The Benefits of Estradiol on Cognitive Aging in Rats Are Independent From Its Effects on Cardiometabolic Health
Christian Montanari, Emma L Dong, Shruti Srinivasan, Ana Paula De Oliveira Leite, Alyssa F Delarge, Matthieu J Maroteaux, Lucie D Desmoulins, Riva Menon, Alice B Walker, Sarah H Lindsey, Andrea Zsombok, Jill M Daniel

TL;DR
This study shows that estradiol improves cognitive aging in rats independently of its effects on heart and metabolic health.
Contribution
It demonstrates that estradiol's cognitive benefits are not dependent on its cardiometabolic effects.
Findings
Continuous estradiol treatment improved cognitive aging and cardiometabolic health.
Previous estradiol exposure improved cognition but not cardiometabolic health.
Delayed estradiol treatment improved health but not cognition.
Abstract
Research in preclinical models of menopause indicates that exogenously administered estrogens positively affect cognitive aging. However, clinical evidence indicates that the effects of estrogen therapy on cognition are inconsistent and may be modulated by preexisting cardiometabolic conditions. The extent to which cardiometabolic health affects the cognitive outcomes of estrogen therapy remains unclear. This study aimed to determine whether variations in cardiometabolic health, both prior to and resulting from different estradiol treatment regimens, are related to the ability of estradiol to improve the cognitive aging trajectory in ovariectomized Long-Evans rats. Cognitive