Gametocyte production and transmission fitness of African and Asian Plasmodium falciparum isolates with differential susceptibility to artemisinins
Nicholas I. Proellochs, Chiara Andolina, Jordache Ramjith, Rianne Stoter, Geert-Jan van Gemert, Wouter Graumans, Susana Campino, Leen N. Vanheer, Martin Okitwi, Patrick K. Tumwebaze, Melissa D. Conrad, Taane G. Clark, David A. Fidock, Didier Ménard, Sachel Mok, Teun Bousema

TL;DR
This study compares how African and Asian malaria parasites with different resistance to artemisinin drugs produce and transmit gametocytes in mosquitoes.
Contribution
The study reveals transmission fitness differences among Plasmodium falciparum isolates with varying artemisinin resistance and K13 genotypes.
Findings
ART-sensitive isolates consistently produced gametocytes and transmitted effectively in all tested mosquito species.
ART-R isolates showed variable transmission potential, with some maintaining high infectivity despite resistance.
DHA exposure reduced mosquito infectivity for all isolates regardless of K13 genotype.
Abstract
The emergence of Plasmodium falciparum parasites partially resistant to artemisinins (ART-R) poses a significant threat to recent gains in malaria control. ART-R has been associated with PfKelch13 (K13) mutations, which differ in fitness costs. This study investigates the gametocyte production and transmission fitness of African and Asian P. falciparum isolates with different K13 genotypes across multiple mosquito species. We tested three ART-sensitive (ART-S) isolates (NF54, NF135, and NF180) and three ART-R isolates (ARN1G, 3815, and PAT-023) for sexual conversion and transmission to Anopheles stephensi, An. gambiae, and An. coluzzii. ART-R levels were quantified in vitro using the Ring-stage Survival Assay (RSA), and the transmission-reducing effects of dihydroartemisinin (DHA) on mature gametocytes were assessed. Results showed that ART-S parasite lines consistently produced…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
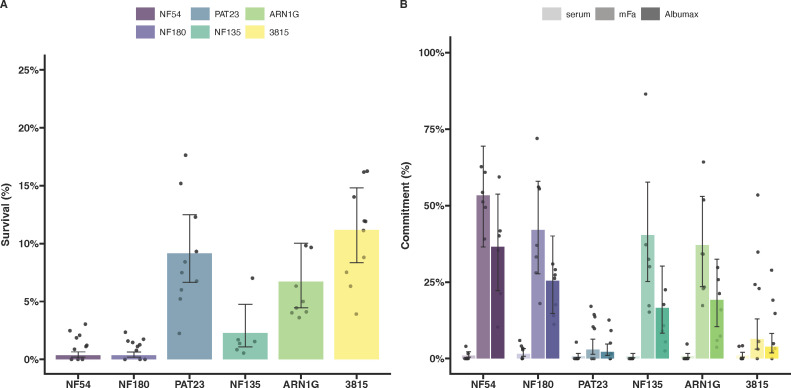 Fig 1
Fig 1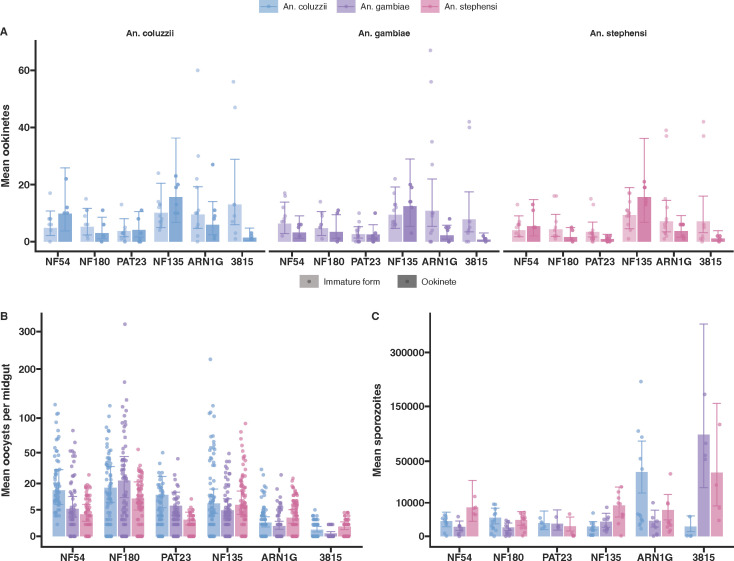 Fig 2
Fig 2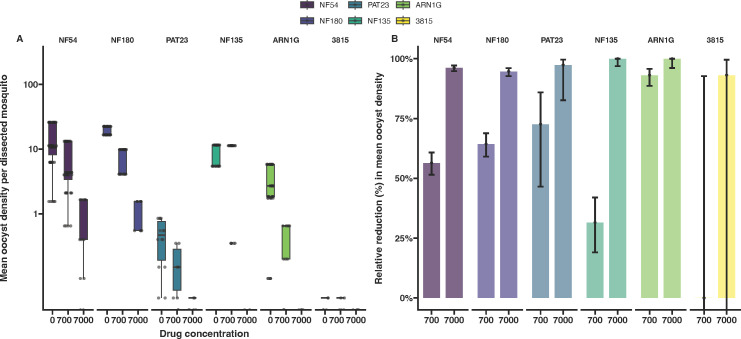 Fig 3
Fig 3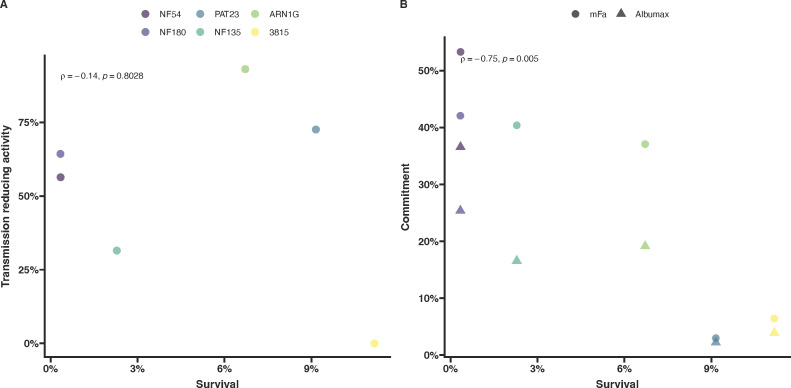 Fig 4
Fig 4| Parasite isolate | Origin (Year) | Artemisinin resistance phenotype | Drug resistance-conferring mutations | ||||
|---|---|---|---|---|---|---|---|
| Artemisinin | Chloroquine | Mefloquine | Pyrimethamine | Sulfadoxine | |||
| NF54 | Africa (1979) | S | – | – | – | – | – |
