Start, stop, resume, and proceed: ZmSSRP1 mediates the progression of RNA polymerase II and kernel development in maize
Christian Damian Lorenzo

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
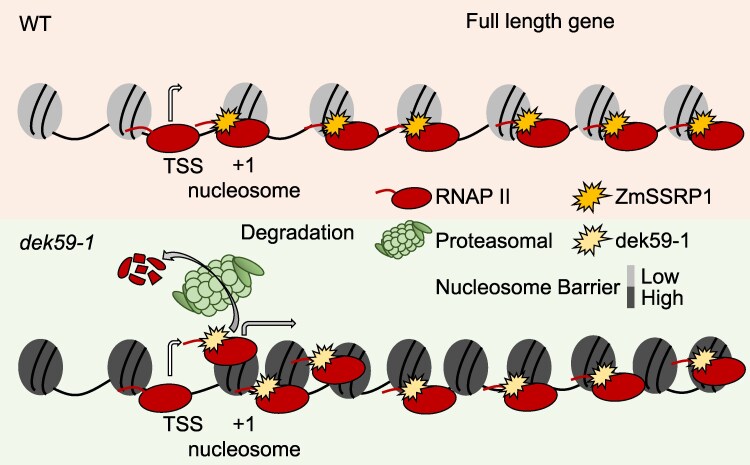 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic Mapping and Diversity in Plants and Animals · Plant Genetic and Mutation Studies · Rice Cultivation and Yield Improvement
Gene transcription is an essential process carried out by RNA Polymerase II (RNAP II) through a cycle of 4 key steps. It begins with initiation, followed by a promoter-proximal pausing phase, during which serine residues in the carboxy-terminal domain of RNAP II become phosphorylated. Next comes elongation, where the full RNA transcript is synthesized, and finally, termination, during which the mRNA is processed and the poly(A) tail is added (Liu et al. 2015). The journey of RNAP II through a gene is anything but easy, and the elongation step, particularly, can be influenced by several factors (Obermeyer et al. 2023). Nucleosomes (formed by DNA wrapped around an octamer of histones), for example, can hinder RNAP II movement. The complex FACILITATES CHROMATIN TRANSCRIPTION (FACT), composed of the 2 subunits STRUCTURE SPECIFIC RECOGNITION PROTEIN1 (SSRP1) and SUPPRESSOR OF TY (SPT16), promotes both nucleosome disassembly and reassembly, clearing the path for RNAP II to move along (Formosa and Winston 2020). FACT has been observed to co-purify with RNAP II and has also been implicated in the regulation of other steps of transcription (Tettey et al. 2019). Although RNAP II and FACT complex genes have been well characterized in Arabidopsis, the conservation of their functions in other plants is under investigation.
In recent research, Jin-Yu Wang and collaborators (Wang et al. 2025a) identified a maize mutant, dek59-1, with a reduced endosperm phenotype associated with a maize ortholog of SSRP1. dek59-1 was identified in an EMS screen due to its small kernels, resulting from a reduction in seed starch content. Positional cloning of dek59-1 led to a change between the wild type and the mutant in the last nucleotide of intron 12 of ZmSSRP1, a region critical for proper splicing. Wang et al. found that the mutation caused 2 types of misspliced events: one with a 16-nucleotide addition leading to a loss of function, and another with a 15-nucleotide deletion that led to a fully translated protein detectable by western blot. The authors further noticed that dek59-1 presented cell cycle–related problems, such as reduced cell number in endosperm tissue and disturbed endoreduplication values compared to the wild type. These results were supported by RNA-seq analysis between the wild type and dek59-1. Chromatin immunoprecipitation sequencing assays using a specific antibody against ZmSSRP1 revealed that this protein binds more to genic regions, displaying more occupancy on highly expressed genes. The group also observed through co-immunoprecipitation that ZmSSRP1 indirectly interacts with the maize RNAP II.
To investigate the nature of this interaction in vivo, the authors performed chromatin immunoprecipitation-seq assays in dek59-1 and wild-type lines using an antibody specific for RNAP II Ser2P. The mutation of ZmSSRP1 leads to a shift of RNAP II distribution toward the transcription starting site and decreases at the 3′ end of genes, suggesting reduced polymerase processivity. Additionally, chromatin accessibility in +1 nucleosomes (a barrier for RNAP II) is highly decreased in dek59-1 mutants as measured by ATAC-seq, hinting that ZmSSRP1 mutation could lead to reduced RNAP II progression due to less accessible chromatin. This effect was more substantial for longer genes than for shorter ones. Finally, the researchers also discovered that arrested ZmSSRP1 in dek59-1 is eliminated through proteasome-mediated degradation (see Figure).
