecks on its adaxial surface, 7.4–7.6 × 4.5 mm, apiculate. Lateral sepals connate in an oblong, slightly concave synsepal, obtuse, 4-veined, 6.6 × 5.8–5.9 mm. Petals linear-lanceolate, oblique, acute, 1-veined, 5.6–5.7 × 1.0–1.1 mm. Lip triangular-ovate, resting on the synsepal, 4.1–4.3 × 2.7–2.8, margins raised, apex slightly recurved, acute, glenion a deep cavity between the raised lateral margins. Column straight, transversely subrectangular, 1.7–1.9 mm long, apically 2.1 mm wide, with a ca. 1 mm column foot, the anther and stigma apical. Anther cap ovate, cucullate, obtuse at the base, bilobed apically, 2-celled. Pollinarium composed of 2, narrowly ovate-pyriform pollinia, connected to a liquid, drop-like viscidium. Fruits and seeds unknown. This description is based on A.P. Karremans & J. Gange 9036, D.E. Mora s.n., and D. Bogarín et al. 13652.
Additional specimens examined.
Costa Rica. • Cartago: Cordillera de Talamanca, alt. 2400 m, D.E. Mora s.n. (USJ!). Cartago: El Guarco, Cañón, Bajo Gloria, Centro Ecoturístico Los Robles, inicio del sendero Danta, 9°42'14.97"N, 83°54'55.96"W, 2332 m, epífitas en árboles alrededor de los senderos, bosque muy húmedo montano bajo, 12 February 2022, fl. in cult. 19 April 2022, D. Bogarín 13652, S. Bogarín, M. Bonilla & O.A. Pérez-Escobar (JBL-Spirit, E1544!; Fig. 4B). • San José-Cartago: Dota-El Guarco, Copey-Cañon, ca. 100 m deviation point in front of the Liceo Rural of Cañon, slopes of Cerro Artieda, 9°40'52.44"N, 83°55'10.04” W, 2545 m, epiphytic on scattered trees in pastures and remnants of secondary woods, 3 March 2022, fl. in cult. 21 April 2022, F. Pupulin 8951 & D. Bogarín (JBL-spirit, E1545!).
Pleurothallismatrisilvae Karremans, J.Gange & Pupulin composite A the florally similar P.bothros (JBL-sn) B bearing a single open flower and several developing buds (Bogarín 13652) C epiphytic plant in its natural habitat in Madreselva on the central Talamanca range, showing the prolific growth and asexual production of new units. Photographs by APK.
Eponymy.
From the Latin “matrisilva”, mother-forest, honoring the locality Madreselva, where the type specimen was collected.
Phenology.
Flowering has been recorded at least from November to April, which approximately corresponds to the end of the rainy season and the first months of the dry season in Costa Rica.
Distribution and ecology.
Currently known only from central Costa Rica, where the species is found growing either epiphytically, mainly on oaks, or terrestrially, on the leaf mulch rich ground, in montane or cloud forests at high elevations around 2300–2550 m (Fig. 4C). Pleurothallismatrisilvae is not a particularly rare species, but all currently known specimens were collected in the same general area on the Cordillera de Talamanca in the province of Cartago.
Notes.
Contrary to the other species described here, Pleurothallismatrisilvae is typically prolific when growing as an epiphyte, while it is mostly non-prolific when found growing on top of organic matter on the ground. The new species is morphologically similar to P.bothros and its relatives. It, however, grows at higher elevations and is distinguished by the often-prolific habit, the thin ramicauls and especially by having mainly one or two flowers open at once, which is unlike all other species in the P.bothros group (Karremans and Jiménez 2018). It is most similar to Pleurothallisbothros, which is endemic to the Cordillera de Guanacaste and Tilarán in the northern provinces of Alajuela, Guanacaste, and Puntarenas in Costa Rica, where it grows at mid elevations between 750 and 1750 m. The other two members of the P.bothros group, P.hawkingii Karremans & J.E.Jiménez and P.vide-vallis Karremans & J.E.Jiménez can be easily separated from P.matrisilvae by the non-prolific habit (vs. prolific), three or more flowers opening simultaneously (vs. typically one or two), their pale yellow flowers suffused with a pink (vs. solid yellow), the broad, oblique petals (vs. narrow, straight), and the proportionally much larger lip. Both are only known from the Cordillera de Guanacaste in northern Costa Rica (Karremans and Jiménez 2018).