function and health status were assessed at age 10 months after which rats were ovariectomized and administered vehicle or various estradiol treatments. Rats were assessed again at 18 (middle age) and 22 (old age)…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
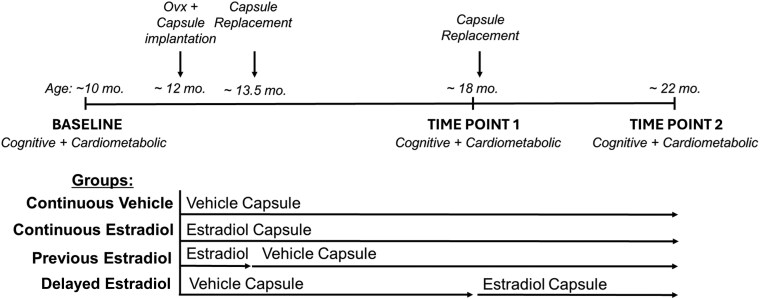 Figure 1
Figure 1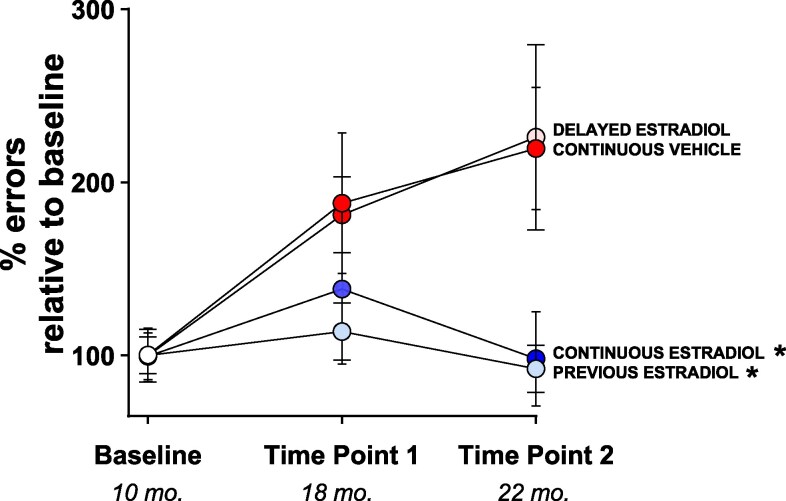 Figure 2
Figure 2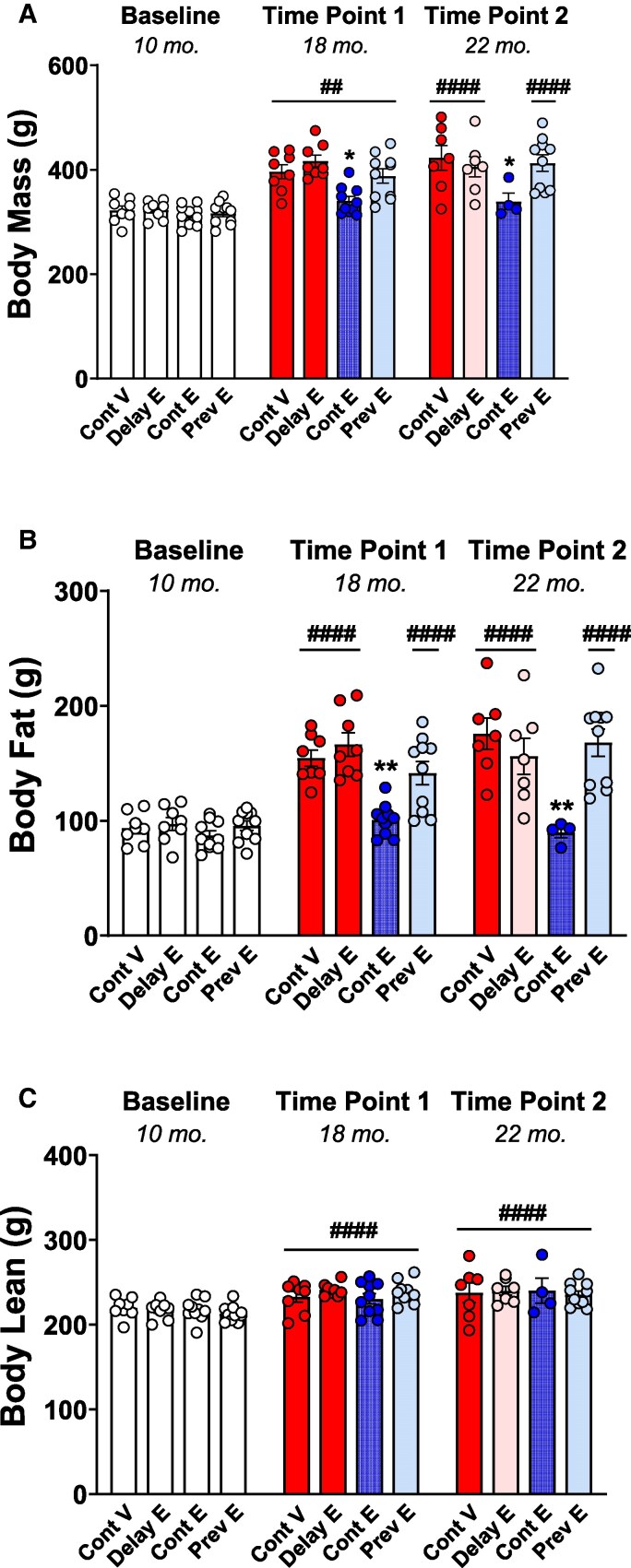 Figure 3
Figure 3 Figure 4
Figure 4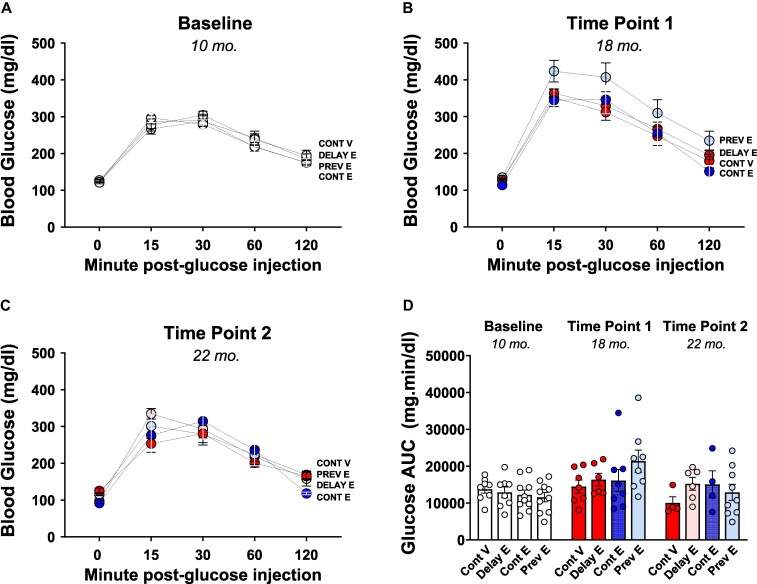 Figure 5
Figure 5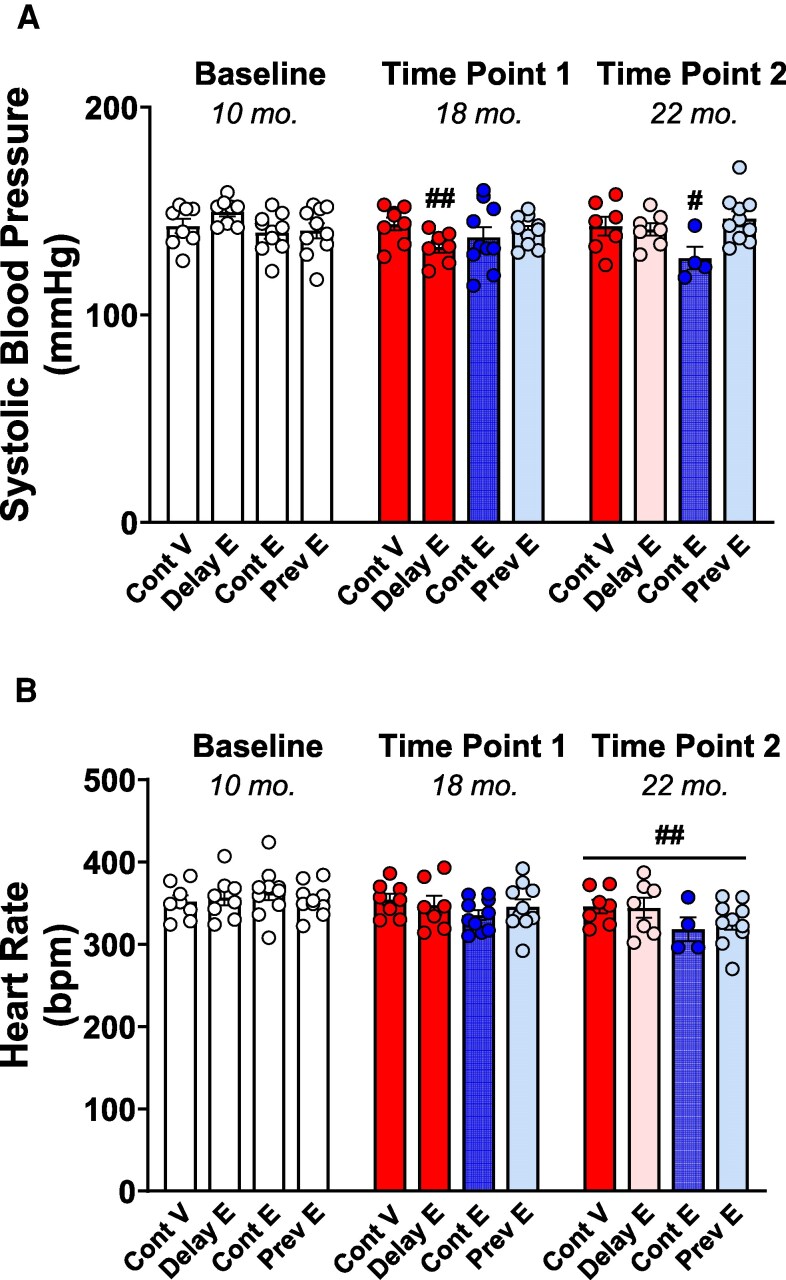 Figure 6
Figure 6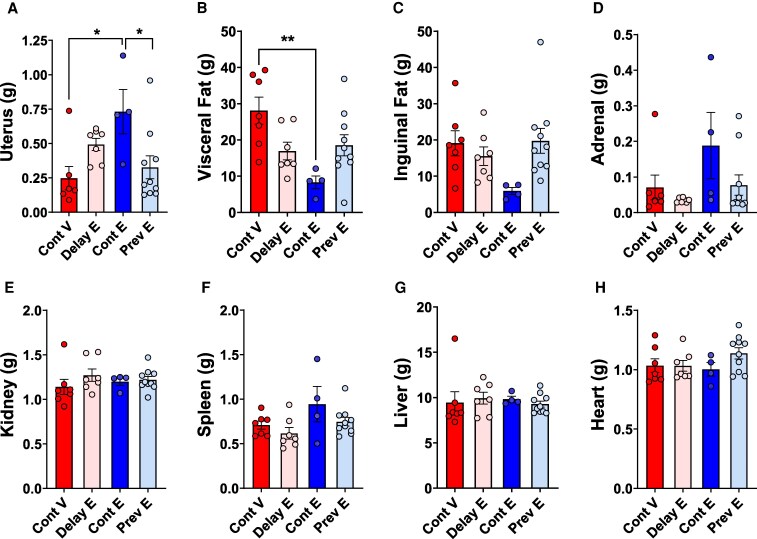 Figure 7
Figure 7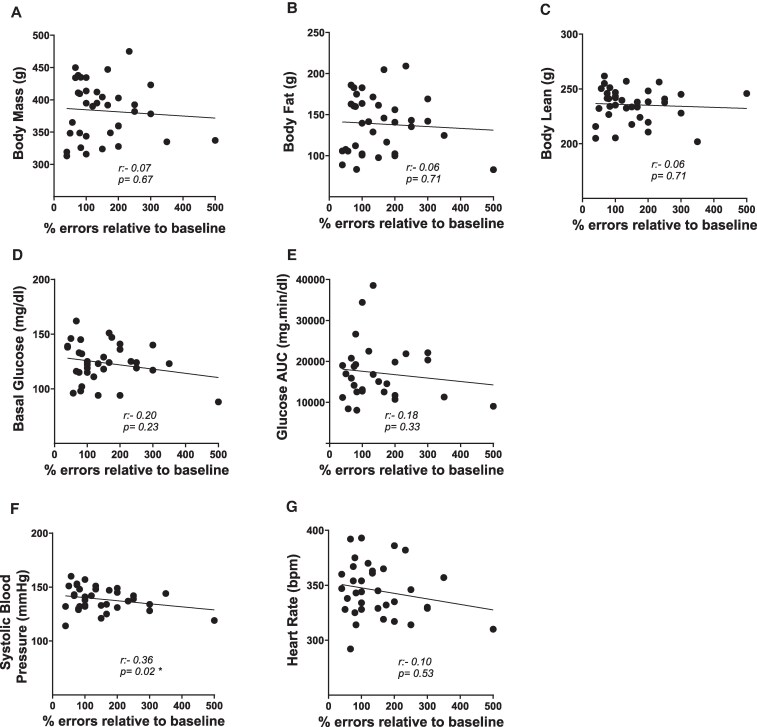 Figure 8
Figure 8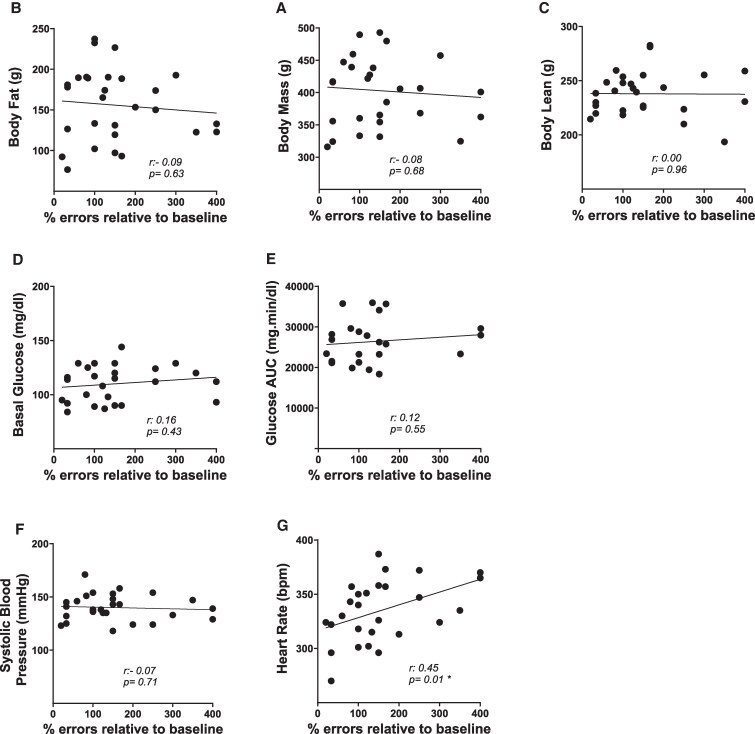 Figure 9
Figure 9- —NIH10.13039/100000002
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMenopause: Health Impacts and Treatments · Estrogen and related hormone effects · Cancer-related cognitive impairment studies
Decades of research in preclinical models of menopause have consistently demonstrated that estrogens exert neuroprotective effects on the brain and improve memory in aging female rodents (1). Research has demonstrated that estrogens enhance synaptic plasticity (2), promote cholinergic neurotransmission (3), stimulate adult neurogenesis (4), and improve mitochondrial bioenergetics in neurons (5). These effects are particularly noticeable in brain regions critical for cognitive function, such as the hippocampus and cerebral cortex (1).