Thus, Wang et al. have shed light on the conserved role of SSRP1 in transcript elongation in maize and how its misregulation can lead to severe phenotypes, opening the path to future studies of the pivotal functions of RNAP II and FACT complex members in other plant species.
Recent related articles in The Plant Cell
Liu et al. (2025) shed light on how 2 different transcription factors, MULTI-FLORET SPIKELET1 (TaMFS1) and the STRUCTURE-SPECIFIC RECOGNITION PROTEIN 1 (TaSSRP1), regulate VRT-A2 expression, a gene with important agronomical breeding potential in wheat. Wang et al. (2025b) described an abundance of alternative transcription starting sites present the in coding regions of several tissue-specific soybean genes. These alternative transcription starting sites within CDSs were epigenetically tuned and associated with tissue-specific functions. Sheng et al. (2025) isolated a gene COMPACT PLANT 3 (CT3), whose mutation leads to a compact plant architecture and increased plant gene in high-density stands. CT3 acts as a co-regulator interacting with other transcription factors and increasing their DNA binding affinity.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Formosa T, Winston F. The role of FACT in managing chromatin: disruption, assembly, or repair? Nucleic Acids Res. 2020:48(21):11929–11941. 10.1093/nar/gkaa 91233104782 PMC 7708052 · doi ↗ · pubmed ↗
- 2Liu J, Dong C, Liu X, Guo J, Chai L, Guo W, Ni Z, Sun Q, Liu J. Decoupling the pleiotropic effects of VRT-A 2 during reproductive development enhances wheat grain length and weight. Plant Cell. 2025:37(2):koaf 024. 10.1093/plcell/koaf 02439951393 PMC 11827615 · doi ↗ · pubmed ↗
- 3Liu X, Kraus WL, Bai X. Ready, pause, go: regulation of RNA polymerase II pausing and release by cellular signaling pathways. Trends Biochem Sci. 2015:40(9):516–525. 10.1016/j.tibs.2015.07.00326254229 PMC 4553066 · doi ↗ · pubmed ↗
- 4Obermeyer S, Schrettenbrunner L, Stöckl R, Schwartz U, Grasser KD. Different elongation factors distinctly modulate RNA polymerase II transcription in Arabidopsis. Nucleic Acids Res. 2023:51(21):11518–11533. 10.1093/nar/gkad 82537819035 PMC 10681736 · doi ↗ · pubmed ↗
- 5Sheng H, Zhang H, Deng H, Zhang Z, Qiu F, Yang F. Maize COMPACT PLANT 3 regulates plant architecture and facilitates high-density planting. Plant Cell. 2025:37(2):koaf 029. 10.1093/plcell/koaf 02939928526 PMC 11879032 · doi ↗ · pubmed ↗
- 6Tettey TT, Gao X, Shao W, Li H, Story BA, Chitsazan AD, Glaser RL, Goode ZH, Seidel CW, Conaway RC. A role for FACT in RNA polymerase II promoter-proximal pausing. Cell Rep. 2019:27(13):3770–3779.e 7. 10.1016/j.celrep.2019.05.09931242411 · doi ↗ · pubmed ↗
- 7Wang J-Y, Wang Q, Peng Y-X, Jiang L-G, Lu Z-Z, Zheng L-M, Li X-H, Liu J, Long J-C, Liu J-H, et al Zm SSRP 1 facilitates the progression of RNA polymerase II and is essential for kernel development in maize. Plant Cell. 2025 a:37(4). 10.1093/plcell/koaf 071PMC 1198328140166832 · doi ↗ · pubmed ↗
- 8Wang X, Duan J, Clark CB, Feng W, Ma J. Noncanonical transcription initiation is primarily tissue specific and epigenetically tuned in paleopolyploid plants. Plant Cell. 2025 b:37(1):koae 288. 10.1093/plcell/koae 288PMC 1166355539540911 · doi ↗ · pubmed ↗