Morphological and phylogenetic analysis reveal three new species Phyllosticta (Phyllostictaceae, Botryosphaeriales) in China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
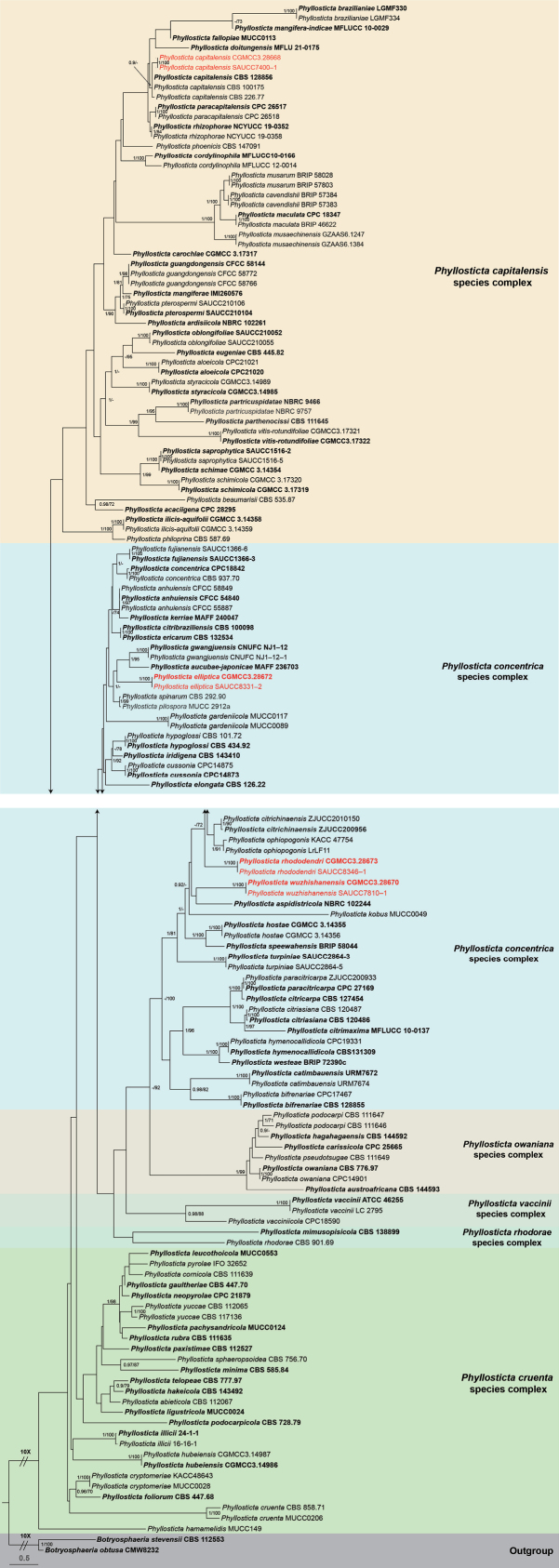 Figure 1
Figure 1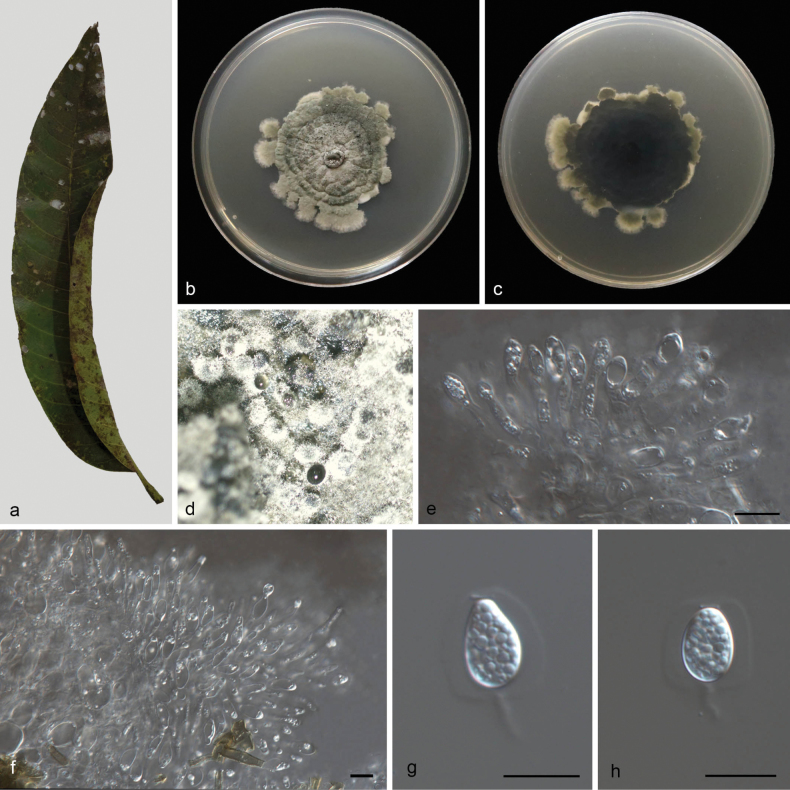 Figure 2
Figure 2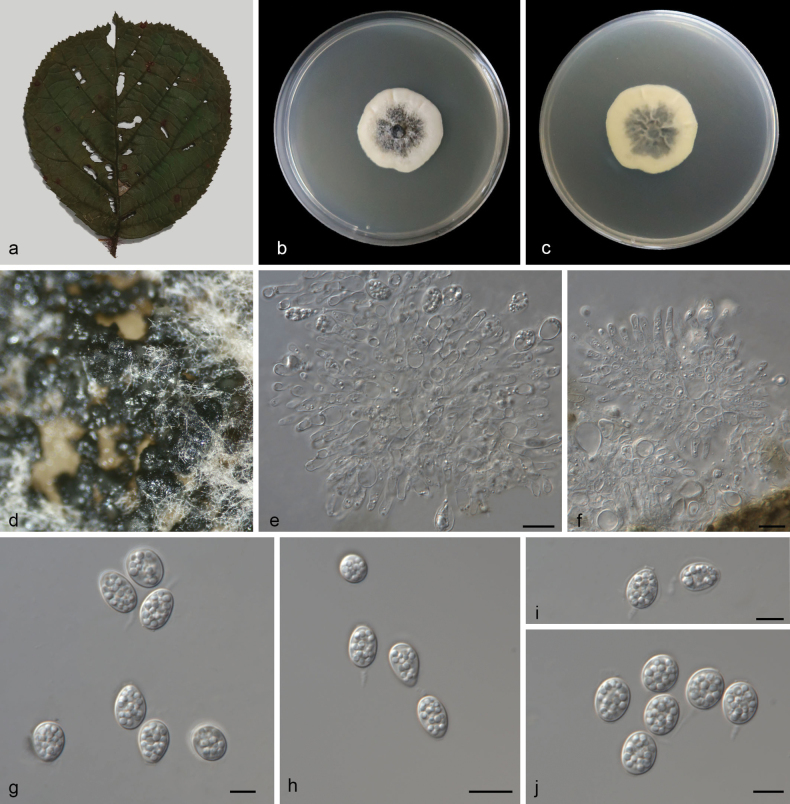 Figure 3
Figure 3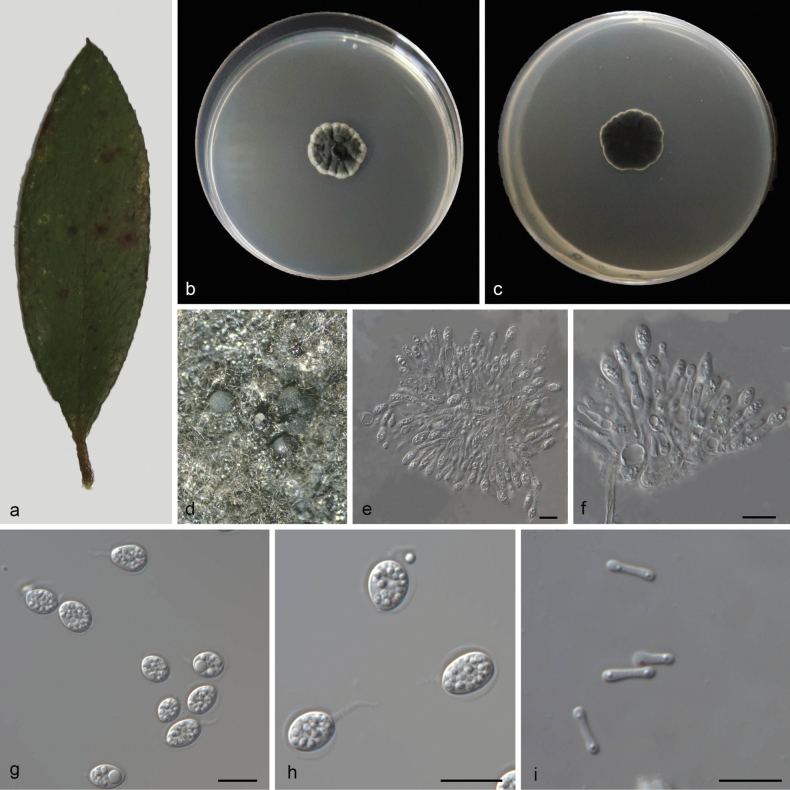 Figure 4
Figure 4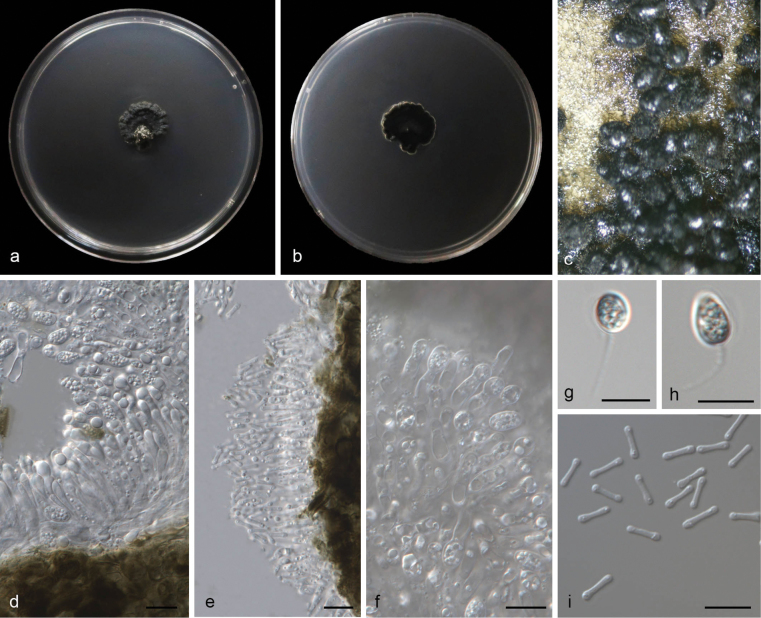 Figure 5
Figure 5- —National Natural Science Foundation of China 501100001809 https://ror.org/01h0zpd94 http://doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions · Lichen and fungal ecology
Introduction
Phyllosticta Pers. was established by Persoon in 1818. Phyllostictaconvallariae was designated as the type species (Donk 1968), which was later synonymized with P.cruenta (van der Aa 1973, Wikee et al. 2013a). Phyllosticta is a significant plant pathogenic genus known for causing leaf spots and fruit diseases on a wide range of economically important plants and ornamentals. Notable species include P.citricarpa (McAlpine) Aa, which causes citrus black spot (Baayen et al. 2002; Glienke et al. 2011); P.ampelicida (Engelm.) Aa, which is responsible for black rot disease on Vitisvinifera in North America (Kuo and Hoch 2018); and Phyllostictaophiopogonis Wulandari & K.D. Hyde, which causes leaf spots of Ophiopogonjaponicus in Thailand (Wikee et al. 2012). Phyllostictacapitalensis acts as a common endophyte with antagonistic potential against pathogenic species such as P.citricarpa and produces melanized appressoria for host penetration (Wikee et al. 2013b; Tran et al. 2019). A total of 3,226 Phyllosticta names are recorded in the Index Fungorum (accessed on 21 March 2025).