Pleurothallis
pridgeoniana
Taxon classificationPlantaeAsparagalesOrchidaceae
Karremans, Bogarín & Pupulin sp. nov.
A8E88CED-A6B4-5C96-82E1-EF16E883DE23
urn:lsid:ipni.org:names:77362476-1
Type.
Costa Rica. Puntarenas: Buenos Aires, Buenos Aires, camino a Cerros Utyum, 9°18'04.93"N, 83°12'51.59"W, 2157 m, bosque pluvial montano bajo, epífitas en bosque secundario, 16 January 2017, fl. in cult. 20 December 2017, A.P. Karremans, D. Bogarín, M. Cedeño, I. Chinchilla, M. Díaz, E. Kaes, P. Lehmann & O. Zúñiga 7600 (holotype: JBL-spirit, E1514!; isotype, USJ!; Fig. 5).
Pleurothallispridgeoniana Karremans, Bogarín & Pupulin A habit showing the typical prolific vegetative growth B prolific growth featuring a single flower on a secondary ramicaul C flower D dissected perianth (dorsal sepal, two lateral petals, two lateral sepals fused into a synsepal and a view of the lip) E lip F column and lip, lateral view G column, lip and ovary, lateral view H anther cap and pollinarium. LCDP prepared by APK based on Karremans 7600 (A, C–H) and Bogarin 8632 (B).
Diagnosis.
Vegetatively similar to P.vinealis, but distinguished by the significantly shorter plants, up to 30 cm long (vs. exceeding 1 m), the shorter ramicauls 7.5–26.0 cm (vs. up to 40 cm long), the significantly smaller dorsal sepal (6.8–7.0 × 3.3–3.4 mm vs. 11–14 × 4.5–5.5 mm), synsepal (5.5–6.5 × 4.1–4.2 mm vs. 11–14 × 4.5–5.5 mm), and lip (2.4–2.6 × 1.6–1.7 mm vs. 5.5–6.5 × 3 mm). The flowers are transparent yellow, with a rose to purple suffusion (vs. brown), the lip is triangular-ovate (vs. oblong) the margins being shortly glandular (vs. denticulate), slightly raised but lacking a central sulcus (vs. prominently sulcate).
Description.
Epiphytic, caespitose, strictly prolific, erect to suberect herb, up to 30 cm long. Roots flexuous, thin, ca. 1 mm in diameter, densely spaced, appearing fasciculate. Primary ramicauls erect to suberect, slender, 7.5–26.0 cm long, with 2 basal tubular, tightly adpressed, papyraceous sheaths, up to 2.0–6.0 cm long. Secondary ramicauls 1.5–8.0 cm long, produced profusely from the floral meristem on the apex of prior ramicauls, being able to produce clumps and chains of multiple ramicauls before severing naturally. Leaves spreading to pendent, coriaceous, sessile, narrowly ovate to cordate, acute, blades of primary ramicauls 4.0–7.7 × 1.8–5.0 cm, blades on secondary ramicauls 2.5–5.2 × 1.0–2.5 cm. Inflorescence persistent, forming successive multi-flowered coflorescences with a single open flower, subtended by a nearly prostrate or suberect spathe which appears deeply torn over time, ca. 1 cm long; pedicels cylindrical. Ovary slightly clavate, ca. 3 mm long, green, with a few black dots and occasional crystals. Flowers spreading, transparent yellow, with a rose to purple suffusion of varying intensity, especially observed on the petals, lip, column, and anther cap. Dorsal sepal narrowly ovate to elliptic, acute, 3-veined, 6.8–7.0 × 3.3–3.4 mm. Lateral sepals connate in an ovate synsepal, acute, 4-veined, 5.5–6.5 × 4.1–4.2 mm. Petals naturally drooping, linear-lanceolate, oblique, acute, 1-veined, 5.1–5.4 × 0.6–0.7 mm, glandular, with the margins erose. Lip triangular-ovate, resting on the synsepal, 2.4–2.6 × 1.6–1.7 mm, blade glandular-papillose, margins minutely glandular, slightly raised, acute, shortly apiculate, with a pair of shoulder-like basal lobes. Glenion a shallow, oblong cavity between raised lateral margins. Column straight, transversely suboblong, ca. 1.5 mm long, with a short column foot, the anther and stigma apical. Anther cap ovate, cucullate, obtuse, 2-celled. Pollinarium composed of 2 narrowly ovate-pyriform pollinia connected to a liquid, drop-like viscidium. Fruits and seeds unknown. This description is based on A.P. Karremans et al. 7600, A.P. Karremans et al. 9135, D. Bogarín et al. 8632, D. Bogarín et al. 12131 and I. Chinchilla et al. 2865.