| NF180 | East Africa (Uganda 2016) | S | – | – | – | DHFR (N51I, S108N) | DHPS (A437G, K540E) |
| PAT-023 | East Africa (Uganda 2021) | R | ART-R | PfMDR1 (Y184F) | – | DHFR (C59R, S108N) | DHPS (A437G, K540E) |
| NF135 | Southeast Asia (Cambodia 1993) | S | – | PfCRT (M74I, N75E, K76T, A220S, Q271E, I356T, R371I) | – | DHFR (N51I, C59R, S108N, I164L) | DHPS (S436A, A437G, K540E) |
| ARN1G | Southeast Asia (Thailand 2011–2013) | R | K13 (G449A) | PfCRT (M74I, N75E, K76T, A220S, Q271E, N326S, I356T, R371I) | – | DHFR (N51I, C59R, S108N, I164L) | DHPS (A437G, K540E, A581G) |
| 3815 | Southeast Asia (Cambodia 2010) | R | K13 (C580Y) | PfCRT (M74I, N75E, K76T, A220S, Q271E, I356T, R371I) | – | DHFR (N51I, C59R, S108N, I164L) | DHPS (S436A, A437G, K540E) |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Mosquito-borne diseases and control · Invertebrate Immune Response Mechanisms
INTRODUCTION
Despite global efforts to reduce the malaria burden, progress has plateaued in the last decade, and in some areas, malaria is again increasing (1). The recent emergence in Sub-Saharan Africa of Plasmodium falciparum parasites with partial resistance to artemisinins (ART-R) adds to the concerns about how sustainable malaria control may be. ART-R is characterized by prolonged parasite clearance times (half-life >5 h) or persistence of parasitemia on day 3 following artemisinin monotherapy or artemisinin-based combination therapy (ACT) (2–6). ART-R is predominantly associated with specific mutations in the essential PfKelch13 (K13) protein’s β-propeller domain (7–10). These mutations reduce K13 protein levels, disrupting hemoglobin import into the parasite’s digestive vacuole, a critical step for ART activation (8, 11–13). While there are validated K13 mutations that associate with ART-R, their impacts on resistance and parasite fitness are variable and highly dependent on the mutation and parasite background (13–16); as such, it is difficult to predict which mutations are of particular concern.
Clinically relevant mutations in K13 first were observed in isolates collected in 2002 in western Cambodia (17). While partial resistance subsequently spread across the Greater Mekong Subregion (7, 18), the anticipated migration to the African continent has not occurred. It was hypothesized that lower drug pressures combined with high parasite diversity in many settings in Sub-Saharan Africa may have allowed wild-type parasites to outcompete less-fit K13 mutant parasites, making de novo emergence of ART-R less likely and limiting the spread