Since Phyllosticta is distinct from other genera in its family, Seaver (1922) initially classified it in the family Phyllostictaceae Fr. and placed it in the order Phyllostictales. However, subsequent studies (Crous et al. 2006; Schoch et al. 2006; Liu et al. 2012) reassigned Phyllosticta to the family Botryosphaeriaceae Theiss. & Syd., within the order Botryosphaeriales C.L. Schoch et al. However, phylogenetic analysis by Wikee et al. (2013a) positioned Phyllosticta in a clade sister to Botryosphaeriaceae, leading to its reclassification into the family Phyllostictaceae under the order Botryosphaeriales (Zhang et al. 2013). In recent years, an increasing number of new Phyllosticta species have been described based on a combination of molecular data and morphological characteristics (Su and Cai 2012; Wang et al. 2012, 2013; Wong et al. 2012; Zhang et al. 2012, 2013; Wikee et al. 2013a; Wulandari et al. 2013; Crous et al. 2014, 2016, 2017, 2018, 2019, 2021; Zhou et al. 2015; Guarnaccia et al. 2017; Lin et al. 2017; Hattori et al. 2020; Norphanphoun et al. 2020). Norphanphoun et al. (2020) compiled all species classified under Phyllosticta in GenBank, analyzing a comprehensive dataset of five loci, and consequently proposed six species complexes: the P.capitalensis species complex, P.concentrica species complex, P.cruenta species complex, P.owaniana species complex, P.rhodorae species complex, and P.vaccinii species complex. Subsequently, multiple novel taxa or new records were introduced based on morphological descriptions and multi-locus phylogenetic analysis (Zhang et al. 2023; Gomdola et al. 2024; Jiang et al. 2024).
The sexual morph of Phyllosticta species is characterized by erumpent, uniloculate, globose to subglobose ascomata with a central ostiole and pseudoparaphyses at maturity. Asci are clavate to ellipsoidal or ovoid, with an ocular chamber, while ascospores are aseptate, hyaline, ellipsoidal to limoniform, guttulate, and smooth-walled, often with mucoid caps at both ends (van der Aa 1973, Wong et al. 2012, Wikee et al. 2013a). The asexual morph produces conidia that are generally aseptate, hyaline, ovoid to ellipsoidal, and are surrounded by a mucilaginous sheath and possess an apical appendage. The morphology of appendages may vary depending on the growth medium (Wikee et al. 2013a). Spermatia are hyaline, aseptate, cylindrical to dumbbell-shaped, with guttules at each end. Because of overlapping morphological traits, species delimitation relies primarily on multi-locus phylogenetic analysis (Norphanphoun et al. 2020). Fungi associated with leaf spots were collected from Mangiferaindica, Rhododendron×pulchrum, and Rubusellipticusvar.obcordatus. We used sequences of five gene loci, including the internal transcribed spacer of ribosomal RNA (ITS rDNA), large subunit of ribosomal RNA (LSU rDNA), translation elongation factor 1 alpha (tef1), actin (ACT), and glycerol-3-phosphate dehydrogenase (GPDH). We also incorporated their morphology and then identified these fungi as three new species of the P.concentrica species complex.
Materials and methods
Isolation and morphological studies
Dead, healthy, and diseased leaves were collected from Hainan and Yunnan (from 2023 to 2024) and transported to the laboratory in paper bags. Fungal isolates were obtained using the tissue isolation method (Zhang et al. 2023, 2025). Leaf lesion fragments (5 × 5 mm) were excised from the margins of symptomatic tissues, surface-sterilized by sequential immersion in 75% ethanol for 30 seconds, rinsed in sterile distilled water, then treated with 5% sodium hypochlorite solution for 30 seconds, and finally rinsed in sterile distilled water for 1 minute (Jiang et al. 2021). After drying on sterilized tissue paper, the fragments were placed on 2% PDA and incubated at 25 °C for two to four days. Actively growing hyphal tips were subsequently transferred to fresh PDA plates and cultured for morphological examination. Colony morphology was documented on days 7 and 14 using a digital camera (Canon Powershot G7X, Canon Co., Ltd., Beijing, China). Micromorphological features were observed using an Olympus SZX10 stereomicroscope and Olympus BX53 microscope (Olympus Corporation, Tokyo, Japan), both equipped with an Olympus DP80 high-resolution color digital camera (Olympus Corporation, Tokyo, Japan) for imaging fungal structures. All fungal strains were stored in 10% sterilized glycerin at 4 °C for further studies. Structural measurements were performed using the Digimizer software (https://www.digimizer.com/), with thirty measurements taken for each morphological feature. Holotype specimens were deposited in the Herbarium of Plant Pathology, Shandong Agricultural University (HSAUP). Ex-type cultures and other living cultures were deposited in the China General Microbiological Culture Collection Center (CGMCC), Beijing, China, and the Shandong Agricultural University Culture Collection (SAUCC), Shandong, China. Taxonomic information for the newly described taxa was registered to MycoBank (http://www.mycobank.org).