Additional specimens examined.
Costa Rica-Panama. • Puntarenas-Chiriquí: Coto Brus-Renacimiento, línea fronteriza entre Cerro Quijada del Diablo y Cerro Pando, entre mojones N.336–338, 8°54'51.9"N, 82°43'59.13"W, 2205 m, bosque muy húmedo premontano, epífita en bosque primario, “in itinere per limitem Costa Rica et Panama inter montis Quijada del Diablo et montis Pando”, 19 April 2011, fl. in cult. 14 March 2022, D. Bogarín, D. Jiménez & A.P. Karremans 8632 (JBL-spirit, E1542!, Fig. 5B). Costa Rica-Panama. • Puntarenas-Chiriquí: Coto Brus-Renacimiento, línea fronteriza entre Cerro Quijada del Diablo y Cerro Pando, entre mojones N.336–338, 8°54'51.9"N, 82°43'59.13"W, 2205 m, bosque muy húmedo premontano, epífita en bosque primario, “in itinere per limitem Costa Rica et Panama inter montis Quijada del Diablo et montis Pando”, 19 April 2011 fl. in cult. 8 October 2021, D. Bogarín 8637, D. Jiménez & A.P. Karremans (JBL-spirit, A0558!). Costa Rica. • Puntarenas: Coto Brus, Sabalito, Zona Protectora Las Tablas, 13 km al noreste de Lucha, Sitio Coto Brus, entre Río Surá y Quebrada Sutú, Finca de Miguel Sandí, 8°56'46.1"N, 82°44'30.9"W, 1778 m, bosque pluvial montano bajo, epífitas en potreros arbolados, 6 June 2010, A.P. Karremans 2829 & D. Bogarín (Fig. 6I). • Puntarenas: Buenos Aires, Buenos Aires, camino a Cerros Utyum, 9°18'04.93"N, 83°12'51.59"W, 2157 m, bosque pluvial montano bajo, epífitas en bosque secundario, 16 January 2017, D. Bogarín, M. Cedeño, I. Chinchilla, M. Díaz, E. Kaes, A.P. Karremans, P. Lehmann & O. Zúñiga 12131 [JBL-spirit, A0375! (fl. in cult. 9 May 2019), A0877! (fl. in cult. 21 March 2023)]. • Puntarenas: Buenos Aires, Buenos Aires. Olán, en el sendero de la Transutyum, después del arbolado, 9°17'56.67"N, 83°12'54.12"W, 2107 m, bosque pluvial montano bajo, bosque primario, epífita, a media luz, 16 January 2017, I. Chinchilla, D. Bogarín, A.P. Karremans, M. Díaz-Morales, M. Cedeño & E. Kaes 2865 [JBL-spirit, A0187! (fl. in cult. 19 December 2018), K0154! (fl. in cult. 18 agosto 2021), E1543! (fl. in cult. 14 marzo 2022), Fig. 6R]. • Puntarenas: Buenos Aires, Potrero Grande, Altamira, Parque Internacional La Amistad, Sector Altamira, sendero al Valle del Silencio, base del Cerro Hoffmann, 9°05'05"N, 82°58'42"W, 2450 m, 15 May 2022, A.P. Karremans, I. Chinchilla, L. Oses, G. Rojas-Alvarado & F. Vargas Acuña 9135 [JBL-spirit, A0934! (fl. in cult. 25 July 2024), A0937! (fl. in cult. 25 July 2023), K0412! (fl. in cult. 3 April 2024)]. • Puntarenas: Coto Brus, Sabalito, Zona Protectora Las Tablas, 13 km al noreste de Lucha, Sitio Coto Brus, entre Río Surá y Quebrada Sutú, Finca de Miguel Sandí, 8°56'46.1"N, 82°44'30.9"W, 1778 m, bosque pluvial montano bajo, epífitas en potreros arbolados, 6 June 2010, D. Bogarín & A.P. Karremans 7718 (Fig. 6E, H, N). Same locality and date, D. Bogarín 7742 & A.P. Karremans (Fig. 6B). Same locality and date, D. Bogarín 7743 & A.P. Karremans (Fig. 6D). • Límite entre Limón y Puntarenas: Talamanca-Buenos Aires, Bratsi-Potrero Grande, Parque Internacional La Amistad, Sector Altamira, sendero al Valle del Silencio, Cerro Hoffman, sobre la divisoria de aguas, 9°05'38.2"N, 82°58'37.73"W, 2553 m, bosque pluvial montano, 14 August 