of the K13 mutants upon introduction. The recent independent emergence of K13 mutations associated with ART-R including in East Africa (Uganda, Rwanda, and Tanzania) and the Horn of Africa (Eritrea, Ethiopia, and Sudan) (19–23) raises questions about their fitness, resistance, and transmission potential. Transmission to mosquitoes depends on the formation of viable male and female gametocytes, with considerable variation in gametocyte production between parasite isolates (24, 25). The transmission potential of parasites with K13 mutations is particularly relevant with changes in vector populations and variation in P. falciparum vector competence (26). The recent invasion of the competent Asian vector Anopheles stephensi in urban African settings (27, 28) raises concerns about the spread of ART-R parasites of African and Asian origin in settings previously not endemic for malaria.
Understanding variations in vector competence and determining whether K13 mutant parasites possess a transmission advantage, both in the presence and absence of artemisinin derivatives, are crucial for predicting the potential spread of ART-R on the African continent. In this study, we investigate the gametocyte production and transmission fitness of ART-S and ART-R field isolates. We compared ART-S isolates (29, 30) with ART-R isolates carrying two common Southeast Asian K13 mutations G449A and C580Y (18, 31, 32), as well as a recently isolated ART-R K13 independent parasite line from Uganda, where the local emergence of ART-R has been documented, including through K13 mutations.
MATERIALS AND METHODS
Parasite cultures
All parasite lines, including the ART-S NF54 (Africa), NF135 (Cambodia) and NF180 (Uganda), as well as the ART-R P. falciparum field isolates ARN1G (Thailand), 3815 (Cambodia), and PAT-023 (Uganda) were cultured in an automated culture system using RPMI media supplemented with 10% human serum, with gametocyte cultures for mosquito feeding set up at a uniform 1% parasitemia and allowed to mature for 14 days prior to feeding (33). Asexual parasite cultures were maintained for no more than 25 cycles in a synchronized state through magnet separation or sorbitol lysis. Gametocyte induction was performed using either minimal fatty acid media or 0.5% Albumax; gametocytes were allowed to mature in RPMI media supplemented with 10% human serum (25). Parasite DNA was extracted with the QIAGEN Blood DNA kit, sequenced on the Illumina Novaseq 6000 platform, and analysed using the malaria profiler tool (34).