DNA extraction and sequencing
Fungal DNA was extracted from fungal mycelia grown on PDA using a modified cetyltrimethylammonium bromide (CTAB) protocol or a kit method (OGPLF-400, GeneOnBio Corporation, Changchun, China) for Sanger sequencing (Wang et al. 2023; Zhang et al. 2023, 2025). In this study, a total of eight pairs of primers were used; the internal transcribed spacer region (ITS) with intervening 5.8S rRNA gene, large subunit of rRNA gene (LSU), translation elongation factor 1-alpha gene (tef1), actin gene (ACT), and glyceraldehyde-3-phosphate dehydrogenase gene (GPDH) were amplified and sequenced using the primer pairs ITS5/ITS4 (White et al. 1990), LR0R/LR5 (White et al. 1990), EF1-728F/EF2 (O’Donnell et al. 1998; Carbone and Kohn 1999), ACT-512F/ACT-783R (Carbone and Kohn 1999), and GDF1/GAPDH (Myllys et al. 2002), respectively. Amplification reactions were performed in a 20 μL reaction volume, which contained 10 μL 2 × Hieff Canace® Plus PCR Master Mix (Shanghai, China) (with dye) (Yeasen Biotechnology, Cat. No. 10154ES03), 0.5 μL of each forward and reverse primer (10 μM) (TsingKe, Qingdao, China), and 1 μL template genomic DNA, adjusted with distilled deionized water to a total volume of 20 μL. PCR amplification products were visualized on 2% agarose electrophoresis gel. The Gel Extraction Kit (Cat: AE0101-C) (Shandong Sparkjade Biotechnology Co., Ltd.) was used for gel recovery. Sanger sequencing was performed using an Eppendorf Master Thermocycler (Hamburg, Germany) at the Youkang Biotechnology Co., Ltd. (Qingdao, China) bi-directionally. All sequences generated in this study were deposited in GenBank (Suppl. material 1).
Phylogenetic analysis
The generated consensus sequences were subjected to BLAST searches in NCBI’s GenBank nucleotide database to identify closely related sequences (Zhang et al. 2000). For phylogenetic analysis based on ITS, LSU, tef1, ACT, and GPDH sequences, a subset of alignments from Jiang et al. (2024) was used as the backbone. Newly generated sequences from this study were aligned with related sequences retrieved from GenBank (Suppl. material 1) using the online tool MAFFT 7 with the Auto strategy (Katoh et al. 2019; http://mafft.cbrc.jp/alignment/server/) and corrected manually using BioEdit (Hall 2006). To determine the species identity of the isolates, phylogenetic analysis was initially performed for each locus separately, followed by a concatenated analysis of all loci (ITS-LSU-tef1-ACT-GPDH).
For phylogenetic analysis, we followed the methods by Zhang et al. (2023) using both maximum likelihood (ML) and Bayesian inference (BI) algorithms. ML was run on the CIPRES Science Gateway portal (Miller et al. 2012) (https://www.phylo.org/) or with RAxML-NG v1.2.1 (https://github.com/amkozlov/raxml-ng), and Bayesian inference (BI) analysis was performed using MrBayes v3.2.7a, running with 8 threads on a Linux system. For ML analysis, the default parameters were used, and 100 rapid bootstrap replicates were run with the GTR+G+I model of nucleotide evolution; BI analysis was performed using a fast bootstrap algorithm with an automatic stop option (Zhang et al. 2023). The burn-in fraction was set to 0.25, and posterior probabilities (PP) were determined from the remaining trees. All resulting trees were plotted using FigTree v. 1.4.4 (http://tree.bio.ed.ac.uk/software/figtree), and the layout of the trees was edited in Adobe Illustrator CC 2019.
Results
Phylogenetic analysis
A total of 149 isolates representing Phyllosticta species were subjected to phylogenetic analysis, with Botryosphaeriaobtusa (CGMCC 3.14986) and B.stevensii (CBS 112553) designated as outgroup taxa. The final alignment consisted of 2,934 characters, corresponding to the following loci: 1–669 (ITS), 670–1,409 (LSU), 1,410–1,917 (tef1), 1,918–2,186 (ACT), and 2,187–2,934 (GPDH). Among these, 1,775 characters were constant, 215 were variable and parsimony-uninformative, and 944 were parsimony-informative. ML analysis yielded the best-scoring RAxML tree with a final likelihood value of –26,187.533107. The alignment contained 1,432 distinct patterns, with 36.76% undetermined characters or gaps. The estimated base frequencies were as follows: A = 0.200968, C = 0.314322, G = 0.270974, and T = 0.213736; substitution rates were AC = 1.162897, AG = 3.568637, AT = 1.418189, CG = 1.163532, CT = 6.776257, and GT = 1.0. The gamma distribution shape parameter alpha was estimated at 0.679976. As the ML and BI trees produced topologically congruent trees, only the ML tree (Fig. 1) is presented, with posterior probabilities and bootstrap provided for well-supported clades. Based on the five-gene phylogenetic framework (Fig. 1), the 149 isolates were assigned to 98 species. The present study identified three novel species, viz. Phyllostictaelliptica sp. nov., P.rhododendri sp. nov., and P.wuzhishanensis sp. nov.
Phylogram of the Phyllosticta, inferred from a concatenated alignment of ITS, LSU, tef1, ACT, and GPDH sequences. Botryosphaeriaobtusa (CMW8232) and B.stevensii (CBS 112553) were used as outgroup taxa. BI posterior probabilities and ML bootstrap support values above 0.90 and 70% are shown at the first and second positions, respectively. Ex-type cultures are highlighted in bold, while strains obtained in this study are marked in red. Some branches have been shortened for layout optimization, indicated by double diagonal lines with the corresponding reduction factor. The scale bar at the bottom left represents the number of substitutions per site.