In women, the loss of ovarian estrogens at menopause is hypothesized to be a risk factor for Alzheimer disease (6, 7). However, contrary to studies in animal models, clinical data have been inconsistent regarding the benefits of menopausal estrogen therapy on the brain and cognition, showing results that vary from beneficial to harmful (8, 9). Early observational studies and small clinical trials suggested that hormone therapy might help prevent age-related cognitive decline (for review, see (10)). Conversely, the large Women's Health Initiative (WHI) Memory Study (11-14) conducted by the National Institutes of Health (NIH), reported that hormone therapy may increase the risk of dementia. The Kronos Early Estrogen Prevention Study (KEEPS, (15)) and the KEEPS-Continuation Study (16) reported no harm or benefit from hormone therapy, as evaluated during the 4-year treatment period (15) and approximately 10 years after treatment termination (16).
Discrepancies in findings on the effects of estrogens on cognition between preclinical models and clinical studies, as well as among clinical studies themselves, might stem from differences in overall health status of participants. Indeed, preclinical research typically involves models of healthy aging, whereas clinical studies often include individuals with varying health conditions. Interestingly, preliminary evidence suggests that variations in clinical outcomes of hormone therapy might be partly explained by preexisting cardiometabolic conditions, such as type 2 diabetes and hypertension, which may blunt the effects of estrogens on cognition (17-19).
The extent to which cardiometabolic health affects the cognitive outcomes of estrogen therapy remains unclear. In our laboratory, we have successfully established a surgical model of menopause in rodents and consistently demonstrated that midlife 17β-estradiol treatment in recently ovariectomized rats leads to long-lasting memory enhancements that persisted well beyond estradiol exposure (20-25).
The present study aimed to determine whether variations in cardiometabolic health, prior to and/or following different estradiol treatment regimens, influence the ability of estradiol to positively affect cognitive aging. We conducted a 1-year longitudinal study assessing the cognitive aging trajectory and cardiometabolic health of rats from middle (10 months) to old (22 months) age. Our treatments aimed to model different populations of women, including those who start estrogen therapy at menopause and continue indefinitely, those who begin estrogen therapy at menopause but discontinue after a few years, those who start taking estrogens years after the onset of menopause, and those who never use menopausal estrogen therapy. We hypothesized that the effects of estrogens on cognition would be related to cardiometabolic health status. Understanding the nuanced relationship between cognitive and cardiometabolic health could lead to personalized treatment strategies that optimize cognitive health in aging women, ultimately reducing the risk of neurodegenerative diseases.
Materials and Methods
Subjects
Female Long-Evans rats (n = 40) were obtained from Envigo at age 70 days and allowed to age until the experiment began. The rats were pair-housed in a temperature-controlled vivarium on a 12-hour light/dark cycle (lights on at 7 Am), with unrestricted access to food (5V5R—PicoLab Select Rodent 50 IF/6F diet, PMI LabDiet; it contains a targeted level of isoflavones for estrogen-sensitive protocols) and water, unless otherwise specified. All experimental procedures were performed during the light cycle. Animal care adhered to the NIH’s Guide for the Care and Use of Laboratory Animals (26). All experimental procedures were approved by the Institutional Care and Use Committee of Tulane University. For experimental timeline, see Fig. 1.
Experimental timeline. Cognitive function and cardiometabolic health were evaluated at baseline (age 10 months). At age 12 months, rats were ovariectomized and implanted with either estradiol or vehicle capsules. The capsules were replaced at age 13.5 and 18 months, either maintaining the current vehicle or estradiol treatment or switching between them, resulting in the following treatment groups: continuous vehicle, continuous estradiol, previous estradiol, and delayed estradiol. To assess the effects of the different estradiol treatments over time, cognitive function and cardiometabolic health were reassessed at time point 1 (age 18 months, middle age) and at time point 2 (old age, 22 months). Ovx, ovariectomy.
Radial-Arm Maze Training and Baseline Cognitive Assessment
When rats reached age 9 months, training on the 8-arm radial maze spatial memory task began. During training, rats were food-restricted to maintain approximately 90% of their free-feeding weight. Rats were trained for 24 days (1 trial per day, 5 days per week). The maze (Coulbourn Instruments; 66 cm long, 9.5 cm wide, 11.5 cm high) had a metal grated floor and clear acrylic walls, with 8 arms extending radially from a central hub (28 cm diameter). The maze was placed on a table approximately 1 meter above the ground, centered in a room with multiple extra-maze cues. A single food reward (∼¼ of Froot Loops; Kellogg) was placed in an opaque dish (5.5 cm diameter, 1.25 cm tall) at the end of each arm, hidden from the maze center. For each trial, the rat was placed in the center of the maze, facing a pseudo-randomly chosen arm, and was allowed to explore the maze until all arms had been visited or 5 minutes elapsed. An arm entry was counted when all 4 paws crossed the arm midline, and reentries (errors) were scored if the rat entered a previously visited arm. The arm entry sequence was scored in real time by an observer located in a fixed position in the room. Baseline cognitive function for each animal was determined at the end of training and when rats were age 10 months by averaging the errors made during the first 8 arm choices over the final 4 days of radial-arm maze training.
Baseline Cardiometabolic Health Assessment
Following completion of radial-maze training, rats were returned to a free-feeding schedule. Approximately 2 weeks later and after body weights had stabilized at pretraining levels, baseline cardiometabolic assessment took place.
Cardiometabolic assessment included body composition analysis (dual-energy x-ray absorptiometry [DXA]), glucose tolerance testing, and blood pressure and heart rate measurements (tail-cuff plethysmography). The starting order of glucose tolerance and tail-cuff plethysmography were counterbalanced between subjects, with the body composition analysis being the final procedure.
Dual energy x-ray absorptiometry
Body composition was determined using DXA, calibrated with a reference phantom as described by the manufacturer (InAlyzer2, model S, Micro Photonics Inc). Rats were anesthetized by gas inhalation (isoflurane 2%) during the whole procedure. After the animals were placed inside the main unit, DXA scans were performed using the “rat” sample type with the “optimum” mode setting (scanning time: 84 seconds). The device uses dual-energy x-ray beams (80 kV/1.2 mA and 55 kV/0.7 mA) in a fan beam configuration. In vivo, the device achieves a precision of less than 5% coefficient of variation, with an SD of less than 1%. The scan area was 5.5 × 8.27 inches (∼14 × 21 cm), with an image pixel pitch of 99 × 99 μm. Body mass, body fat, and body lean were collected as weight (in g).
Glucose tolerance test
After a 4-hour fasting period in clean cages with free access to water, rats were injected intraperitoneally with a solution of 20% glucose (2 g/kg body weight). Blood samples were collected from the tail vein before the glucose injection (T0, to assess basal glucose) and at 15, 30, 60, and 120 minutes post glucose injection to track changes in glucose levels over a 2-hour period and calculate its area under the curve (AUC). Blood glucose levels were measured using a glucometer (OneTouch Verio Flex).
Tail-cuff plethysmography
Systolic blood pressure (measured in mm Hg) and heart rate (measured in beats per minute, bpm) were monitored using an automated tail-cuff volume-pressure recording system (Kent Scientific CODA system). Animals were acclimated to the clear plastic tube restraints for 2 days and measurements were obtained over 3 to 5 consecutive days to reduce the effect of restraint-induced stress. Ten to 15 consecutive measurements were taken for each animal while warming at 35 °C under slight restraint. Measures were averaged over consecutive days excluding values that were ±2 SDs from the mean.
Ovariectomy and Hormone Treatments
After completion of baseline cognitive and cardiometabolic assessments and when animals were approximately age 12 months, experimental treatments began. All animals were ovariectomized. Rats were anesthetized with intraperitoneal injections of ketamine (100 mg/kg; Bristol Laboratories) and xylazine (7 mg/kg; Miles Laboratories) and administered buprenorphine (0.375 mg/kg subcutaneously; Reckitt Benckiser Health Care) prior to surgery. Ovariectomy surgery involved bilateral flank incisions through the skin and muscle wall and removal of ovaries.
At the time of ovariectomy, rats were implanted with a subcutaneous 5-mm Silastic capsule (0.058-inch [∼1.5-mm] inner diameter and 0.077-inch [∼2-mm] outer diameter; Dow Corning) on the dorsal aspect of the neck. Capsules contained either cholesterol vehicle or 25% 17β-estradiol (Sigma-Aldrich) diluted in vehicle. Forty days (at age 13.5 months) and 6 months (at age 18 months) after ovariectomy and initial capsule implantation, rats were anesthetized by gas inhalation (isoflurane 2%) for capsule replacement. Capsule replacement involved either maintaining the current vehicle or estradiol treatment, or switching between them, resulting in the following randomly assigned estradiol treatment regimens (see Fig. 1): 1) continuous vehicle (n = 10; received vehicle for 10 months and modeling women who never use menopausal estrogen therapy); 2) continuous estradiol (n = 10; received estradiol for 10 months and modeling women who take and remain on estrogen therapy); 3) previous estradiol (n = 10; received estradiol for 40 days followed by vehicle for 8.5 months to model women who take menopausal estrogen therapy only for a few years in midlife); and 4) delayed estradiol (n = 10; received vehicle for 6 months followed by estradiol for 4 months to model women who begin taking estrogens years after menopause). We have previously demonstrated that capsules remain active for up to 5 months (20) and maintain blood serum estradiol levels at approximately 37 pg/mL in middle-aged female Long-Evans rats (27), which is within the physiological range.
