METTL3-dependent m6A modification of GHR mRNA regulates mitochondrial function through mitochondrial biogenesis during myoblast differentiation
Changbin Zhao, Bowen Hu, Zhijun Wang, Ze Zhang, Wen Luo, Hongmei Li, Xiquan Zhang

TL;DR
This study shows that METTL3-dependent m6A modification of GHR mRNA reduces mitochondrial function during muscle cell development by inhibiting mitochondrial biogenesis.
Contribution
The study identifies METTL3-dependent m6A modification as a novel regulator of GHR mRNA and mitochondrial biogenesis during myoblast differentiation.
Findings
METTL3-dependent m6A modification at GHR-139 down-regulates GHR mRNA and protein expression.
m6A modification impairs mitochondrial biogenesis and mitochondrial function during myoblast differentiation.
Overexpression of METTL3 alone inhibits GHR gene expression and mitochondrial biogenesis.
Abstract
N6-methyl-adenosine (m6A) methylation has recently been shown to play a critical role in muscle development. We recently revealed that local GHR knockdown impairs mitochondrial function by inhibiting mitochondrial biogenesis, thereby repressing myoblast differentiation. And we identified m6A modification peaks in the GHR mRNA of chicken muscle tissue. However, whether m6A modification may regulate GHR mRNA expression to impinge on mitochondrial function through mitochondrial biogenesis during myoblast differentiation is lagging. We first predicted three potential m6A modification sites (GHR-139, GHR-203, GHR-385) on GHR mRNA through SRAMP online prediction website. We then confirmed that GHR-139 is the METTL3-dependent m6A modification site. Further, METTL3-dependent m6A modification down-regulated the GHR mRNA and protein expression, and blunted the GHR mediated GH-GHR-IGFs axis signal…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1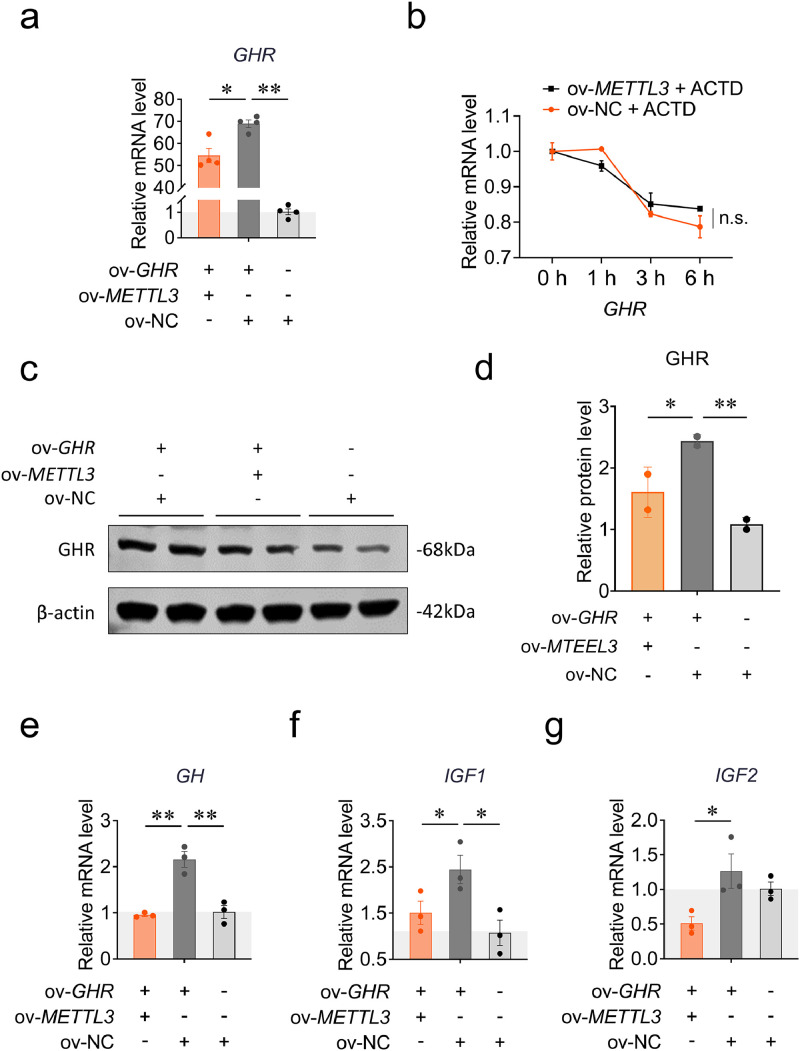 Figure 2
Figure 2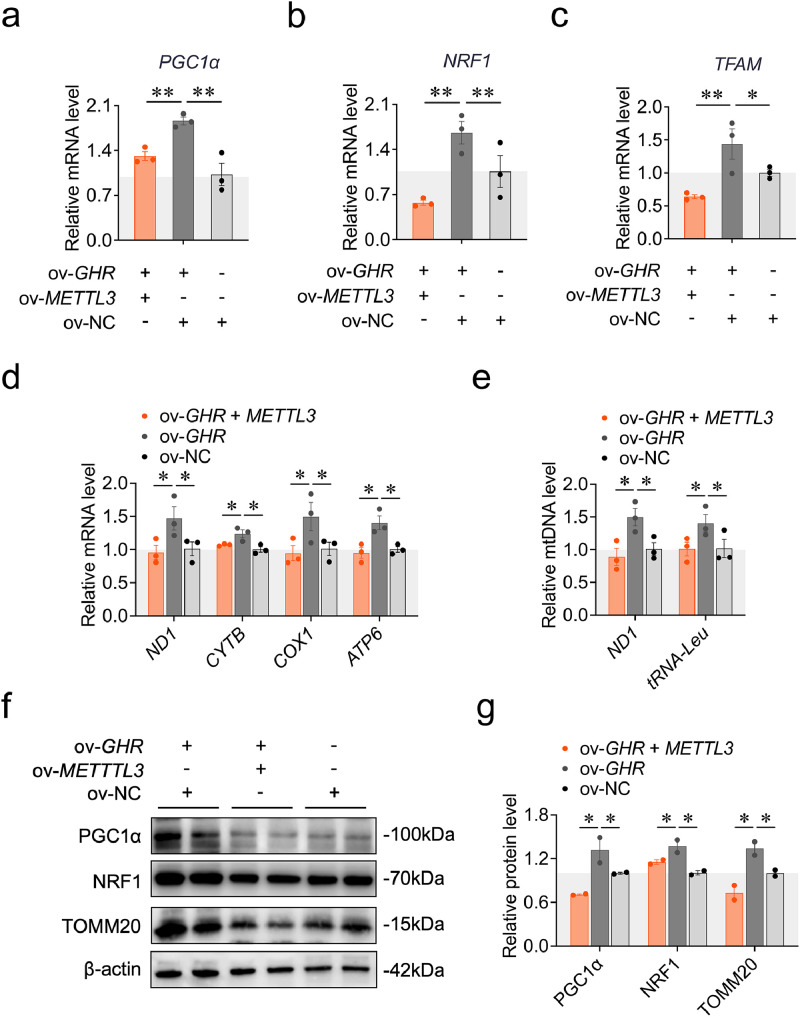 Figure 3
Figure 3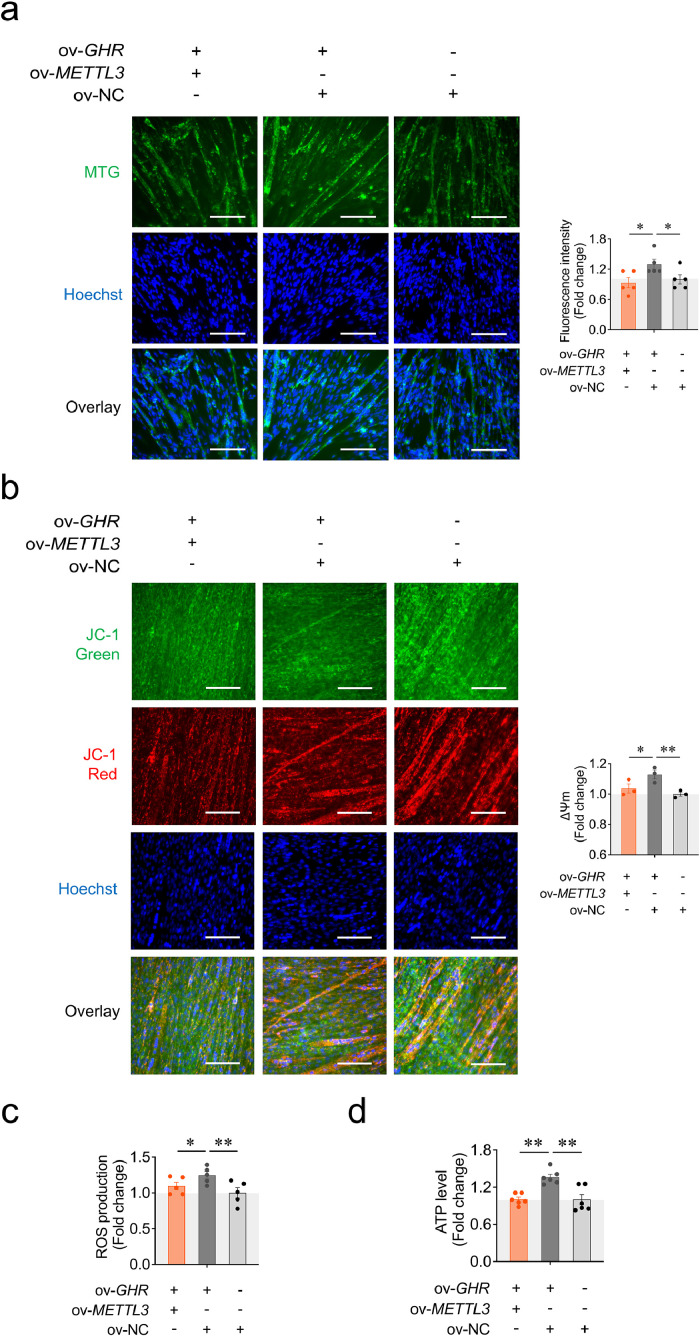 Figure 4
Figure 4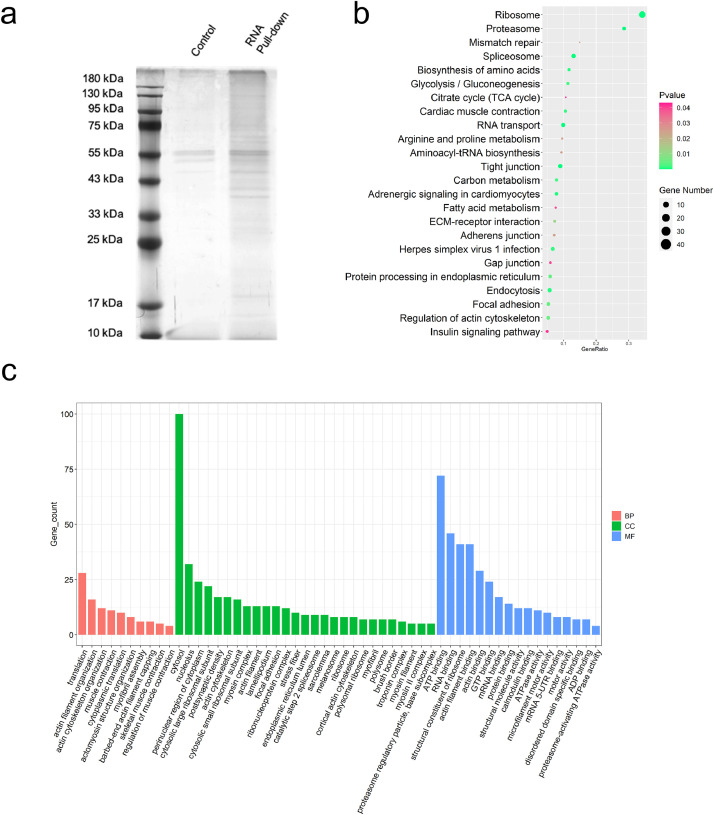 Figure 5
Figure 5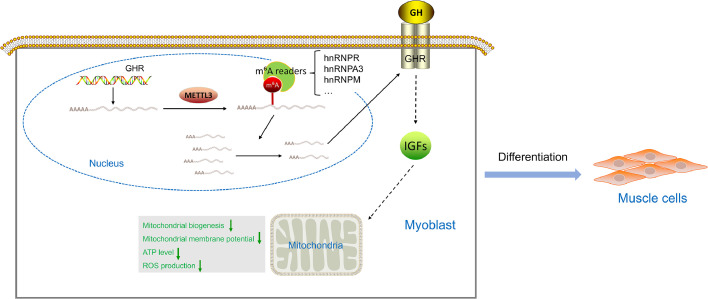 Figure 6