Taxonomy
Phyllostictacapitalensis species complex
Based on molecular analysis and morphological characteristics, the Phyllostictacapitalensis species complex comprises 32 species: P.acaciigena, P.aloeicola, P.ardisiicola, P.beaumarisii, P.brazilianiae, P.capitalensis, P.carochlae, P.cavendishii, P.cordylinophila, P.doitungensis, P.eugeniae, P.fallopiae, P.guangdongensis, P.ilicis-aquifolii, P.maculata, P.mangiferae, P.mangifera-indicae, P.musaechinensis, P.musarum, P.oblongifoliae, P.paracapitalensis, P.parthenocissi, P.partricuspidatae, P.philoprina, P.phoenicis, P.pterospermi, P.rhizophorae, P.saprophytica, P.schimae, P.schimicola, P.styracicola, and P.vitis-rotundifoliae.
Phyllosticta
capitalensis
Taxon classificationFungiBotryosphaerialesPhyllostictaceae
Henn., Hedwigia 48: 13 (1908)
51CE15EB-41BF-5200-B9E9-83172F695A7A
Description.
Leaf endogenic and associated with leaves of Mangiferaindica. Sexual morph: Not observed. Asexual morph: Conidiomata pycnidial, mostly aggregated in clusters, black, erumpent. In PDA culture, exuding colorless to opaque conidial masses within 12 d or longer. Pycnidial walls multilayered, textura angularis, brown, inner walls of hyaline. Conidiophores indistinct, often reduced to conidiogenous cells. Conidiogenous cells 6–12 × 2.2–2.8 μm, subcylindrical, ampulliform, hyaline, smooth. Conidia 6.7–12 × 3.6–5.8 μm (L/W 1.42–1.53), ovoid, ampulliform, ellipsoidal to subglobose, hyaline, aseptate, thin and smooth walled, multi-guttulate, or with a single large central guttule, surrounded by a mucilaginous sheath. Sheath 2.6–2.9 μm thick, thicker on both sides, and bearing a hyaline, apical mucoid appendage. Appendages 5.7–6.7 × 1.2–2.1 μm, flexible, unbranched, tapering towards an acutely rounded tip.
Phyllostictacapitalensis (CGMCC3.28668) a diseased leaf of Mangiferaindicab, c colonies (left-above, right-reverse) after 14 d on PDA d conidiomata e, f conidiogenous cells with conidia g, h conidia. Scale bars: 10 μm (e–h).
Culture characteristics.
Colonies on PDA 46–52 mm in diameter after 14 d at 25 °C in darkness, with a growth rate of 3.3–3.7 mm/day, undulate at edge, grey white to greenish-black on obverse and reverse.
Additional specimen examined.
China • Hainan Province, Wanning City, Xinglong tropical botanical garden, on diseased leaves of Mangiferaindica L., 28 March 2024, M.Y. Zhang (HSAUP7403), living culture CGMCC3.28668; ibid., Z.X. Zhang (HSAUP7400), living culture SAUCC7400–1.
Notes.
The holotype (CBS 128856) of P.capitalensis was collected from Stanhopeagraveolens (Glienke et al. 2011). Two isolates (CGMCC3.28668 and SAUCC7400–1), collected from diseased leaves of Mangiferaindica, cluster in the P.capitalensis clade (Fig. 1). In morphology, they possess the same morphological characters, such as subcylindrical to ampullate conidiogenous cells (6–12 × 2.2–2.8 vs. 7–10 × 3–5 μm), ellipsoidal to subglobose conidia (6.7–12 × 3.6–5.8 vs. 11–12 × 6–7 μm), and hyaline, apical mucoid appendages (5.7–6.7 × 1.2–2.1 vs. 6–8 × 1–1.5 μm). Therefore, we defined these two isolates as P.capitalensis.
Phyllostictaconcentrica species complex
Based on molecular analysis and morphological characteristics, the Phyllostictaconcentrica species complex comprises 32 species: P.anhuiensis, P.aspidistricola, P.aucubae-japonicae, P.bifrenariae, P.catimbauensis, P.citriasiana, P.citribraziliensis, P.citricarpa, P.citrichinensis, P.citri-maxima, P.concentrica, P.cussonia, P.elliptica, P.elongata, P.ericarum, P.fujianensis, P.gardeniicola, P.gwangjuensis, P.hostae, P.hymenocallidicola, P.hypoglossi, P.iridigena, P.kerriae, P.kobus, P.ophiopogonis, P.paracitricarpa, P.pilospora, P.rhododendri, P.speewahensis, P.turpiniae, P.westeae, and P.wuzhishanensis.
Phyllosticta
elliptica
Taxon classificationFungiBotryosphaerialesPhyllostictaceae
M.Y. Zhang, Z.X. Zhang & X.G. Zhang sp. nov.
B28C49F7-70A1-559A-B228-9EEE7D319D08
857335
Etymology.
The specific epithet “elliptica” refers to the genus name of the host plant Rubusellipticusvar.obcordatus.
Type.
China • Yunnan Province, Hongta District, Yuxi City, Longma Mountain Scenic Area, on diseased leaves of Rubusellipticusvar.obcordatus (Franch.) Focke, 12 May 2024, M.Y. Zhang (holotype HSAUP8332), ex-type living culture CGMCC3.28672.
Description.