Ring-stage Survival Assay (RSA)
The Ring-stage Survival Assay (RSA) was performed as previously published (16, 35) with minor modifications. To avoid the negative impact of sorbitol synchronisation on conversion rates within the same cycle (36, 37), we opted for a gentler approach using double magnet purification. Highly synchronized parasites were first passed through a magnetic column (MACS) to collect segmented schizonts. These schizonts were put back into culture for 3–4 h to allow bursting and reinvasion of new red blood cells (RBCs). The culture was subsequently passed through the same magnetic column, isolating the flow-through, which contained only newly invaded ring-stage parasites. Parasites were split into two plates: one for the sexual conversion assay and one for the RSA. For the RSA, parasites were diluted to 1%–2% parasitemia in 2% hematocrit. Each line was tested in four wells: two for DMSO controls and two for DHA. Washes were performed in separate tubes following DHA treatment before being added to a fresh plate. Survival rates were determined using both Giemsa smears and flow cytometry using MitoTracker Red as a live cell stain from the same well for all replicates.
Sexual conversion assay
Sexual conversion rates were performed as previously published (25), with minor modifications. After double synchronization, parasites were returned to culture for 24 h prior to initiating the conversion assay. The assay was performed in a plate-based format with each parasite isolate split over three wells (one for each media type) at 1% parasitemia in 5% hematocrit. Conversion rates were calculated by Giemsa-stained smears (25).
Mosquito infections
Laboratory colonies of An. stephensi (Nijmegen Sind-Kasur strain) (38), An. coluzzii (N’gousso strain) (39), and An. gambiae s.s (Kisumu strain) (40) were maintained under controlled conditions: 26°C, 70%–80% humidity and a 12 h reverse day/night cycle. Gametocyte cultures were checked for exflagellation to confirm maturation and gametocytaemia reached a minimum of 0.1% before feeding. Culture material was not further normalized for gametocyte density prior to feeding. Groups of 100 female Anopheles stephensi, An. gambiae, and An. coluzzii mosquitoes, aged 1–5 days, were blood-fed for 15 min using glass membrane mini-feeders (15 mm diameter, convex bottom) connected to a heated circulating water bath. Fully fed mosquitoes were selected and maintained at 30°C with access to 5%–10% glucose.
Ookinete count
Mosquito midguts were examined 18–24 h post-infection to identify the presence of round forms, retort forms, and mature ookinetes. Five midguts from each infection group were dissected and incubated with a 1:50 diluted Anti-25KD-FITC conjugate in Evans blue solution. Midguts were gently disrupted using a pipette tip to release the blood meal. Following incubation in the dark for 30 min at room temperature, the solution was washed with 1.4 mL of phosphate-buffered saline (PBS), vortexed to dissolve the pellet, and centrifuged for 2 min at 10,000 rpm. After removal of the supernatant, the pellet was resuspended in 25 µL of PBS, and 5 µL of the suspension was loaded into a Bürker–Turk counting chamber. Round forms, retort forms, and mature ookinetes were counted using an incident light fluorescence microscope with a GFP filter at 400× magnification.
Oocyst count
Seven days post-infection, 20 mosquito midguts per group were dissected, stained with 1% mercurochrome, and examined under an optical microscope at 100× magnification to detect and quantify oocysts.
Sporozoite count
Mosquitoes used to examine sporozoite development received a second (uninfected) bloodmeal on day 4–6 post-infection to synchronize oocyst development. Fifteen days post-infection, mosquito salivary glands were dissected in PBS and transferred to oocyst lysis buffer (NaCl 0.1M: EDTA 25 mM: TRIS-HCl 10 mM). Following overnight incubation at 56°C with Proteinase K, DNA was extracted with the automated MagNA Pure LC instrument using the MagNA Pure LC DNA Isolation Kit–High performance. Sporozoite density was analyzed using COX-I qPCR (41).