Cognitive Aging Trajectory
After ovariectomies, rats underwent a weekly radial-arm maze testing trial, which was identical to the baseline trials, for the remainder of the experiment. During this time, they remained on free-feeding and underwent a 24-hour food deprivation period prior to each testing day. To determine the effect of the various hormone treatments on the cognitive aging trajectories of the animals, baseline performance that was determined before ovariectomy and hormone treatments was compared to performance at 2 later time points by averaging results (errors of first 8 arm choices) from 4 testing trials at each time point. Time point 1 assessment occurred when animals were approximately age 18 months, which was 4.5 months after termination of estradiol treatment in the previous estradiol group and before estradiol treatment began in the delayed estradiol group. Time point 2 testing occurred when animals were approximately age 22 months, which was 4 months following initiation of estradiol treatment in the delayed estradiol group.
The errors averaged at time point 1 and time point 2 were used to assess the cognitive aging trajectory of rats in each group, calculated as the percentage in the number of errors compared to baseline performance at 10 months. Using the mean centering method, the distribution of errors averaged at baseline first was centered around zero by subtracting the mean of the data from each individual data point; this preserved the variability in the data that reflected individual differences in memory. Then, to put baseline data on the same scale as subsequent time points (ie, percentage), 100 was added to each score.
Cardiometabolic Health Assessment Over Time
Cardiometabolic evaluations (including body composition analysis, glucose tolerance testing, blood pressure, and heart rate) were conducted at time point 1 (18 months) and time point 2 (22 months) approximately 2 weeks after completing spatial memory testing for the corresponding time point.
Hormone Treatment Verification
Vaginal smears were collected from each rat for 4 consecutive days before each of the 2 capsule replacements (at age ∼13.5 and 18 months, respectively) to confirm ovariectomy and hormone treatments. Smears from ovariectomized, cholesterol-treated rats were characterized by a predominance of leukocytes, while smears from ovariectomized, estradiol-treated rats were characterized by a predominance of cornified and nucleated epithelial cells, indicating that hormone treatments were effective. At the time of euthanasia, a 1-cm sample of the right uterine horn was collected from each rat and weighed to verify hormone treatment at the time of death.
Euthanasia and Tissue Collection
Rats were euthanized by decapitation under anesthesia induced by ketamine (100 mg/kg) and xylazine (7 mg/kg). Various organs and tissues (uterus, adrenal gland, kidney, spleen, liver, heart, inguinal fat, and visceral fat) were collected and weighed.
Statistical Analyses
Prior to group assignment and hormone treatment initiation, 4 rats were excluded from the experiment (2 rats for failing to learn the radial-arm maze training and 2 due to health issues). As a result, 36 rats were included at baseline and time point 1 (continuous vehicle: n = 8; delayed estradiol: n = 8; continuous estradiol: n = 10; previous estradiol: n = 10). Due to health issues, 4 rats were euthanized between time point 1 and time point 2 cognitive assessment and 4 additional rats between the 2 weeks separating the cognitive and cardiometabolic assessment at time point 2. Consequently, at time point 2, 32 rats were included in the cognitive assessment (continuous vehicle: n = 7; delayed estradiol: n = 8; continuous estradiol: n = 7; previous estradiol: n = 10) and 28 rats in the cardiometabolic and organ weight analyses (continuous vehicle: n = 7; delayed estradiol: n = 7; continuous estradiol: n = 4; previous estradiol: n = 10). Additional exclusions from the GTT were due to procedural issues. Final sample sizes for each procedure are provided in the figure legends.
All variables are expressed as mean ± SEM. Statistical analyses and graphs were generated using GraphPad Prism 10.2.3 software. Baseline cognitive performance at age 10 months, prior to the start of treatments, was assessed using a one-way analysis of variance (ANOVA). Cognitive aging trajectory from age 10 to 22 months was assessed using a mixed-effects model analysis with treatment (continuous vehicle, continuous estradiol, previous estradiol, delayed estradiol) as the between-subjects factor and time (baseline, time point 1, time point 2) as the within-subjects factor, followed by Dunnett post hoc tests in case of a statistically significant main or interaction effects (P ≤ .05).
Body composition parameters (body mass, body fat, body lean), basal glucose, glucose AUC, systolic blood pressure, and heart rate were assessed using a mixed-effects model analysis with treatment (continuous vehicle, continuous estradiol, previous estradiol, delayed estradiol) as the between-subjects factor and time (baseline, time point 1, time point 2) as the within-subjects factor, followed by Tukey post hoc tests in case of a statistically significant main or interaction effects (P ≤ .05).
At each testing (baseline, time point 1, time point 2), glucose levels following glucose injection were analyzed using 2-way ANOVA, with treatment (continuous vehicle, continuous estradiol, previous estradiol, delayed estradiol) as the between-subjects factor and minutes post glucose injection (0, 15, 30, 60, 120 minutes) as the within-subjects factor, followed by Tukey post hoc tests in case of a statistically significant main or interaction effects (P ≤ .05).
Both at time point 1 and time point 2, Pearson correlation coefficient (2-tailed) was performed to investigate the relationship between cognitive aging trajectory (measured as percentage errors in the radial arm maze relative to baseline) and cardiometabolic health measures. Correlation analyses were conducted on the total sample and independently within each treatment group, regardless of overall statistical significance.
The weights of organs collected at the time of death were analyzed using one-way ANOVA, followed by Tukey post hoc tests in case of a statistically significant main effect (P ≤ .05).
Results
Baseline Cognitive Performance
There was no statistically significant effect of group on memory performance at baseline (10 months), prior to the start of treatments (F(3, 32) = 2.152; P = .1130). Group mean errors ± SEM made during the first 8 arm choices were continuous vehicle: 0.81 ± 0.15; delayed estradiol: 0.72 ± 0.15; continuous estradiol: 1.12 ± 0.13; and previous estradiol: 0.90 ± 0.11.
Cognitive Aging Trajectory
As illustrated in Fig. 2, analysis of the percentage number of errors during radial-arm maze testing at time point 1 and time point 2 compared to baseline performance revealed significant main effects of treatment (F(3, 92) = 4.951; P = .0031) and time (F(2, 92) = 5.587; P = .0051) and no statistically significant interaction of treatment × time (F(6, 92) = 1.706; P = .1283). Post hoc comparisons indicated that the cognitive aging trajectory of the continuous vehicle group differed from both the continuous estradiol and previous estradiol groups (P ≤ .05 for both comparisons) but not from the delayed estradiol group (P = .94), and that performance at baseline across groups differed from performance both at time point 1 and time point 2 (P ≤ .05 for both comparisons).
*Cognitive aging trajectory. Mean (±SEM) percentage of errors during radial-arm maze testing at time point 1 (age 18 months) and time point 2 (age 22 months), compared to baseline performance at 10 months, for the continuous estradiol, continuous vehicle, previous estradiol, and delayed estradiol groups. At baseline, all groups are represented with white circles, as treatments had not yet begun. Following treatment initiation, groups were color-coded based on their respective estradiol treatment regimens: the continuous estradiol group is shown in blue circles both at time point 1 and time point 2 (indicating ongoing estradiol treatment at the time of testing); the continuous vehicle group is shown in red circles at both time point 1 and time point 2 (indicating ongoing vehicle treatment at the time of testing); the previous estradiol group is shown in light blue circles both at time point 1 and time point 2 (indicating estradiol treatment had been terminated months prior to testing); the delayed estradiol group is shown in red circles at time point 1 (indicating ongoing vehicle treatment at the time of testing), and in pink circles at time point 2 (indicating an ongoing delayed estradiol treatment initiated after testing at time point 1). Sample size at baseline: continuous vehicle, n = 8; delayed estradiol, n = 8; continuous estradiol, n = 10; previous estradiol, n = 10; sample size at time point 1: continuous vehicle, n = 8; delayed estradiol, n = 8; continuous estradiol, n = 10; previous estradiol, n = 10; sample size time point 2: continuous vehicle, n = 7; delayed estradiol, n = 8; continuous estradiol, n = 7; previous estradiol, n = 10. P less than or equal to .05 compared to continuous vehicle.