Figure 6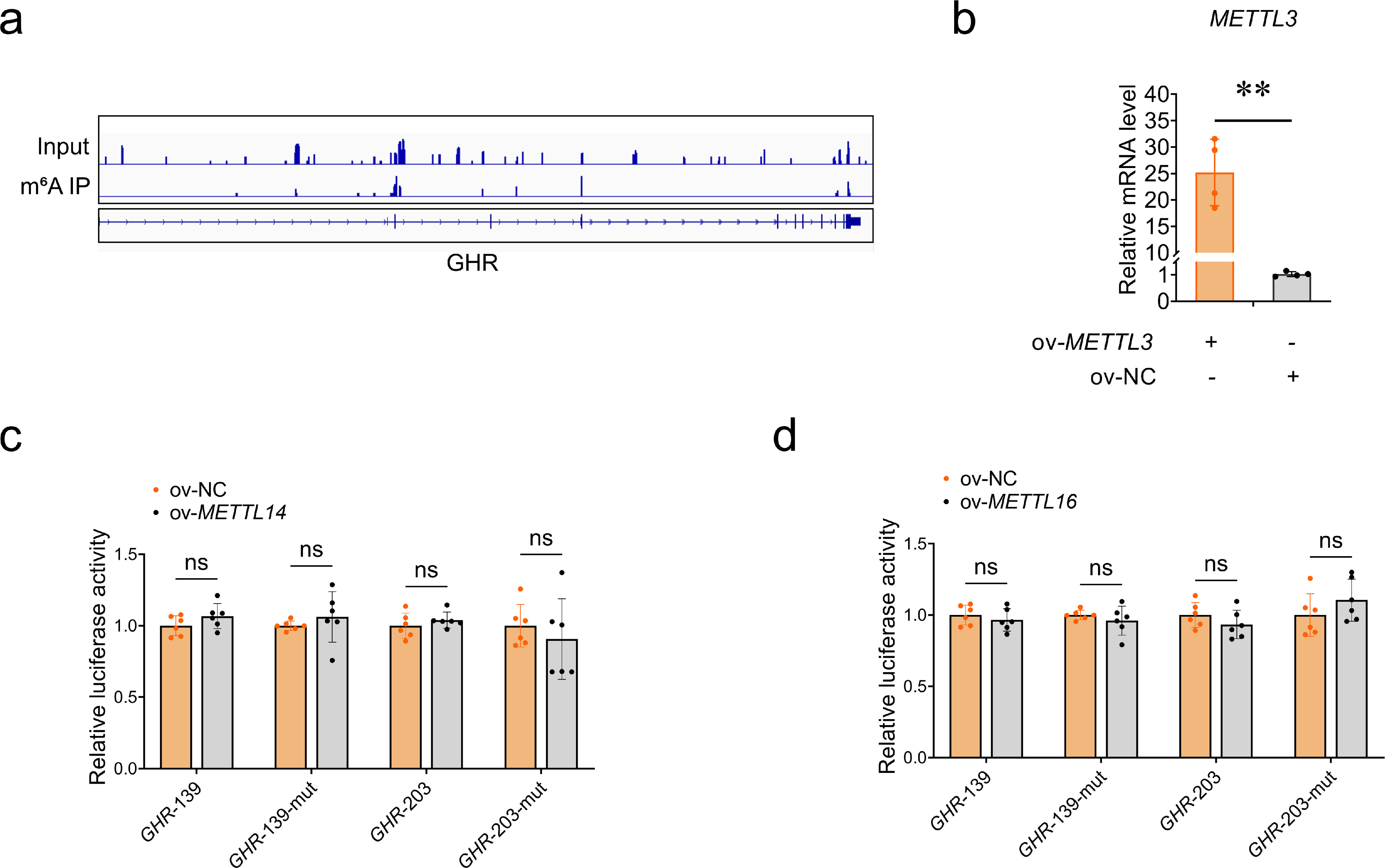 Figure 7
Figure 7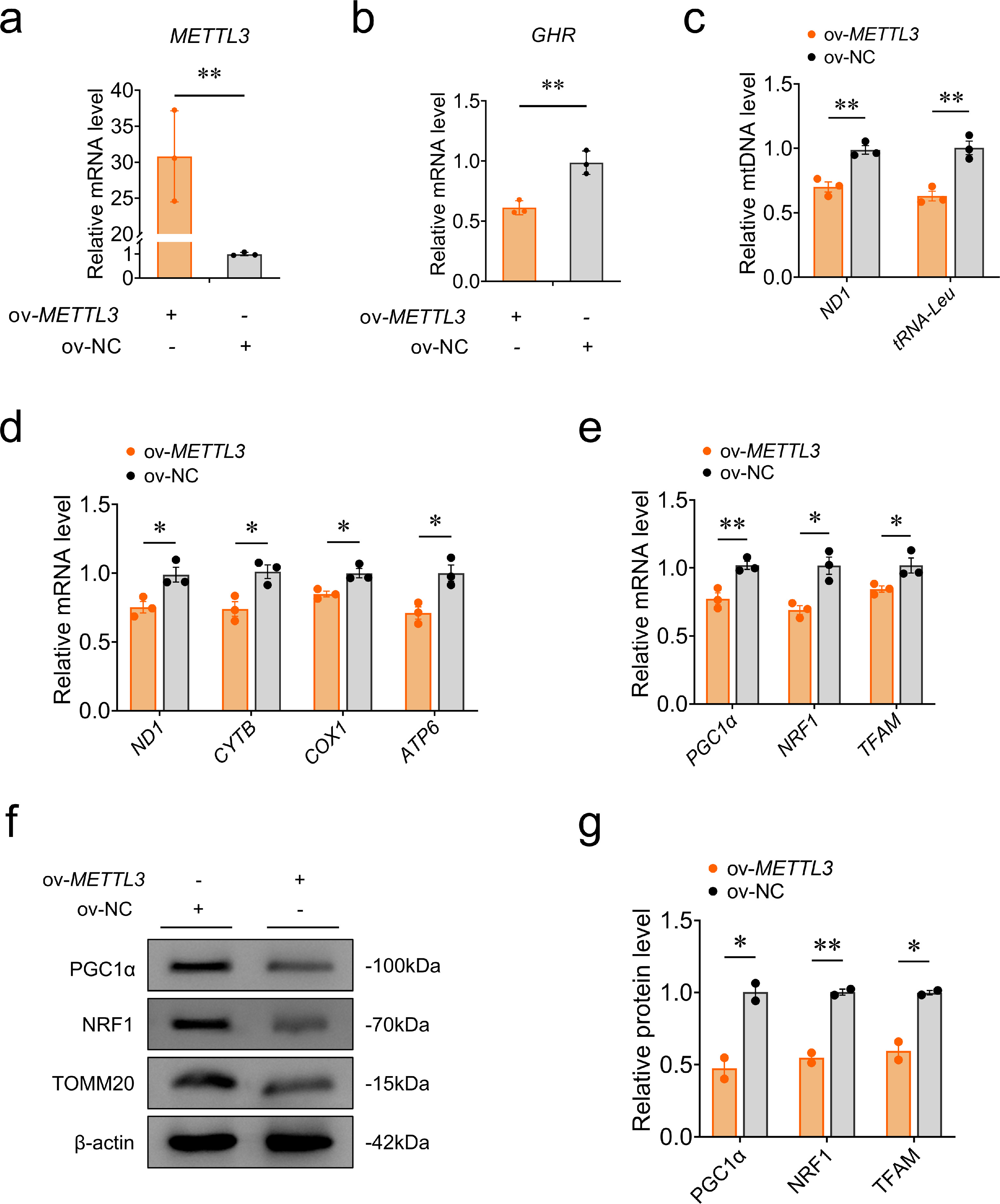 Figure 8
Figure 8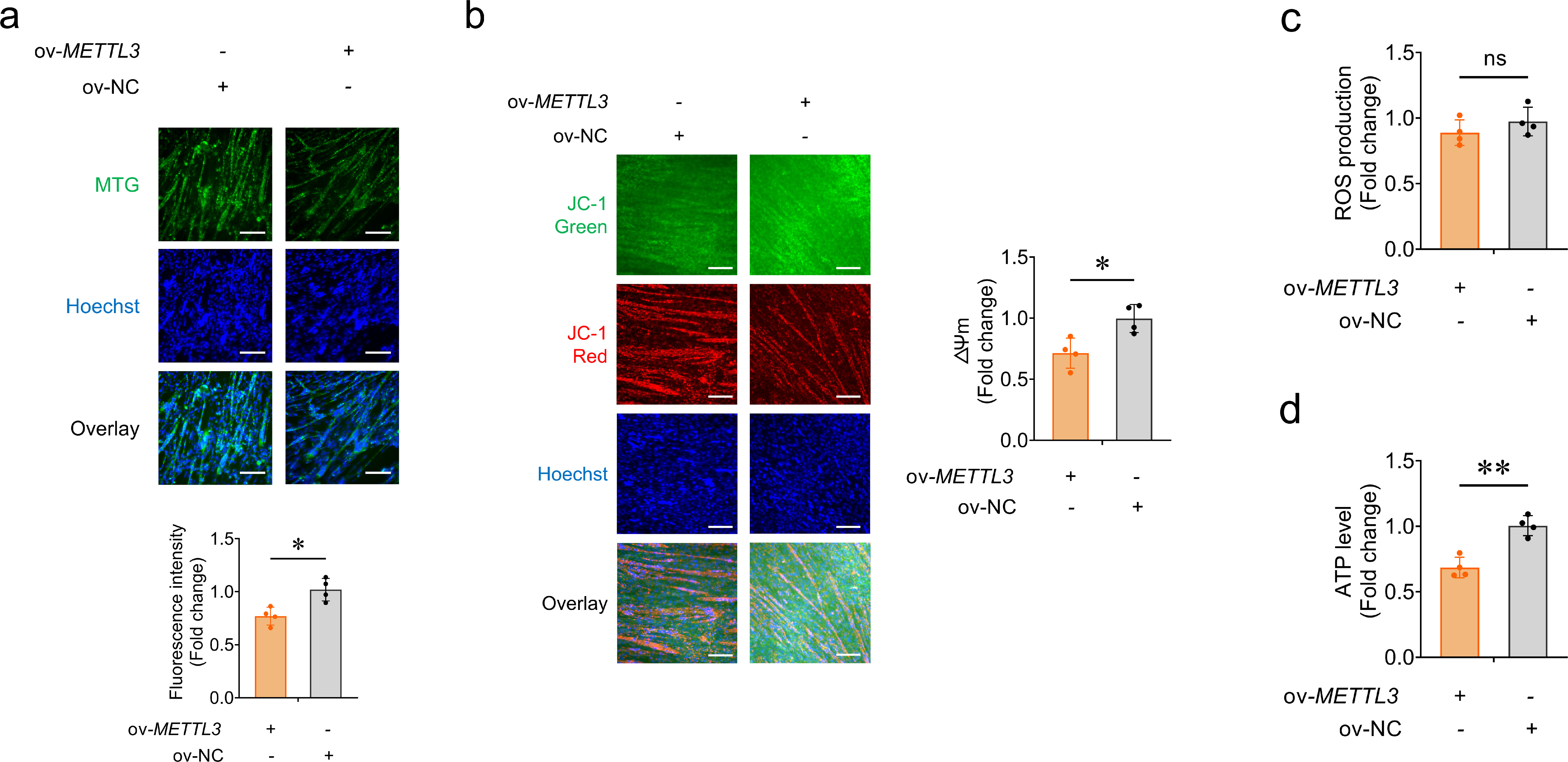 Figure 9
Figure 9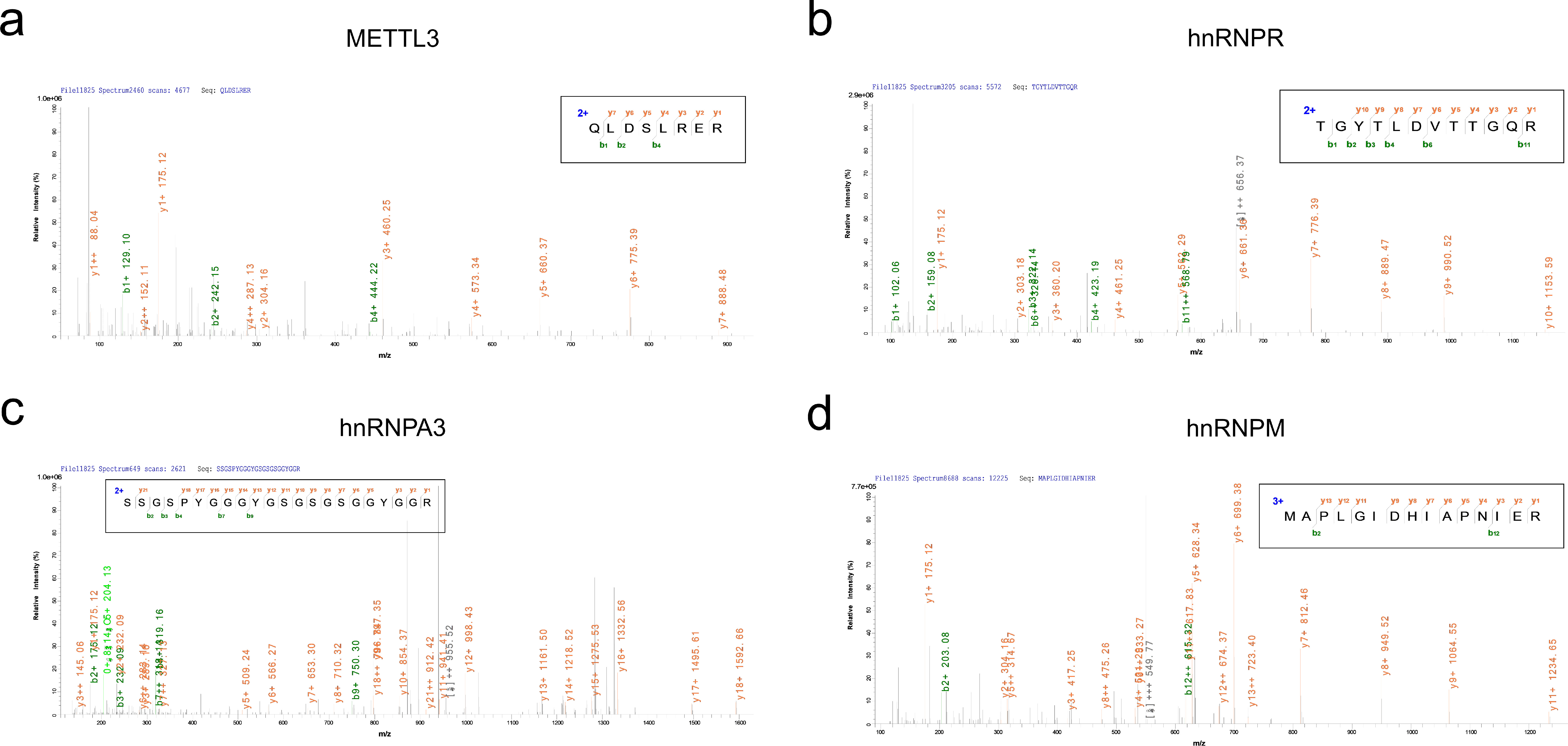 Figure 10
Figure 10Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA modifications and cancer · HVDC Systems and Fault Protection · RNA Research and Splicing
Introduction
As a crucial endocrine system regulating growth and development in the body, the growth hormone (GH)-growth hormone receptor (GHR)-insulin-like growth factor 1 (IGF1) axis plays a pivotal role in muscle development. GHR, a member of the type I cytokine receptor family, is widely expressed across various tissues and cell types (Hu et al., 2021). Previous studies have shown that GH binding to GHR activates the JAK2–STAT signaling pathway, leading to the transcription of specific genes such as IGF1 and IGF2, thereby influencing cell proliferation and differentiation (Ranke and Wit, 2018). On the other hand, mitochondria mainly generate adenosine triphosphate (ATP) through oxidative phosphorylation (OXPHOS) to provide energy for myoblast differentiation (Rahman and Quadrilatero, 2021). We recently revealed that local GHR knockdown impairs mitochondrial function by inhibiting mitochondrial biogenesis via the IGF1-PI3K/AKT/CREB pathway, thereby repressing myoblast differentiation (Hu et al., 2023). Our data indicate that local GHR acts as a control value to maintain mitochondrial function by regulating mitochondrial biogenesis during myoblast differentiation.
N6-methyladenosine (m^6^A) is a prevalent post-transcriptional RNA modification in eukaryotic mRNA transcripts, such as mRNA splicing, export, translation, and degradation (Desrosiers et al., 1975; Jia et al., 2012; Jiang et al., 2021), which impinges on manifold biological processes including muscle development (Li et al., 2021). Dou et al. (2023) investigated m^6^A modifications in the longissimus dorsi muscle of pigs at different developmental stages and identified differential modifications in the periodic gene CCND2 and the key transcription factor MYOD1, which