2012, fl. in cult. 7 February 2019, D. Bogarín 9806, M. Fernández, J. Godínez, A.P. Karremans, J. Kruizinga & C. Smith (JBL-spirit, A0216!, Fig. 6F, O). • Puntarenas: Coto Brus, Sabalito, Zona Protectora Las Tablas, 13 km al noreste de Lucha, Sitio Coto Brus, 8°56'46.1"N, 82°44'30.9"W, 1778 m, finca “El Capricho” de Miguel Sandí, principalmente en árboles de Quercus en las lomas y potreros al margen del río Sutú, bosque muy húmedo premontano, 6 October 2010, M. Fernández 382, R.L. Dressler, D. Bogarín & F. Pupulin [JBL-spirit, D5618!, A0209! (fl. in cult. 5 February 2019), Fig. 6G, L]. Same locality and date, M. Fernández 385, R.L. Dressler, D. Bogarín & F. Pupulin (Fig. 6A). • Puntarenas: Buenos Aires, Buenos Aires, Olán, camino a los cerros Utyúm en bosque maduro, 9°18'04.54"N, 83°12'48.72"W, 2129 m, bosque pluvial montano bajo, 16 January 2017, M. Díaz 305, D. Bogarín, M. Cedeño, I. Chinchilla, A.P. Karremans, P. Lehmann y O. Zúñiga [JBL-spirit, A0560! (fl. in cult. 8 October 2021), A0575! (fl. in cult. 3 November 2021), Fig. 6S–T]. • Puntarenas: Buenos Aires, Potrero Grande, Altamira, Parque Internacional La Amistad, Sector Altamira,, sendero al Valle del Silencio, ca. 1 km después de Cerro Quemado, 9°04'38.7"N, 82°58'37.4"W, 2284 m, bosque pluvial montano, 14 August 2012, A.P. Karremans 5685, D. Bogarín, M. Fernández, J. Godínez, J. Kruizinga & C.M. Smith (Fig. 6M). • Puntarenas: Coto Brus, Sabalito, Zona Protectora Las Tablas, 13 km NE of Lucha, Sitio Coto Brus, finca Sandí “El Capricho”, 8°56'46.1"N, 82°44'30.9"W, 1778 m, epiphytic, mostly on Quercus sp. in pastures and along the river Sutú, wet premontane forest, 6 October 2010, F. Pupulin 7887, D. Bogarín, R.L. Dressler & M. Fernández (Fig. 6Q). Same locality and date, fl. in cult. 6 December 2012, F. Pupulin et al. 7910 (JBL-spirit, D5640!, Fig. 6K). Same locality and date, F. Pupulin et al. 7893 (JBL-spirit, Fig. 6J). • Puntarenas: Coto Brus, Sabalito, Finca Las Tinieblas, propiedad de Billen Gamboa, filas hacia el norte del potrero principal. 8°55'25.6"N, 82°44'55.1"W, altitud 1975 m. Bosques maduros dominados por Quercus spp. 10 December 2023, fl. in cult. 9 September 2024, L. Álvarez 1404, B. Gamboa, M. Mata-Quirós, G. Ramírez, F. Rodríguez, J. H. Flores (JBL-spirit!).
Floral variation in PleurothallispridgeonianaAFernández 385BBogarin 7742CKarremans s.n.DBogarín 7743EBogarín 7718FBogarín 9806GFernández 382HBogarin 7718IKarremans 2829JPupulin 7893KPupulin 7910LFernández 382MKarremans 5685NBogarin 7718OBogarin 9806PKarremans s.n. QPupulin 7887RChinchilla 5865S, TDiaz 305. Photographs by the authors.
Eponymy.
Honoring the English botanist Alec M. Pridgeon, renowned worldwide for his seminal research on orchid anatomy and phylogenetics, including Pleurothallidinae. A founding editor of the journal Lindleyana and main editor of the monumental series of books Genera Orchidacearum, which laid the foundation for the contemporary understanding of orchid relationships.
Phenology.
Plants in cultivation have been recorded in flower virtually year-round, with flowering peaks in November and February, coinciding in Costa Rica with the end of the rainy season and the beginning of the dry season.
Distribution and ecology.