Body Composition Analysis
Body mass
As illustrated in Fig. 3A, analysis of body mass revealed a statistically significant main effect of treatment (F(3, 32) = 4.853; P = .0068) and time (F(2, 1.171, 32.7856) = 161.2; P < .0001) as well as a significant interaction of treatment × time (F(6, 56) = 8.478; P < .0001). Post hoc comparisons revealed no group differences at baseline, before treatments began. However, both at time point 1 and time point 2, the continuous estradiol group exhibited significantly lower body mass compared to all other groups (P ≤ .05 for all comparisons). When evaluating changes in body mass over time, the continuous vehicle and previous estradiol groups showed higher body mass both at time point 1 and time point 2 compared to baseline (P ≤ .0001 for all comparisons), with an additional increase from time point 1 to time point 2 (P < .05 for both groups). Results reveal a progressive weight gain in absence of estradiol (continuous vehicle) or when estradiol treatment was interrupted months before testing (previous estradiol). In the delayed estradiol group, body mass was significantly higher both at time point 1 and time point 2 compared to baseline (P ≤ .0001 for both comparisons), but no difference was observed between time point 1 and time point 2 (P = .4383), indicating stabilization of body mass as estradiol treatment started after testing at time point 1. In contrast, the continuous estradiol group showed an increased body mass compared to baseline only at time point 1 (P ≤ .01), suggesting that continuous ongoing estradiol treatment blunted naturally occurring weight gain following the loss of ovarian function.
*Body composition. Mean (±SEM) A, body mass; B, body fat; and C, body lean mass measured at baseline (10 months), time point 1 (18 months), and time point 2 (22 months) in the continuous vehicle (Cont V), delayed estradiol (Delay E), continuous estradiol (Cont E), and previous estradiol (Prev E) groups. At baseline, all groups are depicted in white, as treatments had not yet begun. At time points 1 and 2, group colors indicate treatment conditions: continuous vehicle (red), continuous estradiol (blue), and previous estradiol (light blue); the delayed estradiol group is depicted in red at time point 1, reflecting ongoing vehicle treatment at the time of testing, and in pink at time point 2, indicating the initiation of delayed estradiol treatment after testing at time point 1. Sample size at baseline: Cont V, n = 8; Delay E, n = 8; Cont E, n = 10; Prev E, n = 10; sample size at time point 1: Cont V, n = 8; Delay E, n = 8; Cont E, n = 10; Prev E, n = 10; sample size at time point 2: Cont V, n = 7; Delay E, n = 7; Cont E, n = 4; Prev E, n = 10. *P less than or equal to .05, and *P less than or equal to .01 compared to all the other groups; ##P less than or equal to .01, and ####P less than or equal to .0001 compared to the corresponding baseline within each group.
Body fat mass
As illustrated in Fig. 3B, analysis of body fat mass revealed a statistically significant main effect of treatment (F(3, 32) = 9.924; P < .0001), time (F(2, 56) = 125.1; P < .0001), and a significant time × treatment interaction (F(6, 56) = 12.32; P < .0001). Consistent with findings in body mass, no differences were identified among groups at baseline (and before treatments began), whereas both at time point 1 and time point 2 the continuous estradiol group had lower body fat compared to the other treatment groups (P ≤ .01 for all comparisons). Additionally, the continuous estradiol group did not show changes in body fat over time (baseline vs time point 1; P = .1106; baseline vs time point 2, P = .9789; time point 1 vs time point 2, P = .4406). In the continuous vehicle and previous estradiol groups, body fat levels were elevated both at time point 1 and time point 2 compared to baseline (P ≤ .0001 for all comparisons), with a further increase noted from time point 1 to time point 2 (P ≤ .05). For the delayed estradiol group, body fat was significantly higher both at time point 1 and time point 2 compared to baseline (P ≤ .0001 for all comparisons), with no difference between time point 1 and time point 2 (P = .2829).
Body lean mass
As illustrated in Fig. 3C, analysis of body lean mass indicated a statistically significant main effect of time (F(2, 56) = 62.80; P < .0001), but no significant effect of treatment (F(3, 32) = 0.1343; P = .9389) or treatment × time interaction (F(6, 56) = 1.679; P = .1432). Post hoc comparisons indicated that body lean was significantly higher both at time point 1 and time point 2 compared to baseline (P < .0001 for both comparisons).
Basal Glucose Levels
As illustrated in Fig. 4, analysis of basal glucose levels prior to glucose injection revealed a statistically significant main effect of treatment (F(3, 32) = 7.441; P = .0006) and time (F(1.672, 44.30) = 26.35; P < .0001) as well as a significant time × treatment interaction (F(6, 53) = 5.376; P = .0002). No group differences were detected at baseline. At time point 1, basal glucose in the continuous estradiol group was lower than the previous estradiol group (P = .0201). At time point 2, basal glucose in the continuous estradiol group was lower than both the previous estradiol (P = .0007) and continuous vehicle (P = .0012) groups, while basal glucose in the delayed estradiol group was lower than the continuous vehicle group (P = .0275), indicating that ongoing estradiol treatment, even if initiated long past loss of ovarian function, lowered basal glucose levels. All groups except the continuous vehicle group showed lower basal glucose levels at time point 2 compared to baseline (P ≤ .05 for all comparisons). A statistically significant decrease between time point 1 and time point 2 was detected in the delayed estradiol and previous estradiol groups (P ≤ .01 for both comparisons).
*Basal glucose levels. Mean (±SEM) basal glucose levels at baseline (10 months), time point 1 (18 months), and time point 2 (22 months) in the continuous vehicle (Cont V), delayed estradiol (Delay E), continuous estradiol (Cont E), and previous estradiol (Prev E) groups. At baseline, all groups are depicted in white, as treatments had not yet begun. At time points 1 and 2, group colors indicate treatment conditions: continuous vehicle (red), continuous estradiol (blue), and previous estradiol (light blue); the delayed estradiol group is depicted in red at time point 1, reflecting ongoing vehicle treatment at the time of testing, and in pink at time point 2, indicating the initiation of delayed estradiol treatment after testing at time point 1. Sample size at baseline: Cont V, n = 8; Delay E, n = 8; Cont E, n = 10; Prev E, n = 10; sample size at time point 1: Cont V, n = 8; Delay E, n = 8; Cont E, n = 10; Prev E, n = 9; sample size at time point 2: Cont V, n = 7; Delay E, n = 6; Cont E, n = 4; Prev E, n = 9. *P less than or equal to .05, **P less than or equal to .01, and **P less than or equal to .001 indicate group differences; #P less than or equal to .05 compared to the corresponding baseline within each group.
Glucose Tolerance Test
As illustrated in Fig. 5A to 5C, at each testing (baseline, time point 1, time point 2), there was a statistically significant main effect of minutes post glucose injection (baseline: F(3.175, 101.6) = 199.9; P < .0001; time point 1: F(2.315, 57.88) = 134.9; P < .0001; time point 2: F(2.162, 41.08) = 75.08; P < .0001). Post hoc comparisons revealed that at each test, nearly all pairwise comparisons were statistically significant (P ≤ .05 for all comparisons). Glucose levels at 15 and 30 minutes post glucose injection were significantly higher compared to basal preinjection levels (0 minutes post glucose injection; P ≤ .01 for all comparisons), while comparisons between 30- and 120-minute post glucose injections indicated a decrease of glucose levels (P ≤ .05 for all comparisons). This suggests a peak in glucose levels 30 minutes after the glucose injection, followed by a gradual decline.
Glucose tolerance test. Mean (±SEM) A to C, glucose levels over the 2-hour period following glucose injection measured at A, baseline; B, time point 1; and C, time point 2 in the continuous vehicle (Cont V), delayed estradiol (Delay E), continuous estradiol (Cont E), and previous estradiol (Prev E) groups. The corresponding mean (±SEM) area under the curve (AUC) values are shown in panel D. Analysis of glucose AUC revealed a statistically significant main effect of time point (F(1.458, 55.40) = 5.535; P < .05), with post hoc tests indicating higher AUC at time point 1 compared to baseline (P = .0066). At baseline, all groups are depicted in white, as treatments had not yet begun. At time points 1 and 2, group colors indicate treatment conditions: continuous vehicle (red), continuous estradiol (blue), and previous estradiol (light blue); the delayed estradiol group is depicted in red at time point 1, reflecting ongoing vehicle treatment at the time of testing, and in pink at time point 2, indicating the initiation of delayed estradiol treatment after testing at time point 1. Sample size at baseline: Cont V, n = 8; Delay E, n = 8; Cont E, n = 10; Prev E, n = 10; sample size at time point 1: Cont V, n = 7; Delay E, n = 6; Cont E, n = 8; Prev E, n = 8; sample size at time point 2: Cont V, n = 4; Delay E, n = 6; Cont E, n = 4; Prev E, n = 9.