are involved in muscle development and consequently influence muscle growth. Similarly, Feng et al. (2024) reported that METTL3-mediated m^6^A modification enhances MYOD expression, thereby promoting the activation of muscle satellite cells and skeletal muscle growth in adolescents. Moreover, m^6^A modification also influences skeletal muscle development by regulating the expression of non-coding RNAs (Mao et al., 2025; Su et al., 2025). It remains unclear whether the GH-GHR-IGF1 axis, a critical regulatory pathway for skeletal muscle development, is also affected by m^6^A modification.
The m^6^A modification requires well collaboration of methyltransferases (METTL3, METTL14 and METTL16), demethylases (FTO and ALKBH5) and methylated readers (YTHDF1/2/3, YTHDC1/2 and IGF2BP1/2/3) (Yue et al., 2015; Zaccara et al., 2019). METTL3 is the core catalytic subunit of the m^6^A methyltransferase complex that plays a pivotal catalytic role in m^6^A modification (Bokar et al., 1997). Research has found that METTL3 and IGFBP2-mediated m^6^A modifications of NPR3 and GHR may play a profound regulatory role in rheumatoid arthritis (Xiao et al., 2022). However, the regulatory role in muscles remains unclear. Our previous data has revealed that there have m^6^A-modified peaks on the growth hormone receptor (GHR) mRNA through MeRIP-Seq in the pectoralis major muscle of Xinghua and White Recessive Rock chickens (Wang et al., 2024), suggesting that m^6^A modification might regulate GHR mRNA transcript during muscle development. Therefore, we wonder if m^6^A modification may regulate GHR mRNA expression to impinge on mitochondrial function through mitochondrial biogenesis during myoblast differentiation.
Based on this, we modulated the expression of the growth hormone receptor (GHR) gene and key methyltransferase genes in primary chicken myoblasts to investigate their roles in mitochondrial biogenesis and function during myoblast differentiation. Our results demonstrate that METTL3-dependent N⁶-methyladenosine (m⁶A) modification inhibits mitochondrial biogenesis by suppressing GHR mRNA expression during myoblast