Statistical analysis
Statistical analyses were conducted using R software (v 3.1.12) (42). Mean counts of oocysts, sporozoites, and ookinetes (with 95% confidence intervals) were estimated using a mixed-effects negative binomial regression model, incorporating random intercepts for biological replicates and fixed effects for parasite line, mosquito species, and their interactions. Mean survival and commitment rates (with 95% confidence intervals) were calculated using a mixed-effects beta regression model. Transmission-reducing activity (TRA) and transmission-blocking activity (TBA)—reflecting reductions in oocyst density and the proportion of infected mosquitoes, respectively—were assessed for 700 and 7,000 nM DHA using a Bayesian Poisson regression model.
RESULTS
Parasite backgrounds
We used well-characterized P. falciparum reference lines from Africa (NF54), Southeast Asia (NF135, Cambodia) (29), and a new isolate from East Africa (NF180, Uganda). These lines were complemented with previously published ART-R K13 mutant parasite lines from Thailand (ARN1G, K13-449A) (32), from Cambodia (3815, K13-580Y) (31, 35) and a recently collected isolate from Uganda with in vitro ART-R that is independent of K13 mutations (PAT-023). Genotyping revealed that some isolates harbored mutations associated with resistance to chloroquine and sulfadoxine-pyrimethamine (Table 1).
Artemisinin resistance
ART-R lines ARN1G and 3815 showed survival rates in the RSA that were higher than the ART-S NF54, as previously published [(16, 32, 35); Fig. 1A; Table S1]. The ART-S NF135 line showed variable survival rates, which were lower than those observed for known ART-R lines but higher than those observed for ART-S NF54 and NF180 lines. The PAT-023 parasite line exhibited a survival rate of 9.1% (95% CI: 6.6%–12.5%), higher than the ART-S lines.
Parasite survival after exposure to dihydroartemisinin (DHA) and gametocyte conversion rates of P. falciparum isolates with different K13 genotypes and genetic backgrounds. (A) Survival rate from the Ring-stage Survival Assay (RSA) of each parasite line following 6 h exposure to 700 nM DHA compared with DMSO controls in duplicate wells (4–6 replicates were performed for each parasite line). The dots represent biological replicate survival rates at 72 h post invasion from counts either by microscopy or flow cytometry, and the error bars represent the 95% confidence intervals. (B) Sexual conversion in a plate-based assay that quantifies conversion rates from a single asexual round using three different media types (shown in different color gradient of the bars). Conversion rates are calculated by dividing the final gametocytemia by the starting parasitemia in the same well. Dots represent a single well from independent plates; error bars represent 95% confidence intervals.
Sexual conversion
We assessed the ability of ART-R and ART-S lines to convert under two different conditions that mimic the natural sensing system (25, 43, 44). All conversion assays were performed on a single asexual cycle. The highest rates of gametocyte conversion were typically observed using minimal fatty acid media with intermediate conversion rates for media containing Albumax only, and lowest conversion in non-inducing serum media (Fig. 1B; Table S2) (25). The conversion rates observed for K13 mutants were comparable to those recorded for ART-S lines, except for the Cambodian line 3815, which exhibited highly variable conversion rates (Fig. 1B). The Ugandan ART-R line PAT-023 exhibited minimal sexual conversion with little difference between media types (Fig. 1B; Table S2).