Currently known only from southern Costa Rica and western Panama, where plants grow either epiphytically or terrestrially on organic matter in oak forests at high elevations, around 1800–2550 m. Pleurothallispridgeoniana is locally abundant at high elevations on the Cordillera de Talamanca, in the Puntarenas (Costa Rica) and Chiriquí (Panama) provinces respectively. This notoriously prolific species is often found forming large pending mats or clumps in primary and mature forests.
Notes.
Pleurothallispridgeoniana is easily recognized by the large bushy habit derived from constitutive prolification, in which each ramicaul eventually produces vegetative growths apically, thus originating chains of multiple ramicauls (Fig. 7). The new species seems to be vegetatively most similar to P.vinealis from Colombia and Ecuador, which Luer (2005) distinguished by its unique vine-like habit, with prolific ramicauls that twine and branch, apparently attaining more than a meter in length. From P.vinealis, P.pridgeoniana can be distinguished by the significantly shorter plants, up to 30 cm long (vs. exceeding a meter), the shorter ramicauls 7.5–26.0 cm (vs. up to 40 cm long), the significantly smaller flowers, dorsal sepal 6.8–7.0 × 3.3–3.4 mm (vs. 11–14 × 4.5–5.5 mm), synsepal 5.5–6.5 × 4.1–4.2 mm (vs. 11–14 × 4.5–5.5 mm), lip triangular-ovate (vs. oblong), 2.4–2.6 × 1.6–1.7 mm (vs. 5.5–6.5 × 3 mm). The flowers are transparent yellow, with a rose to purple suffusion (vs. brown), the lip is triangular-ovate (vs. oblong) the margins glandular (vs. denticulate), slightly raised but lacking a central sulcus (vs. notably sulcate).
Pleurothallispridgeoniana Karremans, Bogarín & Pupulin in its natural habitat in Valle del Silencio on the southern Talamanca range in Costa Rica, close to the border with Panama A general overview of the prolific growth habit and asexual production of new units B detail of a flowering plant. Photographs by APK.
Florally, the non-prolific Pleurothallisapplanata Luer & Dalström and P.undulata Poepp. & Endl. (following Luer 2005) are reminiscent of P.pridgeoniana. The new species can be distinguished from the former, a species from Ecuador and Peru, by longer ramicauls (up to 26 vs. 12 cm), the smaller flowers, with shorter sepals (5.5–7.0 vs. 9.5–10.0 mm long), narrower (0.6–0.7 vs. 1 mm long), glandular petals, with erose margins (vs. smooth, entire), the shorter and narrower (2.4–2.6 × 1.6–1.7 vs. 4.5 × 2.2 mm) lip, shortly apiculate (vs. obtuse), and erect, subrectangular column, (vs. dorsally compressed on a plane with the lip). From the latter, endemic to Peru, it can be distinguished by the shorter (5.5–6.5 vs. 6–9 mm) synsepal, naturally drooping petals (vs. horizontal to elevated), and the shorter lip (2.4–2.6 vs. 3.0–4.0 mm), with a pair of shoulder-like basal lobes (vs. lip unlobed).
Pleurothallis
winkeliana
Taxon classificationPlantaeAsparagalesOrchidaceae
Karremans, Bogarín & Pupulin sp. nov.
07227E51-075C-56C5-99C4-C8EC5704B31E
urn:lsid:ipni.org:names:77362477-1
Type.
Costa Rica. • Puntarenas: Coto Brus, Sabalito, Zona Protectora Las Tablas, 13 km al noreste de Lucha, Sitio Coto Brus, entre Río Surá y Quebrada Sutú, Finca de Miguel Sandí, 8°56'46.1"N, 82°44'30.9"W, 1778 m, bosque pluvial montano bajo, epífita en potreros arbolados, 20 April 2012, fl. in cult. 25 June 2012, A.P. Karremans & J. Geml 5403 (holotype: JBL-spirit, E0966!; Figs 8, 9A–E).
Pleurothalliswinkeliana Karremans, Bogarín & Pupulin A habit showing some prolific vegetative growths B flower C dissected perianth (dorsal sepal, two lateral petals, two lateral sepals fused into a synsepal and a view of the lip) D two views of the lip E column and lip, lateral view F column and ovary, ventral, dorsal and lateral views G anther cap and two views of the pollinarium. LCDP prepared by FP based on Karremans 5403.
Pleurothalliswinkeliana Karremans, Bogarín & Pupulin showing some floral variation A–EKarremans 5403FBogarín 9138GBogarín 9140. Photographs by the authors.
Diagnosis.