differentiation, ultimately leading to impaired mitochondrial function. Understanding the precise roles of m^6^A modification in regulating GH-GHR-IGFs growth axis to impinge on mitochondrial function during myoblast differentiation may provide new insights for the cultivated meat industry and potentially contribute to the improvement of muscle developmental disorders.
Materials and methods
Ethics statement
All animal experiments were conducted in accordance with the protocols approved by the Institutional Animal Care and Use Committee of South China Agricultural University (approval number: 2023c014; taroval date: Feb 18, 2023). All procedures complied with the regulations and guidelines set by the committee and were designed to minimize animal suffering.
Cell culture
Chicken primary myoblasts (CPM) were isolated from the leg muscle of embryos on day 10 of incubation, as previously described (Luo et al., 2014). The CPM were cultured in growth medium (GM) consisting of RPMI-1640 medium (Gibco, USA), 15 % fetal bovine serum (Gibco, USA), and 0.2 % penicillin/streptomycin. When myoblasts reached 90 % confluence, the growth medium was replaced with differentiation medium (DM) including 2 % horse serum. All cells were maintained in a humidified incubator at 37°C with 5 % CO₂.
RNA extraction and quantitative real-time PCR
Total RNA was extracted from cells using RNAiso reagent (Takara, Japan) according to the manufacturer’s instructions. RNA integrity was assessed by agarose gel electrophoresis, and concentration was measured using a Nanodrop 2000c spectrophotometer (Thermo, USA). Only RNA samples meeting quality requirements were used for further analysis. cDNA was synthesized from total RNA using the PrimeScript RT reagent Kit (Takara, Japan), following the manufacturer’s protocol. Quantitative real-time PCR (qRT-PCR) was performed using the MonAmp™ ChemoHS qPCR Mix (Monad, China) on a Bio-Rad CFX96 Real-Time PCR Detection System (Bio-Rad, USA), with β-actin as the internal reference gene. Primer sequences are listed in Table S1 and were synthesized by Sangon Biotech (Shanghai, China) Co., Ltd.