No statistically significant effect of treatment was found: baseline: F(3, 32) = 0.1964; P = .8981; time point 1: F(3, 25) = 1.986; P = .1419; time point 2: F(3, 19) = 0.1531; P = .9264). There was no interaction of minutes post glucose injection × treatment: baseline: F(12, 128) = 1.122; P = .3478; time point 1: F(12, 100) = 0.9442, P = .5068; time point 2: F(12, 76) = 1.400; P = .1848).
Analysis of the glucose AUC (Fig. 5D) revealed a statistically significant main effect of time point (F(1.458, 55.40) = 5.535; P < .05). Post hoc comparisons indicated a higher glucose AUC at time point 1 compared to baseline (P = .0066). No significant main effect of treatment (F(3, 76) = 0.7635) or a significant time × treatment interaction (F(6, 76) = 1.392) was revealed.
Systolic Blood Pressure
As illustrated in Fig. 6A, analysis of systolic blood pressure indicated no statistically significant main effect of treatment (F(3, 32) = 1.672; P = .1927) or time (F(1.655, 44.68) = 3.113; P = .06331) but a significant treatment × time interaction (F(6, 54) = 3.085; P = .0114). Post hoc comparisons showed a significant decrease in systolic blood pressure from baseline to time point 2 in the continuous estradiol group (P = .0588). We also found a lower systolic blood pressure in the delayed estradiol group at time point 1 (prior to estradiol treatment initiation) compared to its baseline (P = .0039).
Systolic blood pressure and heart rate. Mean (±SEM) A, systolic blood pressure and B, heart rate measured at baseline (10 months), time point 1 (18 months), and time point 2 (22 months) in the continuous vehicle (Cont V), delayed estradiol (Delay E), continuous estradiol (Cont E), and previous estradiol (Prev E) groups. At baseline, all groups are depicted in white, as treatments had not yet begun. At time points 1 and 2, group colors indicate treatment conditions: continuous vehicle (red), continuous estradiol (blue), and previous estradiol (light blue); the delayed estradiol group is depicted in red at time point 1, reflecting ongoing vehicle treatment at the time of testing, and in pink at time point 2, indicating the initiation of delayed estradiol treatment after testing at time point 1. Sample size at baseline: Cont V, n = 8; Delay E, n = 8; Cont E, n = 10; Prev E, n = 10; sample size at time point 1: Cont V, n = 8; Delay E, n = 8; Cont E, n = 10; Prev E, n = 10; sample size at time point 2: Cont V, n = 7; Delay E, n = 7; Cont E, n = 4; Prev E, n = 10. #P less than or equal to .05, and ##P less than or equal to .01 compared to the corresponding baseline within each group. bpm, beats per minute.
Heart Rate
As illustrated in Fig. 6B, analysis of heart rate indicated a statistically significant main effect of time (F(1.890, 51.97) = 8.143; P = .0010), but no significant effect of treatment (F(3, 32) = .8723; P = .4656) or treatment × time interaction (F(6, 55) = 1.595; P = .1662). Post hoc comparisons indicated that heart rate was significantly lower at time point 2 compared to baseline (P = .0026), with the comparison between baseline and time point 1 approaching statistical significance (P = .0612).
Organ Weights
As illustrated in Fig. 7, there were statistically significant effects of treatment for uterus (F(3, 24) = 4.326; P = .0142) and visceral fat (F(3, 24) = 5.199; P = .0066). Post hoc comparisons indicated that the uterus weight of the continuous estradiol group was significantly higher than the previous estradiol (P = .0357) and the continuous vehicle (P = .0152) groups. For visceral fat, post hoc comparisons showed that the continuous estradiol group had significantly lower visceral fat compared to the continuous vehicle group (P = .0044).
*Organ weights. Mean (±SEM) weights of A, the uterus; B, visceral fat; C, inguinal fat; D, adrenal; E, kidney; F, spleen; G, liver; and H, heart collected at age 22 months for the continuous vehicle (Cont V), delayed estradiol (Delay E), continuous estradiol (Cont E), and previous estradiol (Prev E) groups. Sample size: Cont V, n = 7; Delay E, n = 7; Cont E, n = 4; Prev E, n = 10. *P less than or equal to .05 compared to Cont V and Prev E; *P less than or equal to .01 compared to Cont V.
There were no statistically significant treatment effects for other organs that were collected (inguinal fat: F(3, 24) = 2.728; P = .0746; adrenal: F(3, 24) = 2.159, P = .1191; kidney: F(3, 24) = 0.7455; P = .5355; spleen: F(3, 24) = 2.340; P = .0987; liver: F(3, 24) = .1889; P = .9029; heart: F(3, 24) = 1.411; P = .2640).
Correlation Between Cognitive Aging Trajectory and Cardiometabolic Health Measures
As shown in Fig. 8, at time point 1 (18 months), the only statistically significant correlation observed in the total sample was between cognitive aging trajectory (measured as percentage number of errors at time point 1 compared to baseline) and blood pressure (r = −0.3681; P = .0296). However, this correlation was independent of the estradiol treatments, as it was found only in the continuous vehicle group (continuous vehicle: r = −0.7114; P = .0478; delayed estradiol: r = −0.5954; P = .1584; continuous estradiol: r = −0.2756; P = .4409; previous estradiol: r = −0.3244; P = .3605).
*Correlation between cognitive aging trajectory and measures of cardiometabolic health in the total sample at time point 1 testing (18 months). Pearson correlation coefficients (r) and corresponding P values for the relationship between cognitive aging trajectory and A, body mass; B, body fat; C, body lean; D, basal glucose; E, glucose AUC; F, systolic blood pressure; and G, heart rate. Sample size: n = 36, except for the correlations involving basal glucose (n = 35) and glucose AUC (n = 29), due to additional exclusions of rats for procedural issues. P less than or equal to .05. AUC, area under the curve; bpm, beats per minute.
As shown in Fig. 9, at time point 2 (22 months), we found only an overall statistically significant correlation between cognitive aging trajectory and heart rate (r = 0.4514; P = .0159), with no significant correlation within groups (continuous vehicle: r = 0.0791; P = .8661; continuous estradiol: r = 0.3716; P = .6254; delayed estradiol: r = 0.4816; P = .2739; previous estradiol: r = 0.4100; P = .2393).
*Correlation between cognitive aging trajectory and measures of cardiometabolic health in the total sample at time point 2 testing (22 months). Pearson correlation coefficients (r) and corresponding P values for the relationship between cognitive aging trajectory and A, body mass; B, body fat; C, body lean; D, basal glucose; E, glucose AUC; F, systolic blood pressure; and G, heart rate. Sample size: n = 28, except for the correlations involving basal glucose (n = 26) and glucose AUC (n = 23), due to additional exclusions of rats for procedural issues. P less than or equal to .05. AUC, area under the curve; bpm, beats per minute.
No additional significant correlations or trends were observed between cognitive aging trajectory and the other cardiometabolic measures, including when analyzed across the total sample and within the 4 different treatment groups.
Discussion
The present 1-year longitudinal study followed aging ovariectomized female rats that were administered various regimens of estradiol treatments and assessed their cognitive aging trajectories and cardiometabolic health status from middle to old age (age 10-22 months). The goal of the work was to determine if there is a relationship between the ability of estrogens to affect cognitive aging and their ability to affect cardiometabolic aging in a rodent model of menopause. Results revealed that a regimen of continuous, ongoing estradiol treatment positively affected both cognition (ie, enhanced performance on spatial memory radial-arm maze task) and cardiometabolic health (ie, reduction of body mass, body fat and visceral fat, basal glucose, and systolic blood pressure). In contrast, a previous history of 40 days of estradiol treatment that was terminated months prior to assessments had positive lasting effects on cognition, but no positive lasting effects on measures of cardiometabolic health. Finally, a delayed estradiol treatment that was initiated 6 months after ovariectomy had no effect on cognition but had positive effects on some measures of cardiometabolic health (ie, reduction of basal glucose). Overall, our results indicate that estrogen’s actions on systems mediating cognitive function and those mediating cardiometabolic health are likely through independent mechanisms and that the effects of estradiol on cognitive aging are not secondary to their effects on cardiometabolic systems.
Effects of Estrogens on Cognition
The results of the present study indicate that, when initiated at the time of ovariectomy and subsequent ovarian function loss, both ongoing, continuous estradiol treatment (continuous estradiol) and a previous short-term estradiol treatment (previous estradiol), positively influenced the cognitive aging trajectory. Noteworthy, a previous 40-day estradiol treatment provided long-term cognitive benefits until the end of the experiment, corresponding to 10.5 months after estradiol treatment cessation. This effect was comparable to that observed in animals that were exposed to estradiol for the entire year of the experiment. On the other hand, exposing animals to estradiol 5 months after ovarian function loss did not benefit the cognitive aging trajectory, in spite of the duration of the delayed estradiol treatment of 4 months. These findings corroborate our previous results, showing that continuous estradiol treatment, as well as a prior estradiol exposure, significantly enhanced long-term memory (20-25, 27). However, unlike previous studies in which comparisons were made only between groups, in the present study the assessment of the effects of estradiol on cognition was conducted over a year-long longitudinal study to track within each group the cognitive aging trajectory from middle to old age (age 10-22 months) and comparing cognitive performance to baseline levels prior to ovariectomy. Collectively, our results indicate that 1) estradiol treatment must be initiated within a critical window period after loss of ovarian function to confer cognitive benefits, and 2) a short 40-day estradiol regimen initiated at the time of ovariectomy offers long-term protection against cognitive decline, comparable to a continuous estradiol treatment.