Morphologically closely resembling P.longipetala Bogarín & Belfort, but distinguished by the occasionally prolific plant that produces clumps of ramicauls (vs. non-prolific), the proportionally broader (ratio length:width = 2:1 vs 3:1) leaves, cordate in shape (vs. narrowly ovate-lanceolate) with overlapping basal lobes (vs. basal lobes non-overlapping), the smaller flower (<1.5 vs > 2 .0 cm), the shorter and narrower petals (5.4–5.7 × 0.7–0.8 vs. 7.9–8.6 × 1.6–1.7 mm) and by the lip, which is pendent and perpendicular to the column (vs erect, parallel to the column), and obscurely glandular (vs. thickly verrucose).
Description.
Epiphytic, caespitose, occasionally prolific, erect to suberect herb, up to 25 cm tall. Roots flexuous, thin, ca. 1 mm in diameter, densely spaced, appearing fasciculate. Primary ramicauls erect to suberect, slender, up to 9.0–23.0 cm long, with 2 tubular, tightly adpressed, papyraceous sheaths, up to 2.5–4.3 cm long, one close to the base, and another reaching the middle of the ramicaul. Secondary ramicauls 1.0–3.5 cm long, produced profusely from the floral meristem on the apex of primary ramicaul, being able to produce clumps of multiple ramicauls before severing naturally. Leaves horizontal to suberect, coriaceous, sessile, cordate, acute, with the basal lobes confluent to overlapping. Blades of primary ramicauls 4.0–9.6 × 1.5–4.0 cm, blades on secondary ramicauls 1.5–5.4 × 0.6–1.7 cm. Inflorescences persistent, forming successive multi-flowered coflorescences with a single open flower, subtended by a nearly prostrate or suberect spathe which appears deeply torn over time, ca. 1 cm long; pedicels terete, pale gray-yellow, with irregular black flecks. Ovary slightly clavate, bent, 3.0–3.2 mm long, green, with occasional black dots. Flowers spreading, transparent purple, with dark purple petals and lip, a whitish column, suffused with pink, and a yellowish anther cap. Dorsal sepal elliptic, acute, 3-veined, 8.5 × 4.0–4.2 mm. Lateral sepals connate in a narrowly ovate synsepal, acute, 4-veined, 7.3–7.8 × 4.7–4.8 mm. Petals linear, oblique, acute, 1-veined, 5.4–5.7 × 0.7–0.8 mm, with glandular margins. Lip triangular-ovate, resting on the synsepal, 2.6–2.8 × 2.3 mm, blade glandular, margins glandular-erose, acute, shortly apiculate, glenion a deep, oblong basal cavity. Column straight, transversely subrectangular, ca. 1.5 mm long, with a very short, glandular column foot, the anther and stigma apical. Anther cap ovate, cucullate, obtuse, 2-celled. Pollinarium composed of 2, narrowly ovate-pyriform pollinia, connected to a liquid, drop-like viscidium. Fruits and seeds unknown. This description is based on A.P. Karremans & J. Geml 5403.
Additional specimens examined.
Costa Rica. • Limón: Talamanca, Telire. Cordillera de Talamanca, Parque Internacional La Amistad (ACLA-C), sendero de la transtalamanca, bajando de la Fila Bugú hacia el Río Tapari, 09°26'48.00"N, 83°11'12.00"W, 1380 m, bosque pluvial premontano, bosque primario, epífita, a media luz, collected 27 April 2017, I. Chinchilla 3408, A.P. Karremans, G. Rojas-Alvarado, M. Cedeño, E. Kaes & O. Zúñiga [JBL-spirit, J0784! (prepared 14 May 2018), A0366! (prepared 2 May 2019)]. • Puntarenas: Buenos Aires, Potrero Grande, Altamira, Parque Internacional La Amistad, Sector Altamira, sendero al Valle del Silencio, cerca Cerro Hoffman, 9°04'56.2"N, 82°58'36.4"W, 2347 m, bosque pluvial montano, 30 August 2011, D. Bogarín 9138 & A.P. Karremans (Fig. 9F). Same locality and date, D. Bogarín 9140 & A.P. Karremans (Fig. 9F). • Puntarenas: Coto Brus, Sabalito, Zona Protectora Las Tablas, 13 km al noreste de Lucha, Sitio Coto Brus, entre Río Surá y Quebrada Sutú, Finca de Miguel Sandí, 8°56'46.1"N, 82°44'30.9"W, 1778 m, bosque pluvial montano bajo, epífita en potreros arbolados, collected 20 April 2012, A.P. Karremans & J. Geml 5403 (JBL-D6595! (prepared 9 January 2015), JBL-A0237! (prepared 29 November 2018), JBL-A0250! (prepared 21 February 2019), JBL-J1024! (prepared 12 July 2016).