DNA extraction and analysis of mtDNA copy number
Nuclear and mitochondrial DNA (mtDNA) were extracted from cells using the DNA Tissue Kit (Omega, USA), following the manufacturer’s instructions. DNA integrity was assessed via agarose gel electrophoresis, and concentration was determined using a Nanodrop 2000c spectrophotometer (Thermo, USA). Only DNA samples meeting quality criteria were used for subsequent analysis. qRT-PCR was performed using the MonAmp™ ChemoHS qPCR Mix (Monad, China) on a Bio-Rad CFX96 Real-Time PCR Detection System (Bio-Rad, USA), in accordance with the manufacturer's protocol. The relative mtDNA copy number was quantified by qRT-PCR, with each sample analyzed in duplicate. Mitochondrial genes ND1 and tRNA-Leu were used as markers of total mitochondrial content, and the nuclear gene β2M served as the internal reference. Primer sequences are listed in Table S1 and were synthesized by Sangon Biotech (Shanghai, China) Co., Ltd.
Plasmids construct
Overexpression vectors: To generate pcDNA3.1-GHR, pcDNA3.1-METTL3, pcDNA3.1-METTL14 and pcDNA3.1-METTL16, cDNA encoding GHR, METTL3, METTL14 and METTL16 was amplified by PCR using chicken cDNA as the template, the forward primer contained an EcoRI site, and the reverse primer contained an XbaI site. The amplified fragment was purified, EcoRI/XbaI-digested, and ligated into EcoRI/XbaI sites in pcDNA3.1 vectors. Promoter reporter plasmids: pmir-GLO-GHR-139, pmirGLO-GHR-139-mut, pmirGLO-GHR-203, pmirGLO-GHR-203-mut were synthesized by a commercial company (Genecreate, China). The accuracy of all plasmid constructs was confirmed by Sanger DNA sequencing.
Transfection
Cells were plated in culture plates (6 × 10^4^ cells per well of 12-well plate) and incubated overnight prior to the transfection experiment. Transfection was conducted once myoblasts reached 90 % cell confluence. To induce differentiation, the growth medium (GM) used for myoblast proliferation was replaced with differentiation medium (DM). Subsequently, the differentiating myoblasts were transfected using Lipofectamine 3000 reagent (Invitrogen, USA) according to the manufacturer’s instructions. All cells were subjected to further studies and analyses 48 h post-transfection.
Western blot analysis
Cellular proteins were extracted by adding protease inhibitor (Beyotime, China) to protein lysis buffer (Beyotime, China) at a 1:100 ratio. The lysates were centrifuged at 12,000 × g for 5 minutes at 4 °C, and the resulting supernatant containing total cellular proteins was collected. Protein concentration was measured using the BCA Protein Assay Kit (Beyotime, China) according to the manufacturer’s instructions. The proteins were then separated on a 10 % SDS-PAGE and transferred onto a PVDF membrane. Subsequently, standard procedures were followed to probe the membrane with the respective antibodies. The Western blotting included the use of the following antibodies at the indicated dilutions: anti-GHR (bs-0654R; Bioss, China; 1:500), anti-PGC1α (bs-1832R; Bioss, China; 1:500), anti-NRF1 (12482-1-AP; Proteintech, USA; 1:500), anti-TOMM20 (AF1717; Beyotime, China; 1:500), anti-β-actin (bs-0061R; Bioss, China; 1:1000), goat anti-rabbit IgG-HRP (bs-0295 G; Bioss, China; 1:5000), and goat anti-mouse IgG-HRP (bs-0296G; Bioss, China; 1:5000).
MeRIP sequencing and GHR m6A peak visualization
Three 7-week-old lean-type and obese-type broiler chickens were selected from each group, and their pectoral muscle tissue samples were collected. From each sample, 300 μg of RNA was extracted. After fragmentation, samples from each group underwent enrichment using an m^6^A antibody (Synaptic Systems, Göttingen, Germany) and non-enriched input processing. Subsequently, libraries were constructed for both the immunoprecipitated mRNA (IP group) and untreated mRNA (Input group) using the NEBNext UltraTM RNA Library Prep Kit (NEB, Ipswich, USA). Sequencing was performed on the Illumina HiSeq platform using standard protocols (paired-end reads, 150 bp in length). The raw sequencing data have been deposited in the CNCB (GSA database, accession number CRA008607). Based on the raw data, the m^6^A peak distribution of the GHR gene in both the IP and Input groups was visualized using Integrative Genomics Viewer (IGV, Broad Institute, Cambridge, USA).
Methylated RNA immunoprecipitation assay
Methylated RNA immunoprecipitation (MeRIP) assay was performed as previously described (Li et al., 2022). In brief, RNA was first incubated with RNA fragmentation buffer in a thermal cycler at 70°C for 5 min. m^6^A antibody (CST, USA) was incubated with pre-washed protein A/G magnetic beads (MCE, China) overnight at 4°C. Second, the magnetic bead-antibody mixture was separated using a magnetic separation rack and washed twice with IP buffer. The mixture was resuspended in 500 μL of reaction solution and incubated at 4°C for 6-8 h. Third, the magnetic bead-antibody RNA mixture was washed twice in IP buffer, low-salt solution and high-salt solution at 4°C for 5 min each. The m^6^A-enriched RNA was eluted and purified using the RNeasy MinElute Cleanup Kit (Qiagen, Germany). Finally, the m^6^A-enriched RNA was validated through reverse transcription and PCR experiments, primers were shown in Table S2.
RNA pull-down assay
Plasmids expressing full-length GHR mRNA containing F2 tagged and control vector were transfected into CPM. A F2 RNA-Protein Pull-Down Kit (FITGENE, China) was used in RNA-protein pull-down experiments according to the manufacturer’s instructions.
Protein sample preparation and LC-MS/MS
Protein samples were initially treated for reduction by adding 4 µL of 0.05 M Tris(2-carboxyethyl) phosphine (Sigma, USA) and incubating at 60°C for 1 h. Subsequently, alkylation was achieved by adding 2 µL of 55 mM S-methyl methanethiosulfonate (MMTS, Sigma) followed by incubation in darkness at room temperature for 45 minutes. Sample cleanup and buffer exchange were performed using 10 kDa cut-off ultrafiltration tubes. After initial centrifugation at 12,000 g for 20 minutes, the samples were washed twice with 100 µL of UA buffer (8 M urea, pH 8.5, Sigma) and then three times with 100 µL of 0.25 M Triethylammonium bicarbonate (TEAB, Sigma), with each wash step followed by centrifugation at 12,000 g for 20 minutes. For enzymatic digestion, 50 µL of 0.5 M TEAB containing trypsin (Promega, USA) was added at an enzyme-to-protein mass ratio of 1:50. This mixture was incubated overnight (12 h) at 37°C. The following day, digestion was continued by adding trypsin at a 1:100 mass ratio and incubating for an additional 4 h at 37°C. The resulting peptide mixture was transferred to a fresh collection tube, and the filtrate containing the peptides was collected via centrifugation. The collected peptides were then dried using low-temperature vacuum evaporation.
Dried peptide fractions were reconstituted in loading buffer (0.1 % formic acid (Sigma), 2 % acetonitrile (Fisher, USA)) for liquid chromatography-tandem mass spectrometry (LC-MS/MS) analysis. Peptide separation was carried out on a Dionex Ultimate 3000 RSLCnano system (Fisher) equipped with a C18 reversed-phase column (150 mm length, 75 µm inner diameter, 3 µm resin). A binary solvent system consisting of mobile phase A (0.1 % formic acid in water) and mobile phase B (0.1 % formic acid in 80 % ACN) was used. Peptides were eluted over 65 minutes using a linear gradient from 5 % to 50 % mobile phase B at a flow rate of 300 nL/min. The LC eluate was directly analyzed using a Q Exactive Hybrid Quadrupole-Orbitrap mass spectrometer (Fisher) operating in information-dependent acquisition (IDA) mode. Full MS scans (MS1) were acquired over the m/z range of 350-1,800 at a resolution of 70,000, with a maximum ion accumulation time of 250 ms. From each MS1 scan, up to 20 of the most intense precursor ions were selected for fragmentation via higher-energy collisional dissociation (HCD) using a normalized collision energy setting of 30. Tandem mass spectra (MS/MS) were acquired at a resolution greater than 17,500, with a minimum precursor accumulation time of 100 ms and dynamic exclusion enabled for 20 seconds. The raw data have been deposited in the CNCB and are accessible through OMIX series accession number OMIX009631.