Clinical data on the cognitive benefits of menopausal estrogen therapy have been inconsistent. Early observational and clinical studies ((28-32); see 10 for a review) suggest that mid-life estrogen therapy following natural or surgical menopause provides long-term cognitive benefits. In contrast, the WHI Memory Study, a large randomized, double-blind, placebo-controlled clinical trial conducted by the NIH, found no cognitive benefit from hormone therapy. The study reported that hormone therapy regimens involving chronic conjugated equine estrogens (CEEs) (13, 14) or chronic CEE plus medroxyprogesterone (11, 12) did not protect against age-related cognitive decline and may even increase the risk of dementia. However, unlike other studies, the WHI Memory Study administered hormone treatment to women aged 65 years and older (mean age, 73 years), which is, on average, more than 2 decades after ovarian hormone levels decline during menopause. Nonetheless, recent results from the KEEPS (15) and the KEEPS-Continuation investigations (16) found neither harm nor benefit from hormone therapy in recently postmenopausal women, keeping the debate still open. The KEEPS trial was a 4-year, randomized, double-blind, placebo-controlled trial designed to assess the effects of oral CEE or transdermal 17β-estradiol, both with progesterone, in healthy, recently postmenopausal women (mean age, 53 years, with an average of 1.4 years since menopause). The trial demonstrated no significant cognitive benefit or harm during the 4 years of menopausal hormone therapy (15). Moreover, to investigate the long-term effects of the previously administered treatments, the observational KEEPS-Continuation Study (16) evaluated cognitive outcomes approximately 10 years after completion of oral CEE plus progesterone or transdermal 17β-estradiol plus progesterone. Results revealed that women receiving either estrogen formulation exhibited similar cognitive performance to those assigned to placebo.
The critical period hypothesis has been proposed as a possible explanation for the discrepancies across studies, suggesting that the cognitive benefits of estrogens may be apparent only if treatment is initiated near the time of menopause (33, 34); current clinical recommendations emphasize that hormone therapy provides the most benefit for patients when initiated within 10 years of menopause (North American Menopause Society, 2017 (35)).
Effects of Estrogens on Body Composition
Our results indicate that ongoing, continuous estradiol exposure led to lower body mass and body fat compared to the other treatment groups. Moreover, unlike the other groups, body mass and body fat in the continuous estradiol group did not increase over time. In contrast to its lasting enhancement on cognitive aging, a previous history of 40-day estradiol exposure did not exert a lasting effect on body composition parameters. Indeed, the previous estradiol group showed similar body mass and body fat compared to the continuous vehicle and delayed estradiol groups, with both parameters increasing over time compared to baseline. Similarly, when estradiol treatment in the delayed estradiol group was initiated 5 months after ovariectomy, once significant body mass and fat augmentation had occurred, it was not able to suppress ovariectomy-induced body mass and fat increase, even with 4 months of treatment. Interestingly, the different estradiol treatment regimens did not affect body lean mass, which increased over time without significant group differences.
These findings are in line with our previous observations, in which ongoing estradiol treatment counteracted an increase in body mass following ovariectomy only if treatment was initiated at the time of ovarian function loss (3). In the present study we also evaluated changes in body fat and lean mass over a year-long period and weighed different organs at the time of death, highlighting that the ovariectomy-induced increase in body weight is primarily due to changes in the fat content. Moreover, continuous exposure to estradiol led to a reduction in visceral fat content by the end of the experiment.
Our findings align with human studies showing that changes in the hormonal milieu at menopause are associated with increased body mass and fat, which can be attenuated by estrogen administration (36). Moreover, studies have demonstrated a strong link between visceral fat and metabolic and cardiovascular disorders, with an increase in visceral adiposity identified as a risk factor for insulin resistance, type 2 diabetes, and cardiovascular disease mortality (37, 38).
Our results indicate that estradiol regimens initiated at a healthy weight just after ovariectomy (continuous estradiol and previous estradiol groups) led to improved cognitive aging even under conditions of subsequent body weight and fat increase (previous estradiol). However, when estradiol was initiated months after ovariectomy and after body weight and fat increase occurred (delayed estradiol), no improvements in cognitive progression were observed. On one hand, these results support the critical period hypothesis, suggesting that the timing of estradiol initiation is crucial for its cognitive benefits. On the other hand, they also indicate that health status at the time of hormone treatment initiation may determine the efficacy of estradiol. These two possibilities are not mutually exclusive, and both the timing of hormone treatment initiation and health status at the time of initiation could synergistically contribute to the effects of estrogens on cognition.
Effects of Estrogens on Glucose Levels
In our study, ongoing estradiol treatment, initiated either at ovariectomy or months later, was associated with reduced basal blood glucose levels. Specifically, at age 18 months (time point 1), the continuous estradiol group exhibited significantly lower basal glucose levels compared to the previous estradiol group, which had terminated treatment months prior. At 22 months (time point 2), the continuous estradiol group had lower basal glucose levels than both the previous estradiol and continuous vehicle groups, but not compared to the delayed estradiol group, which had significantly lower levels than the continuous vehicle group. These results indicate that ongoing estradiol treatment effectively reduces basal glucose, regardless of when treatment begins after ovarian function loss.
Congruent with these findings, previous preclinical studies support the critical role of estrogens in regulating glucose homeostasis and other metabolic parameters. Metabolic dysregulation has been observed in ovariectomized rats (39) and mice (40, 41) on a standard diet. In these models, estradiol treatment has been shown to reduce both fasting and fed glucose levels, along with improvements in glucose tolerance (41). High-fat diet exposure is commonly used to induce obesity in rodent models, with female rats demonstrating greater protection against metabolic diseases; however, this sex-specific metabolic advantage diminished following ovariectomy (42).
Evidence from clinical trials indicates that estrogens treatment in nondiabetic postmenopausal women, started either in the early postmenopausal period (43-46) or within 10 years after menopause (47), is linked to reductions in fasting glucose and a lower risk of developing diabetes.
Although estradiol treatment resulted in significantly lower basal glucose levels, no group differences were observed in glucose levels during the GTT. This divergence may reflect distinct regulatory mechanisms at play in basal vs glucose-stimulated conditions. Estradiol has been shown to enhance glucose uptake by upregulating and facilitating the translocation of glucose transporter 4 (GLUT4) to the cell surface (48), a process critical in maintaining basal glucose homeostasis. In contrast, other regulatory pathways may predominate during acute glucose challenges. For example, glucokinase, a central component of the glucostat system responsible for glucose homeostasis (49), plays a key role in liver-mediated glucose regulation by acting as a glucose sensor and initiating glycolysis in response to changes in blood glucose levels (50). This pathway integrates signals from various organs, such as the liver, pancreas, and brain, to maintain glucose homeostasis during feeding or glucose challenges (51).
Effects of Estrogens on Cardiovascular Systems
Our study showed that continuous estradiol treatment decreased systolic blood pressure at age 22 months (time point 2), compared with baseline (10 months). Studies conducted in rodent models indicate that experimentally induced ovarian function loss leads to decreased estradiol levels and an elevated risk of hypertension and cardiovascular diseases (for a review, see (52)). Consistent with our findings in the continuous estradiol group, studies have shown that continuous estradiol treatment for 6 months (53) or 80 days (54) during a critical period post ovariectomy, reduces systolic blood pressure in Long-Evans rats of similar age to those in our study. Interestingly, when estradiol was administered months after ovariectomy in the delayed estradiol group, the treatment had no effect on blood pressure. These results support the hypothesis that estradiol needs to be administered within a critical window after the loss of ovarian function to positively affect blood pressure in aging females and are consistent with previous findings in rodent models (55, 56). Unexpectedly, we found that the delayed estradiol group had lower systolic blood pressure at 18 months (time point 1) compared to baseline measures. This measurement was taken before the initiation of estradiol treatment in this group, at a time when its treatment was identical to the continuous vehicle group. This unexpected finding is likely attributable to an elevated systolic blood pressure at baseline in the delayed estradiol group, but not in the others, relative to the typical range for intact Long-Evans female rats of similar age (53, 54).