Eponymy.
Honoring Dutch botanist Gab van Winkel (1955–2023), late editor of Orchideeën, journal of the Dutch Orchid Society (Nederlandse Orchideeën Vereniging), and director of the official website of the European Orchid Council (EOC). Gab has been recognized for devoting his life to the study of orchids (Anghelescu et al. 2024), and his untimely passing has been a great loss.
Phenology.
Flowering of P.winkeliana has been recorded from November to July, mostly corresponding to the dry season in Costa Rica.
Distribution and ecology.
Currently known only from the southern Cordillera de Talamanca in Costa Rica, where plants grow epiphytically in primary forests at around 1400–2300 m of elevation. Pleurothalliswinkeliana appears to be rare but is found on both watersheds of the Cordillera de Talamanca in southern Costa Rica, close to the Panamanian border, where it has been recorded in the neighboring Limón and Puntarenas provinces.
Notes.
Pleurothalliswinkeliana belongs to the P.phyllocardia assemblage (sensu Pupulin et al. 2021), characterized by presenting mostly narrow leaves, an erect to suberect, non prostrate spathaceous bract [except in P.adventurae Karremans & Bogarín (Karremans and Bogarín 2011: 112)], and coriaceous flowers that remain open after anthesis. In Costa Rica, this assemblage includes 13 species and a natural hybrid. Pleurothalliswinkeliana is most similar to P.longipetala, which was described from Tapantí in central Costa Rica, growing at an elevation of 1453 m. From the latter, P.winkeliana is mostly distinguished by the taller habit, and by its frequently prolific stems, that are topped by comparatively broad (ratio length:width = 2:1 vs 3:1) cordate leaves (vs. narrowly ovate-lanceolate), which bear overlapping (vs. non-overlapping) basal lobes. The flowers of Pleurothalliswinkeliana are smaller, with the smaller petals (5.4–5.7 × 0.7–0.8 vs. 7.9–8.6 × 1.6–1.7 mm), distinctly narrower at the base (vs. broadened at the base), and the lip is geniculate at the base and held perpendicularly to the column (vs. straight, parallel), with the blade obscurely glandular (vs. thick verrucose). The new species is also reminiscent of Pleurothallisanthurioides A.Doucette from Costa Rica and Panama (Pupulin et al. 2021), but the flowers are much smaller (e.g. sepal length 7.3–8.5 vs. 12–19 mm, petals 5.4–5.7 × 0.7–0.8 vs. 10–13 × 3–4 mm), the petals are linear (vs. oblong) and the lip triangular-ovate (vs. oblong-peltate).
Discussion
Despite occurring across the Pleurothallidinae (Figs 1, 2), prolification is a poorly studied phenomenon. The three new taxa described here differ slightly in their means of becoming prolific. Pleurothallismatrisilvae is typically prolific when growing as an epiphyte, while being mostly non-prolific when found growing on top of organic matter on the ground. Pleurothallispridgeoniana shows constitutive prolific growth, forming large, bushy plants, with each ramicaul eventually producing vegetative growths apically, resulting in chains of multiple ramicauls. Pleurothalliswinkeliana often forms clumps, rather than chains, of ramicauls at the apex of some, but not all, primary ramicauls. Be it constitutive or facultative, in the case of the genus Pleurothallis, prolification occurs more frequently in individuals and species growing at higher elevations.
Recent studies indicate that the highest rates of orchid speciation in the Neotropics occur in the humid and seasonal forests of Costa Rica and Panama (Pérez-Escobar et al. 2024). Specifically, Pleurothallidinae, which is the most species-rich group in this region, has experienced a rapid and recent radiation, reflected in a high diversity in Costa Rica (Pérez-Escobar et al. 2017). However, biogeographic studies on Pleurothallidinae are still limited due to incomplete species sampling and the reliance on a few molecular markers that exhibit low variability in previous studies (Pérez-Escobar et al. 2017). The sampling of multiple nuclear and plastid markers remains low within the subtribe, and it has not yet been tested whether including hundreds of markers could help resolve recalcitrant nodes in the phylogenetic analyses, specifically within Pleurothallis (Bogarín et al. 2018; Chumová et al. 2021), and thus clarify the exact phylogenetic placement of taxa in species-rich and non-monophyletic groups such as Pleurothallissubsect.Macrophyllae-Fasciculatae, which includes the three novelties.