Protein identification and bioinformatics analysis
The raw mass spectrometry data were acquired in RAW format and converted into MGF format using Proteome Discoverer 1.4 (version 1.4.0.288, Fisher). The MGF file and the protein database (https://www.uniprot.org/taxonomy/9031) were input into ProteinPilot™ Software 4.5 (version 1656, SCIEX, USA) for mass spectrometry identification. After subtracting the control group's data from the identified protein data of the experimental group, the remaining differential proteins were considered the target binding proteins. Gene Ontology (GO, http://geneontology.org) and Kyoto Encyclopedia of Genes and Genomes (KEGG, http://www.kegg.jp) were used to annotate all identified proteins. GO and KEGG enrichment analyses were conducted using Fisher's exact test, with FDR correction applied for multiple comparisons. Enriched GO terms and KEGG pathways were considered statistically significant at p < 0.05.
Mito-tracker green staining and hoechst 33342 staining
Mitochondria were stained using Mito-Tracker Green (MTG) (Beyotime, China), and nuclei were counterstained with Hoechst 33342 (Beyotime, China). At 48 h post-transfection, cells were rinsed twice with PBS and incubated with MTG for 30 minutes. After two additional PBS washes, cells were incubated with Hoechst 33342 for 10 minutes, followed by two final rinses with PBS. Fluorescence images were captured from five randomly selected fields per sample using a TE2000-U fluorescence microscope (Nikon, Japan), and image analysis was performed using NIS-Elements software.
Detection of reactive oxygen species
Intracellular reactive oxygen species (ROS) levels were measured using the ROS Assay Kit (Beyotime, China) following the manufacturer’s instructions. The fluorescence intensity of dichlorofluorescin (DCF) was quantified using a multifunctional microplate reader (BioTek, USA) to evaluate ROS production in myoblasts.
Detection of ATP content
ATP levels in myoblasts were measured using an ATP Detection Kit (Beyotime, China) following the manufacturer’s instructions. The luminescence intensity was detected using a multifunctional microplate reader (BioTek, USA) to quantify intracellular ATP content.
Detection of mitochondrial membrane potential
Mitochondrial membrane potential (ΔΨm) in myoblasts was assessed using the JC-1 Assay Kit (Beyotime, China), following the manufacturer’s instructions. For each sample, five randomly selected fields were imaged using a TE2000-U fluorescence microscope (Nikon, Japan). The captured images were subsequently analyzed using NIS-Elements software.
Statistical analysis
All experiments were performed in at least three independent replicates. Graphical representations were generated using GraphPad Prism v9.0 (GraphPad Software, USA). Data are presented as mean ± standard error of the mean (SEM). Statistical analyses were conducted using Student’s t-test. A p-value of < 0.05 was considered statistically significant. Significance levels are indicated as follows: *p < 0.05, **p < 0.01, ***p < 0.001, and ns indicates no significance.
Results
Verification of potential methyltransferase and m6A modification sites on GHR mRNA in chicken primary myoblasts (CPM)
In previous studies, we identified m^6^A modification peaks on GHR mRNA in the pectoralis muscle of chickens through MeRIP-Seq (Fig. S1a), suggesting that m^6^A modification of GHR mRNA may play a significant role in muscle development. We first explored potential m^6^A modification sites on GHR mRNA by the online prediction SRAMP prediction server (http://www.cuilab.cn/sramp). Three potential m^6^A modification sites, GHR-139, GHR-203, and GHR-385, were predicted on GHR mRNA (Fig. 1a). To confirm the locations of m^6^A modification sites, METTL3 was overexpressed in differentiating myoblasts, with high overexpression efficiency verified (Fig. S1b), followed by MeRIP-PCR to validate the predicted sites. Gel electrophoresis revealed that, following m^6^A immunoprecipitation (IP), the PCR-amplified fragment containing the GHR-139 and GHR-203 sites exhibited a distinct 304 bp band (Fig. 1b), indicating specific enrichment of this region and the presence of m^6^A modifications. In contrast, no corresponding band was observed for the amplified fragment containing GHR-385 after m^6^A IP compared to the input group (Fig. 1c), suggesting that this region was not enriched and did not contain m^6^A modification sites. To further investigate the m^6^A modification enzymes of GHR mRNA and their specific modification sites, we constructed four pmirGLO vectors containing either the wild-type or mutated versions of GHR-139 and GHR-203, and co-transfected them with METTL3, METTL14, and METTL16 in CPM (Fig. 1d). The dual fluorescence assay revealed that METTL3 overexpression significantly inhibited the expression of the wild-type GHR-139 reporter gene but had no effect on the mutant GHR-139 reporter gene. METTL3 overexpression did not affect the expression of either the wild-type or mutant GHR-203 (Fig. 1e). Moreover, overexpression of METTL14 and METTL16 had no impact on either the wild-type or mutant forms of GHR-139 and GHR-203 (Fig. S1c, d). These results indicate that the regulation of GHR expression levels might rely on the METTL3-dependent m^6^A modification at the GHR-139 site during myoblast differentiation.Fig. 1. Verification of potential methyltransferase and m^6^A modification sites on GHR mRNA in CPM. (a) Prediction of potential m^6^A modification sites on GHR mRNA through online prediction SRAMP prediction server (http://www.cuilab.cn/sramp). (b) MeRIP-PCR assays were applied to assess the m^6^A methylation modification sites on GHR-139 and GHR-203 after METTL3 overexpression during myoblast differentiation. (c) MeRIP-PCR assays were applied to assess the m^6^A methylation modification sites on GHR-385 after METTL3 overexpression during myoblast differentiation. (d, e) Relative luciferase activities of CPM co-transfected with plasmids containing wild-type or mutant GHR mRNA and METTL3 cDNA. Firefly luciferase activities were measured and normalized to renilla luciferase activity (n = 6). Data are shown as mean ± SEM, ***p < 0.001, ns means no significant.Fig 1
METTL3-dependent m6A modification regulates GHR mRNA and protein expression, and the mRNA expression involved in GH-GHR-IGFs axis during CPM differentiation
To further investigate the METTL3-dependent m^6^A modification roles in the regulation of GHR mRNA, we co-transfected pcDNA3.1-METTL3 and pcDNA3.1-GHR during CPM differentiation. The GHR mRNA and protein expression were all decreased in the co-transfection group compared with the GHR overexpression group (Fig. 2a, c, d). In addition, we successfully overexpressed the METTL3 gene in CPM (Fig. S2a), and found that this significantly inhibited GHR expression (Fig. S2b). We then determined the stability of GHR mRNA by actinomycin D (ACTD). The GHR mRNA expression did not change significantly in the co-transfection group compared with the GHR overexpression group (Fig. 2b), indicating that METTL3-dependent m^6^A modification did not influence the stability of GHR mRNA during myoblast differentiation. On the other hand, we determined the mRNA expression involved in the GH-GHR-IGFs axis by RT-qPCR. The GH, IGF1 and IGF2 expression were all significantly down-regulated in the co-transfection group compared with the GHR overexpression group (Fig. 2e-f). Taken together, these results indicate that METTL3-dependent m^6^A modification suppresses the GHR mRNA and protein expression, and blunts the GHR-mediated GH-GHR-IGFs signaling during myoblast differentiation.Fig. 2METTL3-dependent m^6^A modification regulates GHR mRNA and protein expression, and the mRNA expression involved in the GH-GHR-IGFs axis during CPM differentiation. (a) The GHR mRNA expression was measured by RT-qPCR at 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR and pcDNA3.1 (n = 4). (b) The RNA stability of GHR mRNA was detected by RT-qPCR with METTL3 overexpression treated with ACTD during myoblast differentiation (n = 3). (c, d) The GHR protein expression was measured by western blots at 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR and pcDNA3.1 (n = 2). (e) The GH mRNA expression was measured by RT-qPCR at 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR and pcDNA3.1 (n = 3). (f) The IGF1 mRNA expression was measured by RT-qPCR at 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR and pcDNA3.1 (n = 3). (g) The IGF2 mRNA expression was measured by RT-qPCR at 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR and pcDNA3.1 (n = 3); Data are shown as mean ± SEM, *p < 0.05, **p < 0.01.Fig 2