Leaf endogenic and associated with leaves of Rubusellipticusvar.obcordatus. Sexual morph: Not observed. Asexual morph: Conidiomata pycnidial, mostly aggregated in clusters, black, erumpent. In PDA culture, exuding white conidial masses within 15 days or longer. Pycnidial walls multilayered, textura angularis, brown, inner walls of hyaline. Conidiophores indistinct, often reduced to conidiogenous cells. Conidiogenous cells 7–15 × 1.8–4 μm, cylindrical, hyaline, smooth. Conidia 12–15 × 7.5–11 μm (L/W 1.17–1.54), ovoid, ampulliform, ellipsoidal to subglobose, hyaline, aseptate, thin and smooth walled, multi-guttulate, or with a single large central guttule, surrounded by a mucilaginous sheath. Sheath 1.3–1.8 μm thick, thicker on both sides, and bearing a hyaline, apical mucoid appendage. Appendages 4–12 × 1–1.2 μm, flexible, unbranched, tapering towards an acutely rounded tip.
Phyllostictaelliptica (CGMCC3.28672) a diseased leaf of Rubusellipticusvar.obcordatusb, c colonies (left-above, right-reverse) after 14 d on PDA d conidiomata e, f conidiogenous cells with conidia g–j conidia. Scale bars: 10 μm (e–j).
Culture characteristics.
Colonies on PDA 33–36 mm in diameter after 14 d at 25 °C in darkness, with a growth rate of 2.3–2.6 mm/day, undulate at edge, creamy white to black in obverse and reverse.
Additional specimen examined.
China • Yunnan Province, Hongta District, Yuxi City, Longma Mountain Scenic Area, on dead leaves, 12 May 2024, M.Y. Zhang (HSAUP8331), living culture SAUCC8331–2.
Notes.
Phyllostictaelliptica is closely related to P.aucubae-japonicae (MAFF 236703) and P.gwangjuensis (CNUFC NJ1–12 and CNUFC NJ1-12-1) based on DNA sequence data in BLAST searches and phylogenetic analysis (Fig. 1). However, P.elliptica differs from P.aucubae-japonicae by 70 nucleotides (31/628 in ITS, 0/737 in LSU, 22/266 in tef, 17/239 in ACT, and 0/725 in GPDH) and from P.gwangjuensis by 60 nucleotides (25/634 in ITS, 0/737 in LSU, 16/376 in tef1, 19/214 in ACT, and 0/725 in GPDH). In morphology, they are distinguished by different hosts (Rubusellipticusvar.obcordatus vs. Aucubajaponica vs. Torreyanucifera) and longer conidia in Phyllostictaelliptica than in P.aucubae-japonicae and P.gwangjuensis (12–15 × 7.5–11 μm (L/W 1.17–1.54) vs. 10–13 × 5–8.5 μm (L/W 1.41–1.65) vs. (8.5–)10–13.5 × 7–9(–9.5) μm (L/W 1.40–1.53)) (Hernandez-Restrepo et al. 2016; Nguyen et al. 2022). Therefore, based on morphology and phylogenetic evidence, we establish this fungus as Phyllostictaelliptica sp. nov.
Phyllosticta
rhododendri
Taxon classificationFungiBotryosphaerialesPhyllostictaceae
M.Y. Zhang, Z.X. Zhang & X.G. Zhang sp. nov.
A7908DF2-AEAB-5FA7-821D-8A0588E45590
857221
Etymology.
The specific epithet “rhododendri” refers to the host plant Rhododendron×pulchrum Sweet.
Type.
China • Yunnan Province, Hongta District, Yuxi City, Longma Mountain Scenic Area, on diseased leaves of Rhododendron×pulchrum, 13 May 2024, M.Y. Zhang (holotype HSAUP8342), ex-type living culture CGMCC3.28673.
Description.
Leaf endogenic and associated with leaves of Rhododendron×pulchrum. Sexual morph: Not observed. Asexual morph: Conidiomata pycnidial, mostly aggregated in clusters, black, erumpent. In PDA culture, exuding white conidial masses within 10 days or longer. Pycnidial wall multi-layered, textura angularis, brown to dark brown. Conidiophores indistinct, often reduced to conidiogenous cells. Conidiogenous cells 7–16 × 1.4–5.2 μm, subcylindrical, ampulliform, hyaline, smooth. Conidia 8.5–11.5 × 6.8–9.4 µm (L/W 1.33–1.53), ovoid, ampulliform, ellipsoidal to subglobose, hyaline, aseptate, thin and smooth walled, multi-guttulate, or with a single large central guttule, surrounded by a mucilaginous sheath. Sheath 0.8–1.5 μm thick, thicker on both sides, and bearing a hyaline, apical mucoid appendage. Appendages 3–7.5 × 1–1.2 μm, flexible, unbranched, tapering towards an acutely rounded tip. Spermatia 6.3–8.2 × 1–1.5 μm, occurring in conidioma with conidia, hyaline, smooth, guttulate to granular, bacilliform.
Phyllostictarhododendri (CGMCC3.28673) a diseased leaf of Rhododendron×pulchrumb, c colonies (left-above, right-reverse) after 14 days on PDA d conidiomata e, f conidiogenous cells with conidia g, h conidia i spermatia. Scale bars: 10 μm (e–i).
Culture characteristics.
Colonies on PDA 19–22 mm in diameter after 14 days at 25 °C in darkness, with a growth rate of 1.4–1.6 mm/day, greenish black with white edge on obverse and reverse.
Additional specimens examined.
China • Yunnan Province, Hongta District, Yuxi City, Longma Mountain Scenic Area, on diseased leaves of Rhododendron×pulchrum, 13 May 2024, Z.X. Zhang (HSAUP8346), living culture SAUCC8346–1.
Notes.
Phylogenetic analysis showed that Phyllostictarhododendri formed an independent clade that is sister to a large clade comprising over ten species (P.anhuiensis, P.aucubae-japonicae, P.citribraziliensis, P.citrichinaensis, P.concentrica, P.cussonia, P.elongata, P.elliptica, P.ericarum, P.fujianensis, P.gardeniicola, P.gwangjuensis, P.hypoglossi, P.iridigena, P.kerriae, P.ophiopogonis, and P.spinarum) (Fig. 1). In morphology, Phyllostictarhododendri was found to produce spermatia, a structure that is rarely observed within this genus. Therefore, we establish this fungus as Phyllostictaelliptica sp. nov.