The description of the three new Pleurothallis brings the total for the genus in Costa Rica to 67 species and four natural hybrids. In Costa Rica, Pleurothallis includes several species complexes that represent local radiations for which multiple species still require recognition, despite recent efforts. Novelties are often found on the highly diverse and largely unexplored Talamanca range in central and southern Costa Rica (Fig. 10).
Distribution map of P.matrisilvae, P.pridgeoniana, and P.winkeliana based on field collections.
To date, detailed analyses of species distributions have not been conducted, partly due to unresolved taxonomic issues and the lack of accuracy in species identification within many diverse or understudied genera of Pleurothallidinae, such as Lepanthes, Pleurothallis, and Stelis. Nevertheless, data available on Pleurothallis suggest distinct distribution patterns among Costa Rica’s mountain ranges. For instance, P.chavezii Luer is found only in the Cordillera de Guanacaste, while P.adventurae Karremans & Bogarín, P.anthurioides, and P.maduroi Luer are exclusive to the Cordillera de Talamanca. In contrast, other species, such as Pleurothallistonduzii Schltr., have a broader distribution across all mountain ranges. Meanwhile, P.angusta Ames & C.Schweinf., and P.phyllocardia Rchb.f. are widespread in the Cordillera de Talamanca and Cordillera Central but do not extend to the Cordillera de Tilarán or Guanacaste (Pupulin et al. 2021). No collections of the three novelties described herein have been found in other mountain ranges of Costa Rica, suggesting that their distribution may be restricted to the Cordillera de Talamanca. Based on previous observations of other Pleurothallidinae species, we do not expect to find these newly described species in the northern mountain ranges of Guanacaste and Tilarán.
A comprehensive study comparing the diversity of orchids across Costa Rica’s mountain ranges has not yet been conducted. By completing the ongoing taxonomic work on these complex and diverse groups within Pleurothallis, integrating new distribution data from recently described species, we can establish a stronger foundation for more accurate biogeographical analyses.
Supplementary Material
XML Treatment for Pleurothallis matrisilvae
XML Treatment for Pleurothallis pridgeoniana
XML Treatment for Pleurothallis winkeliana
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aiton WT (1813) Hortus Kewensis; or, a Catalogue of the Plants Cultivated in the Royal Botanic Garden at Kew. Vol. 5. London, 568 pp.
- 2Ames O (1922) Pleurothallisrowleei.Schedulae Orchidianae 1: 11–12. 10.5962/p.388177 · doi ↗
- 3Anghelescu NE Balogh L Balogh M Kigyossy N Georgescu MI Petra SA Toma F Peticila AG (2024) Gymnadeniawinkeliana—a new orchid species to Romanian flora. Plants 13: 1363. 10.3390/plants 13101363 PMC 1112507638794434 · doi ↗ · pubmed ↗
- 4Belfort-Oconitrillo N Salguero G Oses L Gil-Amaya K Rojas-Alvarado G Chinchilla IFDíaz-Morales M Pupulin F Bogarín D Karremans AP (2024) New species and records of Orchidaceae from Costa Rica. IV.Lankesteriana 24(2): 141–192. 10.15517/lank.v 24i 2.60686 · doi ↗
- 5Bell AD Bryan A (2008) Plant form: An illustrated guide to flowering plant morphology.Timber Press, Portland, 431 pp.
- 6Bogarín DPérez-Escobar OA Groenenberg D Holland SD Karremans AP Lemmon EM Lemmon AR Pupulin F Smets E Gravendeel B (2018) Anchored hybrid enrichment generated nuclear, plastid and mitochondrial markers resolve the Lepantheshorrida (Orchidaceae: Pleurothallidinae) species complex.Molecular Phylogenetics and Evolution 129: 27–47. 10.1016/j.ympev.2018.07.01430041026 · doi ↗ · pubmed ↗
- 7ChumováZZáveskáE HlouškováP Ponert J Schmidt P-AČertner M MandákováT Trávníček P (2021) Repeat proliferation and partial endoreplication jointly shape the patterns of genome size evolution in orchids.The Plant Journal 107: 511–524. 10.1111/tpj.1530633960537 · doi ↗ · pubmed ↗
- 8Garay LA (1956) Studies in American orchids. I.Canadian Journal of Botany 34(2): 241–260. 10.1139/b 56-022 · doi ↗