METTL3-dependent m6A modification of GHR mRNA regulates the mRNA and protein expression involved in mitochondrial biogenesis during CPM differentiation
We asked whether METTL3-dependent m^6^A modification of GHR mRNA regulates mitochondrial biogenesis during myoblast differentiation. We first examined the expression of mitochondrial biogenesis markers, including PGC1α, NRF1, and TFAM, following co-transfection in CPM. The PGC1α, NRF1 and TFAM expression were significantly down-regulated in the co-transfection group compared with the GHR overexpression group (Fig. 3a-c). Consistently, single METTL3 overexpression significantly inhibited PGC1α, NRF1 and TFAM expression (Fig. S2e). We then investigated the effects of METTL3-dependent m^6^A modification of GHR mRNA on mtDNA transcription and replication. The expression levels of mtDNA-encoded genes (ND1, CYTB, COX1, ATP6) and mtDNA copy number (indicated by ND1 and tRNA-Leu) were significantly decreased in the co-transfection group compared to the GHR overexpression group (Fig. 3d, e). Similar results were observed upon METTL3 overexpression alone (Fig. S2c, d). Finally, we assessed the protein levels of mitochondrial biogenesis marker genes. The PGC1α, NRF1 and TOMM20 protein levels were reduced in the co-transfection group compared with the GHR overexpression group (Fig. 3f, g), similarly to the results observed with METTL3 overexpression (Fig. S2f, g). Collectively, these findings suggest that METTL3-mediated m⁶A modification of GHR mRNA suppresses mitochondrial biogenesis during myoblast differentiation.Fig. 3METTL3-dependent m^6^A modification of GHR mRNA regulates the mRNA and protein expression involved in mitochondrial biogenesis during CPM differentiation. (a) The PGC1α mRNA expression was measured by RT-qPCR at 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR and pcDNA3.1 (n = 3). (b) The NRF1 mRNA expression was measured by RT-qPCR at 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR and pcDNA3.1 (n = 3). (c) The TFAM mRNA expression was measured by RT-qPCR at 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR and pcDNA3.1 (n = 3). (d) The mtDNA transcription was measured by RT-qPCR at 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR and pcDNA3.1 (n = 3). (e) The mtDNA copy number was measured by qPCR at 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR and pcDNA3.1 (n = 3). (f, g) The PGC1α, NRF1, TOMM20 protein expression was measured by western blots at 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR and pcDNA3.1 (n = 2). Data are shown as mean ± SEM, *p < 0.05, **p < 0.01.Fig 3
METTL3-dependent m6A modification of GHR mRNA regulates mitochondrial function during CPM differentiation
Mitochondrial biogenesis is essential for maintaining normal mitochondrial oxidative phosphorylation (OXPHOS), raising the question of whether METTL3-mediated m⁶A modification of GHR mRNA affects mitochondrial function during CPM differentiation. To address this, we first assessed mitochondrial mass using MTG staining. A reduction in mitochondrial mass was observed in the co-transfection group compared to the GHR overexpression group (Fig. 4a). We then evaluated mitochondrial function by measuring mitochondrial membrane potential (ΔΨm), intracellular ATP levels (via a luminescence-based assay), and ROS production. The ΔΨm and ATP level as well as ROS production were all significantly decreased in the co-transfection group compared with the GHR overexpression group (Fig. 4b-d). Similarly, overexpression of METTL3 alone reduced the mitochondrial mass of CPM, and decreased the ΔΨm and ATP level, but had no effect on ROS production (Fig. S3a-d). These results suggest that METTL3-dependent m^6^A modification of GHR mRNA impairs mitochondrial function during myoblast differentiation.Fig. 4METTL3-dependent m^6^A modification of GHR mRNA regulates mitochondrial function during CPM differentiation. (a) Mitochondrial mass was measured by the fluorescence of MitoTracker-Green (MTG) at 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR and pcDNA3.1 (n = 5), bar 100 μm. (b) Mitochondrial membrane potential (ΔΨm) was assessed by JC-1 fluorescence 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR, and pcDNA3.1 (n = 3). Scale bar: 100 μm. (c) Reactive oxygen species (ROS) production was measured by DCF fluorescence 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR, and pcDNA3.1 (n = 5). (d) ATP levels were measured 48 h after co-transfection with pcDNA3.1-METTL3, pcDNA3.1-GHR, and pcDNA3.1 (n = 6). Data are shown as mean ± SEM, *p < 0.05, **p < 0.01.Fig 4
Exploration of potential m6A reader protein related to METTL3-dependent m6A modification of GHR mRNA during CPM differentiation
We next attempted to uncover the potential m^6^A reader proteins that bind to GHR mRNA by RNA pull-down coupled with mass spectrometry experiments (Fig. 5a). A total of 435 proteins were found to specifically bind to GHR mRNA (Table S3). The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis revealed that these specific proteins were mainly enriched in the ribosome, spliceosome, tight junction, RNA transport and endocytosis (Fig. 5b). Meanwhile, the gene ontology (GO) functional analysis revealed that these specific proteins enriched biological processes mainly in translation and muscle contraction; molecular functions were mainly in ATP binding and RNA binding; cell components were mainly in cytosol and nucleolus (Fig. 5c). Among these, METTL3 along with three methylated reader proteins hnRNPR, hnRNPA3 and hnRNPM were found to be precipitated (Fig. S4a-d). Taken together, these results suggest that hnRNPR, hnRNPA3 and hnRNPM might be potential m^6^A reader proteins involved in METTL3-dependent m^6^A modification of GHR mRNA during myoblast differentiation.Fig. 5. Exploration of potential reader protein involved in METTL3-dependent m^6^A modification of GHR mRNA during CPM differentiation. (a) Silver staining revealed the total protein precipitated after GHR mRNA pull-down. (b) KEGG pathways analysis of differentially expressed protein between GHR mRNA pull-down group and control group. (c) GO functions analysis of differentially expressed protein between GHR mRNA pull-down group and control group. BP: Biological Process, CC: Cellular Component, MF: Molecular Function.Fig 5
Discussion
The GH-GHR-IGF1 axis has been studied for over a century, with research primarily focused on the treatment of growth-related disorders (Ranke and Wit, 2018). In our previous research, we demonstrated that GHR-IGF1 regulates mitochondrial function and biogenesis via the PI3K/AKT/CREB pathway, thereby promoting myogenic differentiation (Hu et al., 2023). As the central hub of cellular energy metabolism, mitochondria play a vital role in myoblast differentiation and muscle development. In the early stages of myogenic differentiation, cells first remove dysfunctional mitochondria through mitophagy, followed by the induction of mitochondrial biogenesis to produce a large population of new mitochondria that meet the heightened energy demands (Sin et al., 2016). PGC1α, a key regulator of mitochondrial biogenesis, exhibits a pronounced role in promoting myogenic differentiation in skeletal muscle (Beltrà et al., 2022). Furthermore, mitochondrial dysfunction is closely associated with muscle aging and the progression of Duchenne muscular dystrophy (Pääsuke et al., 2016; Vila et al., 2017). We revealed that METTL3-dependent m^6^A modification down-regulates GHR mRNA expression to impair mitochondrial function by inhibiting mitochondrial biogenesis during myoblast differentiation. This disruption may ultimately hinder the differentiation process.