Results revealed a reduced heart rate at 22 months (time point 2) compared to baseline (prior loss of ovarian function) across treatment groups with no effect of estradiol treatment at any time point. Data regarding estradiol and heart rate are complex and often contradictory. For example, ovariectomy in rats did not affect heart rate measured months after loss of ovarian function (57). However, consistent with the present results is a report in which there was a reduction in heart rate over time in ovariectomized rats, with no effect of estradiol (58). In humans, menopause was associated with decreased heart rate variability that was improved by estrogen therapy (59). In contrast was a report in which hormone therapy did not significantly alter resting heart rate (60).
Menopause in humans leads to a decline in endogenous estrogens and an increased risk of cardiovascular diseases (52). Observational trials suggest that women receiving hormone therapy early in menopause have a reduced risk of cardiovascular mortality later in life (61-63). Contrary to this evidence, the WHI Memory Study (64, 65) found no cardiovascular benefits from estrogen therapy. Instead, it indicated that both chronic CEE and CEE plus medroxyprogesterone increased cardiovascular risk, irrespective of the timing of treatment initiation relative to menopause onset. However, a nonsignificant trend was observed, suggesting women who initiated hormone therapy closer to menopause might have a reduced risk compared with those more distant from menopause.
Relationship Between Effects of Estrogens on Cognition and Its Effects on Cardiometabolic Systems
Numerous studies have indicated that cognitive decline is more rapid in individuals with diabetes, who also face a higher risk of Alzheimer and related dementias than their nondiabetic peers (66). Interestingly, some studies suggested that certain antidiabetic medications, like metformin, reduce the risk of neurodegenerative diseases (67). In the female population, obese women with type 2 diabetes have twice the dementia risk of women at a normal weight (68). In 2 WHIMS follow-up studies, estrogen effects were evaluated within the context of type 2 diabetes. One study found that estrogen alone—but not in combination with progestin—amplified diabetes-related risks for dementia and cognitive decline (18). Another study found that hormone therapy was linked to reduced brain volume in women with type 2 diabetes compared to those with type 2 diabetes who received a placebo, while brain volume in nondiabetic women was unaffected (69).
Numerous studies also have demonstrated a direct association between cardiovascular complications and accelerated cognitive decline, contributing to Alzheimer disease and vascular dementia (70). The SPRINT MIND trial highlighted that while intensive blood pressure–lowering (<120 mm Hg) compared to standard treatment (<140 mm Hg) does not prevent dementia, it does reduce the occurrence of mild cognitive impairment (71). Contrasting findings, such as those from the Hypertension in the Very Elderly Trial (HYVET), show that a year of antihypertensive treatment in adults older than 80 years did not significantly lower dementia risk (72). However, age at intervention could explain these differences, as elevated systolic blood pressure after age 60 was counterintuitively found to be associated with improved cognitive performance (73). Regarding women in menopause, some findings indicated that initiating hormone therapy in the initial menopausal years reduces the risk of dementia and cardiovascular mortality years later (63).
Based on this evidence, our expectations for the present study were that cardiometabolic health would modulate the effects of estradiol on cognitive aging trajectory. Surprisingly, our results suggest that the benefits of estradiol on cognition are independent from its effects on cardiometabolic systems. Indeed, we found that both a regimen of continuous ongoing estradiol treatment and a prior short-term period of midlife estradiol treatment improved the cognitive aging trajectory from middle to old age. However, little to no relationship between the cognitive benefits of estradiol and its effects on cardiometabolic health was identified. For example, continuous estradiol treatment positively affected both cognition and cardiometabolic health. In contrast, a previous history of 40 days of estradiol treatment terminated months prior testing had positive lasting effects on cognition, but did not confer any benefit to cardiometabolic health. Additionally, delayed estradiol treatment that was initiated months after ovariectomy showed no effect on cognition but had positive effects on some measures of cardiometabolic health.
Our correlation analyses support the lack of association between the effect of estradiol on cognitive aging and its effects on cardiometabolic health. At time point 1 (18 months), we found a significant negative correlation between systolic blood pressure and our measure of cognitive aging trajectory, indicating that as systolic blood pressure increases, cognitive performance improves (ie, a lower percentage of errors). At time point 2 (22 months), a significant positive correlation was observed between heart rate and our measure of cognitive aging trajectory, suggesting that as heart rate increases, cognitive performance worsens (ie, a higher percentage of errors). In spite of these overall correlations, we found no relationship of these measures of cardiometabolic health with cognition in any of the estradiol treatment groups. Interestingly, our findings align with recent human studies showing that elevated systolic blood pressure after age 60 is associated with a reduced risk of dementia (69), while an increase in heart rate tends to be linked to a decrease in cognitive performance (74).
Considerations and Future Directions
Although the continuous vehicle group showed increased body mass and fat over time as compared to baseline, other measures of cardiometabolic health did not show age-related changes in this group. In the present study, rats were assessed from middle (10 months) to old age (22 months). It may be that age-related decrements in cardiometabolic health may have already been present at our baseline assessment taken in middle age. But importantly, group differences in body mass and fat and basal glucose levels at time points 1 and 2 support group differences in cardiometabolic health over time. Additionally, our study focused on healthy aging and all animals were fed healthy diets. Future studies will assess the effects of estrogens and aging in models of unhealthy aging in which preexisting cardiometabolic disease is present at the time of loss of ovarian function.
A prolonged exposure to estradiol was associated with a lower long-term survival rate in the continuous estradiol group following testing at time point 1, after 6 months of estradiol administration. Despite this, the results from the cognitive and cardiometabolic assessments show beneficial effects of continuous estradiol treatment, observed both at time point 1 (prior to animal loss) and time point 2 (following animal loss). In future studies we will further investigate the timeline of increased mortality with estradiol exposure and explore its multifaced relationship with long-term health.
Finally, future work should examine the relationship between the effect of estradiol on cognitive aging and other measures of cardiometabolic health including low-density lipoprotein and total cholesterol, which is affected by loss of ovarian function in humans (75, 76) and rodent models (77, 78).
In conclusion, our study found no relationship between estradiol's effect on cognitive aging and cardiometabolic health from middle to old age in healthy ovariectomized rats. These results highlight that the cognitive benefits of estradiol are not secondary to its cardiometabolic effects, suggesting that estradiol influences cognitive systems and cardiometabolic systems independently. This insight is important because it challenges the view that cognitive improvements with estradiol may be a byproduct of cardiovascular and metabolic enhancements. Instead, our findings point to direct actions of estradiol on brain systems that support cognition, independent of metabolic and cardiovascular health, at least in the context of aging without preexisting disease.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Luine V, Frankfurt M. Estrogenic regulation of memory: the first 50 years. Horm Behav. 2020;121:104711.32035072 10.1016/j.yhbeh.2020.104711 PMC 7198346 · doi ↗ · pubmed ↗
- 2Stelly CE, Cronin J, Daniel JM, Schrader LA. Long-term oestradiol treatment enhances hippocampal synaptic plasticity that is dependent on muscarinic acetylcholine receptors in ovariectomised female rats. J Neuroendocrinol. 2012;24(6):887‐896.22313316 10.1111/j.1365-2826.2012.02287.x · doi ↗ · pubmed ↗
- 3Bohacek J, Bearl AM, Daniel JM. Long-term ovarian hormone deprivation alters the ability of subsequent oestradiol replacement to regulate choline acetyltransferase protein levels in the hippocampus and prefrontal cortex of middle-aged rats. J Neuroendocrinol. 2008;20(8):1023‐1027.18540996 10.1111/j.1365-2826.2008.01752.x · doi ↗ · pubmed ↗
- 4Duarte-Guterman P, Yagi S, Chow C, Galea LA. Hippocampal learning, memory, and neurogenesis: effects of sex and estrogens across the lifespan in adults. Horm Behav. 2015;74:37‐52.26122299 10.1016/j.yhbeh.2015.05.024 · doi ↗ · pubmed ↗
- 5Rettberg JR, Yao J, Brinton RD. Estrogen: a master regulator of bioenergetic systems in the brain and body. Front Neuroendocrinol. 2014;35(1):8‐30.23994581 10.1016/j.yfrne.2013.08.001PMC 4024050 · doi ↗ · pubmed ↗
- 6Pike CJ . Sex and the development of Alzheimer’s disease. J Neurosci Res. 2017;95(1-2):671‐680.27870425 10.1002/jnr.23827 PMC 5120614 · doi ↗ · pubmed ↗
- 7Scheyer O, Rahman A, Hristov H, et al Female sex and Alzheimer’s risk: the menopause connection. J Prev Alzheimers Dis. 2018;5(4):225‐230.30298180 10.14283/jpad.2018.34PMC 6198681 · doi ↗ · pubmed ↗
- 8Mc Carrey AC, Resnick SM. Postmenopausal hormone therapy and cognition. Horm Behav. 2015;74:167‐172.25935728 10.1016/j.yhbeh.2015.04.018PMC 4573348 · doi ↗ · pubmed ↗