Phyllosticta
wuzhishanensis
Taxon classificationFungiBotryosphaerialesPhyllostictaceae
M.Y. Zhang, Z.X. Zhang & X.G. Zhang sp. nov.
371DA4D1-75A3-528F-9C51-EED858886A6C
857226
Etymology.
The epithet “wuzhishanensis” pertains to Wuzhishan National Nature Reserve, where the type was collected.
Type.
China • Hainan Province, Wuzhishan National Nature Reserve, on saprophytic (dead leaves) leaves, 28 March 2024, M.Y. Zhang (holotype HSAUP7814), ex-type living culture CGMCC3.28670.
Description.
Endophytic on saprophytic (dead leaves) leaves. Sexual morph: Not observed. Asexual morph: Conidiomata pycnidial, mostly aggregated in clusters, black, erumpent. In PDA culture, exuding colorless to opaque conidial masses within 12 days or longer. Pycnidial walls multilayered, textura angularis, brown, inner walls of hyaline. Conidiophores indistinct, often reduced to conidiogenous cells. Conidiogenous cells 6.5–14.5 × 3–4 μm, subcylindrical, ampulliform, hyaline, smooth. Conidia 5.5–10 × 3.8–8.6 μm (L/W 1.27–1.83), ovoid, ampulliform, ellipsoidal to subglobose, hyaline, aseptate, thin and smooth walled, multi-guttulate, or with a single large central guttule, surrounded by a mucilaginous sheath. Sheath 1.6–2 μm thick, thicker on both sides, and bearing a hyaline, apical mucoid appendage. Appendages 10–13 × 1.2–1.5 μm, flexible, unbranched, tapering towards an acutely rounded tip. Spermatia 6.3–8.2 × 1–1.5 μm, occurring in conidioma with conidia, hyaline, smooth, guttulate to granular, bacilliform.
Phyllostictawuzhishanensis (CGMCC3.28670) a, b colonies (left-above, right-reverse) after 14 d on PDA c conidiomata d–f conidiogenous cells with conidia g, h conidia i spermatia. Scale bars: 10 μm (d–i).
Culture characteristics.
Colonies on PDA 17–21 mm in diameter after 14 days at 25 °C in darkness, with a growth rate of 1.2–1.5 mm/day, undulate at edge, black on obverse and reverse.
Additional specimen examined.
China • Hainan Province: Wuzhishan National Nature Reserve, on saprophytic (dead leaves) leaves, 28 March 2024, M.Y. Zhang (HSAUP7810), living culture SAUCC7810–1.
Notes.
Phyllostictawuzhishanensis is closely related to P.aspidistricola (NBRC 102244) based on DNA sequence data in BLAST searches and phylogenetic analysis (Fig. 1). However, P.wuzhishanensis differs from P.aspidistricola by 69 nucleotides (40/621 in ITS, 0/737 in LSU, 0/411 in tef1, 29/252 in ACT, and 0/727 GPDH). In morphology, they are distinguished by different hosts (dead leaves vs. Aspidistraelatior) and conidial size (5.5–10 × 3.8–8.6 μm (L/W 1.27–1.83) in P.wuzhishanensis vs. 9.5–12.5 × 8.5–10 μm (L/W 1.43–1.63) in P.aspidistricola) (Motohashi et al. 2008). Based on morpho-molecular evidence, we establish this fungus as Phyllostictawuzhishanensis sp. nov.
Phyllostictacruenta species complex
Based on molecular analysis and morphological characteristics, the Phyllostictacruenta species complex comprises 22 species: P.abieticola, P.cornicola, P.cruenta, P.cryptomeriae, P.foliorum, P.gaultheriae, P.hakeicola, P.hamamelidis, P.hubeiensis, P.illicii, P.leucothoicola, P.ligustricola, P.minima, P.neopyrolae, P.pachysandricola, P.paxistimae, P.podocarpicola, P.pyrolae, P.rubra, P.sphaeropsoidea, P.telopeae, and P.yuccae.
Phyllostictaowaniana species complex
Based on molecular analysis and morphological characteristics, the Phyllostictaowaniana species complex comprises 6 species: P.austroafricana, P.carissicola, P.hagahagaensis, P.owaniana, P.podocarpi, and P.pseudotsugae.
Phyllostictarhodorae species complex
Based on molecular analysis and morphological characteristics, the Phyllostictarhodorae species complex comprises 2 species: P.mimusopisicola and P.rhodorae.
Phyllostictavaccinii species complex
Based on molecular analysis and morphological characteristics, the Phyllostictavaccinii species complex comprises 2 species: P.vaccinii and P.vacciniicola.
Discussion
In modern fungal taxonomy, integrating molecular data with morphological characteristics has become essential. As the limitations of traditional classification systems become increasingly evident, mycologists have turned to divergence time estimates and phylogenomic data to define clearer taxonomic boundaries (Baayen et al. 2002; Glienke et al. 2011; Wikee et al. 2013a, 2013b; Crous et al. 2014, 2016, 2017, 2018, 2019, 2021; Zhou et al. 2015; Guarnaccia et al. 2017; Lin et al. 2017; Hattori et al. 2020; Norphanphoun et al. 2020; Zhang et al. 2023, 2025). Historically, the identification of Phyllosticta species has relied on morphological characteristics and host associations. However, due to significant overlap in morphological traits among species, recognizing homologous characters has remained a challenge, leading to long-standing difficulties in Phyllosticta species delimitation (Norphanphoun et al. 2020). The advent of molecular phylogenetic approaches has greatly enhanced species recognition and the classification of species complexes (Baayen et al. 2002; Okane et al. 2003; Motohashi et al. 2009; Wulandari et al. 2009; Glienke et al. 2011; Wikee et al. 2012). The ITS region is widely used as a primary genetic marker for genus-level identification (White et al. 1990). However, for accurate species delimitation, additional loci such as LSU, tef1, ACT, and GPDH are required to achieve sufficient resolution (Norphanphoun et al. 2020). To date, six major species complexes encompassing 96 accepted species have been recognized within Phyllosticta, including P.capitalensis species complex (including 32 species), P.concentrica species complex (including 32 species), P.cruenta species complex (including 22 species), P.owaniana species complex (including six species), P.rhodorae species complex (including two species), and P.vaccinii species complex (including two species).