RNA post-transcriptional modifications of RNA are widely present in eukaryotic genomes and play an important role in many biological processes such as development, reproduction and disease (Wu et al., 2018; Tong et al., 2018; Cheng et al., 2021). To date, more than 100 chemical modifications have been identified in the cellular RNA of eukaryotic genomes, including N6-methyladenosine (m^6^A), N7-methylguanine (m^7^G) and 5-methylcytosine (m^5^C). While m^6^A modification is the most common RNA modification in eukaryotes which has been reported to play a pivotal role in animal muscle development (Li et al., 2021). In poultry, cycloleucine consistently reduces the m^6^A modification levels in myoblast cells, thereby inhibiting myoblast proliferation and suppressing myotube formation (Wang et al., 2022b). Conversely, low betaine concentrations increase the level of m^6^A modification, promoting myotube formation but inhibiting cell proliferation in vitro (Wang et al., 2022a). These results indicate that m^6^A modification is associated with chicken muscle development. However, the relationship between m⁶A modification and muscle development is complex. The m^6^A modification is modified by the m^6^A methyltransferases, such as METTL3, METTL14, and METTL16 (Jiang et al., 2021). Studies have shown that METTL3 promotes the myogenic differentiation of goat skeletal muscle satellite cells by regulating the expression of MEF2C (Zhao et al., 2023). In avian species, METTL14 facilitates the differentiation of duck embryonic muscle cells by modulating the expression of miR-133b (Jiang et al., 2025). In contrast, METTL16 promotes the proliferation of chicken myoblasts but inhibits their differentiation (Liu et al., 2024a), which may be due to differences in the target genes modified by distinct methyltransferases. On the other hand, the GH-GHR-IGF1 axis in the liver has been demonstrated to exhibit great complexity in response to muscle development. There is evidence that IGFBP2 and METTL3 were identified as key factors regulating m^6^A of NPR3 and GHR predicted by machine learning in synovial fibroblasts (Xiao et al., 2022). GHR has direct interaction with the m^6^A protein demonstrated by the m^6^A RIP-qRT-PCR experiment in gastric cancer cell lines (Jiang et al., 2022). In this study, we identified that the m⁶A modification site GHR-139 on GHR mRNA is regulated by METTL3, consistent with findings from previous research. Specifically, METTL3-mediated m⁶A modification influences mitochondrial function by modulating GHR expression in myoblasts. Previous studies have found that METTL3 and YTHDF2 can synergistically modify PGC-1α mRNA, mediating its degradation, thereby impairing mitochondrial function and enhancing inflammatory responses (Zhang et al., 2021). Most existing research on m⁶A modification and muscle development focuses on direct gene regulatory mechanisms, with limited attention to mitochondrial function (Yu et al., 2022). Our findings provide a new research avenue for elucidating the role of METTL3-mediated m^6^A modification in muscle development. This insight provides a theoretical basis for developing RNA m⁶A-targeted strategies to restore mitochondrial function and promote muscle growth. However, it is important to note that this study focuses exclusively on METTL3-mediated m⁶A modification of GHR. Further investigation is required to determine whether METTL3 also affects mitochondrial function through additional pathways during myogenic differentiation.
The results of m^6^A modification are intricate and variable which are determined by the dynamic balance of m^6^A-specific reader proteins and m^6^A modified mRNAs (Wang et al., 2014; Zaccara et al., 2019). At present, numerous m^6^A-specific reading proteins have been revealed to regulate m^6^A modification in different ways, including YTHDF1/2/3 (Ye et al., 2020), YTHDC1/2 (Xiao et al., 2016; Shi et al., 2017; Kretschmer et al., 2018; Fei et al., 2020), hnRNPA2B1 and hnRNPC (Yue et al., 2015; Wang et al., 2015), IGF2BP1/2/3 (Huang et al., 2018) as well as eIF3 (Zaccara et al., 2019). Generally, YTHDF1/2/3, IGF2BP1/2/3 and eIF3 regulate mRNA stability and translation, YTHDC1/2 regulates mRNA export, as well as hnRNPA2B1 and hnRNPC regulate primary microRNA processing and mRNA splicing. In this study, we revealed that METTL3-dependent m^6^A modification down-regulated GHR mRNA and protein expression without impinging on its stability by RT-qPCR, Western blots and ACTD experiments. According to the above results, we then uncovered three m^6^A reader proteins hnRNPR, hnRNPA3 and hnRNPM precipitated by RNA pull-down experiments.
HnRNPs mainly play their roles in transcriptional and post-transcriptional regulation of gene expression, including RNA splicing, polyadenylation, capping and translation (Keene, 2007; Glisovic et al., 2008). Each hnRNP family member contains at least one RNA binding domain (RBD), such as an RNA recognition motif (RRM), a K-homology (KH) domain or an arginine/glycine-rich domain (He and Smith, 2009). hnRNPA2B1 has been reported to recognize m^6^A modification sites of mRNAs to progress post-transcriptional modification resemble alternative splicing, or m^6^A modified miRNA sequence to influence primary miRNA processing (Alarcón et al., 2015). While m^6^A modification also influences the RNA secondary structure, hnRNPC has been demonstrated to recognize m^6^A modification sites to regulate mRNA abundance and splicing, which is called "the m^6^A-switch" (Liu et al., 2015). Studies have shown that hnRNPR and hnRNPA2B1 bind to and stabilize ASCL1 mRNA in a m⁶A-dependent manner, thereby promoting the progression of neuroblastoma (Hu et al., 2024). In addition, hnRNPM appears to play a critical role in the regulation of m⁶A-mediated alternative splicing of HO-1 mRNA (Su et al., 2024). Similarly, hnRNPA3 may function as a m⁶A reader protein that recognizes and modulates the alternative splicing of the oncogenic fusion gene AML1/ETO pre-mRNA, thus contributing to the progression of acute myeloid leukemia (Liu et al., 2024b). The roles of these three m⁶A reader proteins in muscle development have not been reported in previous studies. Based on our findings, we hypothesize that m⁶A modification may influence GHR mRNA expression by regulating its splicing via hnRNP proteins. However, the specific molecular mechanisms underlying this regulation require further validation.
Conclusion
In conclusion, our data corroborate that METTL3-dependent m^6^A modification down-regulated GHR mRNA expression to impair mitochondrial function by inhibiting mitochondrial biogenesis during myoblast differentiation (Fig. 6). We conducted an in-depth investigation into the relationship between m^6^A-modified GHR mRNA and mitochondrial function during myogenic differentiation, offering new insights into m^6^A modification and muscle development. This study also identifies potential molecular targets for treating muscular diseases associated with mitochondrial dysfunction. In the field of poultry, this discovery lays a theoretical foundation and supports the selection and breeding of desirable meat traits in chickens.Fig. 6. The schematic diagram illustrates the mechanisms of METTL3-dependent m^6^A modification in GHR mRNA regulation during myoblast differentiation in chickens.Fig 6
Declaration of competing interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alarcón C.R.Goodarzi H..Lee H.Liu X.Tavazoie S.Tavazoie S.F.HNRNPA 2B 1 Is a mediator of m 6A-dependent nuclear RNA processing events Cell 1622015129913082632168010.1016/j.cell.2015.08.011PMC 4673968 · doi ↗ · pubmed ↗
- 2BeltràM.Pin F.Costamagna D.Duelen R.Renzini A.BallaròR.Garcia-Castillo L.Iannuzzi A.Moresi V.Coletti D.Sampaolesi M.Penna F.Costelli P.PGC-1α in the myofibers regulates the balance between myogenic and adipogenic progenitors affecting muscle regenerationi Science 25202210.1016/j.isci.2022.105480 PMC 966388636388980 · doi ↗ · pubmed ↗
- 3Bokar J.A.Shambaugh M..E.Polayes D.Matera A.G.Rottman F.M.Purification and c DNA cloning of the Ado Met-binding subunit of the human m RNA (N 6-adenosine)-methyltransferase RNA 31997123312479409616 PMC 1369564 · pubmed ↗
- 4Cheng B.Leng L.Li Z.Wang W.Jing Y.Li Y.Wang N.Li H.Wang S.Profiling of RNA N 6-methyladenosine methylation reveals the critical role of m 6A in chicken adipose deposition Front. Cell Dev. Biol.9202111410.3389/fcell.2021.590468 PMC 789297433614638 · doi ↗ · pubmed ↗
- 5Desrosiers R.C.Friderici K..H.Rottman F.M.Characterization of Novikoff hepatoma mrna methylation and heterogeneity in the methylated 5′ terminus Biochemistry 1419754367437416989310.1021/bi 00691 a 004 · doi ↗ · pubmed ↗
- 6Dou Y.Wei Y.Zhang Z.Li C.Song C.Liu Y.Qi K.Li X.Li X.Qiao R.Wang K.Yang F.Han X.Transcriptome-wide analysis of RNA m 6A methylation regulation of muscle development in Queshan black pigs BMC Genomics 2420231163714299610.1186/s 12864-023-09346-w PMC 10161540 · doi ↗ · pubmed ↗
- 7Fei Q.Zou Z.Roundtree I.A.Sun H.L.He C.YTHDF 2 promotes mitotic entry and is regulated by cell cycle mediators P Lo S Biol.18202012210.1371/journal.pbio.3000664 PMC 717029432267835 · doi ↗ · pubmed ↗
- 8Feng S.Zhou H.Lin X.Zhu S.Chen H.Zhou H.Wang R.Wang P.Shao X.Wang J.Exercise promotes skeletal muscle growth in adolescents via modulating Mettl 3-mediated m 6A methylation of Myo D in muscle satellite cells Cell Mol. Biol. Lett.29202410.1186/s 11658-024-00670-x PMC 1161619239633280 · doi ↗ · pubmed ↗