In this study, we described and illustrated four Phyllosticta isolates recovered from three host genera (Mangiferaindica, Rhododendron×pulchrum, and Rubusellipticusvar.obcordatus) based on a multi-locus phylogenetic analysis combined with morphological observations in culture. Three new species were proposed, including Phyllostictaelliptica sp. nov. (Rubusellipticusvar.obcordatus), P.rhododendri sp. nov. (Rhododendron×pulchrum), and P.wuzhishanensis sp. nov. (saprophytic leaves). These species were assigned to the P.concentrica species complex. Additionally, Phyllostictapilospora (MUCC 2912a) was confirmed as a member of the P.concentrica species complex (Hattori et al. 2020). Phylogenetic analysis (Fig. 1, Suppl. material 1) and morphological comparisons revealed that P.pilospora (MUCC 2912a) and P.spinarum (CBS 292.90) are closely related (Hattori et al. 2020; Gomzhina and Gannibal 2023).
The USDA Fungal Database contains over 7,500 records of Phyllosticta species associated with plant hosts (excluding synonyms) (https://fungi.ars.usda.gov/, accessed 21 March 2025) (Okane et al. 2001, 2003; Baayen et al. 2002; Glienke et al. 2011; Wikee et al. 2013b; Wu et al. 2014; Zhang et al. 2015; Tran et al. 2019; Hattori et al. 2020; Norphanphoun et al. 2020; Zhang et al. 2022; Gomdola et al. 2024; Jiang et al. 2024). Additionally, Phyllostictacapitalensis is recognized as a globally distributed endophyte and weak plant pathogen, with over 400 host-associated records listed in the USDA Fungal Database (Glienke-Blanco et al. 2002; Silva and Pereira 2007; Silva et al. 2008; Wikee et al. 2013a). These findings suggest that certain Phyllosticta taxa exhibit widespread endophytic lifestyles, warranting further investigation into their ecological roles and evolutionary adaptations.
Supplementary Material
XML Treatment for Phyllosticta capitalensis
XML Treatment for Phyllosticta elliptica
XML Treatment for Phyllosticta rhododendri
XML Treatment for Phyllosticta wuzhishanensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baayen RP Bonants P Verkley G Carroll G Cvan der Aa H Ade Weerdt Mvan Brouwershaven IR Schutte GC Maccheroni Jr W Glienke de Blanco C Azevedo JL (2002) Nonpathogenic isolates of the citrus black spot fungus, Guignardiacitricarpa, identified as a cosmopolitan endophyte of woody plants, G.mangiferae (Phyllostictacapitalensis).Phytopathology 92(5): 464–477. 10.1094/PHYTO.2002.92.5.46418943020 · doi ↗ · pubmed ↗
- 2Carbone I Kohn LM (1999) A method for designing primer sets for speciation studies in filamentous ascomycetes.Mycologia 91(3): 553–556. 10.1080/00275514.1999.12061051 · doi ↗
- 3Crous PW Slippers B Wingfield MJ Rheeder J Marasas WFO Philips AJL Alves A Burgess T Barber P Groenewald JZ (2006) Phylogenetic lineages in the Botryosphaeriaceae.Studies in Mycology 55: 235–253. 10.3114/sim.55.1.23518490983 PMC 2104729 · doi ↗ · pubmed ↗
- 4Crous PW Wingfield MJ Schumacher RK Summerell BA Giraldo A GenéJ Guarro J Wanasinghe DN (2014) Fungal Planet description sheets: 281–319.Persoonia 33: 212–289. 10.3767/003158514 X 68568025737601 PMC 4312934 · doi ↗ · pubmed ↗
- 5Crous PW Wingfield MJ Burgess TI St J Hardy GE Crane C Barrett S Cano-Lira JF Leroux JJ (2016) Fungal Planet description sheets: 469–557.Persoonia 37: 218–403. 10.3767/003158516 X 69449928232766 PMC 5315290 · doi ↗ · pubmed ↗
- 6Crous PW Wingfield MJ Burgess TI Carnegie AJ St J Hardy GE Smith D Summerell BA Cano-Lira JF (2017) Fungal Planet description sheets: 625–715.Persoonia 39: 270–467. 10.3767/persoonia.2017.39.1129503478 PMC 5832955 · doi ↗ · pubmed ↗
- 7Crous PW Schumacher RK Wingfield MJ Akulov A Denman S Roux J Braun U Burgess TI Carnegie AJVáczy KZ Guatimosim E Schwartsburd PB Barreto RW Hernández-Restrepo M Lombard L Groenewald JZ (2018) New and interesting fungi. 1.Fungal Systematics and Evolution 1: 169–215. 10.3114/fuse.2018.01.0832490366 PMC 7259438 · doi ↗ · pubmed ↗
- 8Crous PW Carnegie AJ Wingfield MJ Sharma R Mughini G Noordeloos ME Santini A (2019) Fungal Planet description sheets: 868–950.Persoonia 39: 291–473. 10.3767/persoonia.2019.42.11PMC 671253831551622 · doi ↗ · pubmed ↗