Transmission to mosquitoes
Transmission was examined using three mosquito vector species. We first assessed whether fertilization occurred in mosquitoes for each parasite isolate by quantifying parasite developmental stages in the mosquito midgut 20–24 h post-infectious blood meal. We distinguished between mature and immature stages following Pfs25 antibody labeling. Immature stages included underdeveloped ookinetes and rounded forms classified as either zygotes or unfertilized females. The ART-S parasite isolates NF180 and NF135 showed no clear difference in the numbers of immature or mature ookinetes across the mosquito species tested (Fig. 2A). For NF54, there was a tendency for fewer mature ookinetes in An. gambiae compared with An. stephensi and An. coluzzii, though this was not statistically significant (Fig. 2A). A similar trend was observed with the ART-R ARN1G parasite line. For the Ugandan ART-R isolate PAT-023, significantly fewer mature ookinetes were observed in An. stephensi (Fig. 2A) compared to An. coluzzii (P = 0.019). However, this significance level should be interpreted cautiously due to the number of comparisons made. The ART-R isolate 3815 consistently produced low numbers of mature ookinetes in all Anopheles species tested (Fig. 2A).
Infection of three mosquito species with the six different parasite lines. (A) Ookinete counts 20 h after blood meal. The ookinetes were stained with anti-Pfs25 488 conjugate and counted on a hemocytometer under a fluorescent microscope. Immature forms included rounded zygotes (or unfertilized females) and ookinetes with incomplete maturation (lighter shading bar), while mature forms were only the completely mature ookinetes (dark shaded bar). Dots represent a single count from an independent experiment, and the error bars represent 95% confidence intervals. (B) Average oocysts per midgut were dissected on day 7 post-bloodmeal. A total of 20 mosquitoes per group were dissected, and oocysts were counted by mercurochrome staining. Dots represent the counted oocysts from a single midgut; error bars represent 95% confidence intervals. (C) Salivary glands from individual mosquitoes were dissected, and sporozoites were quantified by qPCR. The dots represent the sporozoites per salivary gland from a single mosquito, and the error bars represent the confidence intervals.
To confirm that mature ookinetes can establish mosquito infections, we quantified oocyst prevalence (Fig. S1) and density (Fig. 2B) on day 7 post-bloodmeal. African parasite lines (NF54, NF180, and PAT-023) exhibited higher oocyst densities in traditional African mosquitoes (An. gambiae s.s., An. coluzzii) compared with An. stephensi, particularly for PAT-023 (P < 0.0001; Table S3). In contrast, NF135 showed consistent infection intensities across all tested mosquito species. For the Asian K13 mutant lines, oocyst densities in An. stephensi were slightly higher compared with An. coluzzii and An. gambiae. Specifically, ARN1G had higher densities in An. stephensi relative to An. coluzzii (P = 0.009) and An. gambiae (P < 0.0001), while 3815 showed similar trends (P = 0.007 compared with An. coluzzii and P < 0.0001 compared with An. gambiae; Table S3). The 3815 isolate exhibited low and highly variable infection levels, consistent with its reduced mature ookinete counts (Fig. 2A and B). Interestingly, a trend towards higher sporozoite production was observed in the two K13 mutant lines (ARN1G and 3815), which persisted even after normalizing for oocyst density within the same batch of mosquitoes. However, transmission results for the 3815 isolate remained highly variable (Fig. S2).
Transmission-blocking effect of DHA
With all parasite lines being able to infect mosquitoes in the absence of drug pressure, we determined the transmission-reducing effect of DHA on mosquito infection intensity and prevalence. Mature gametocytes were exposed to both the physiologically relevant concentration of 700 nM and a 10-fold higher concentration of 7,000 nM for 48 h before mosquito feeding with the drug not removed before mosquito feeding (32, 45). Given the labor-intensive nature of these assays and the known permissiveness of An. coluzzii for the parasites tested herein, experiments were performed exclusively with this mosquito species. All isolates showed a consistent decrease in mosquito infection intensity and prevalence as DHA concentrations increased, independent of the K13 genotype and the parasite background (Fig. 3; Fig. S3). At 7,000 nM DHA, sporadic infections were observed for the African isolates NF54, NF180, and PAT-023, but none of the Asian isolates infected mosquitoes at this concentration. When relative reductions in oocyst intensity were calculated, DHA exposure at 7,000 nM reduced oocyst intensities by >93% for all parasite isolates. There were no indications of reduced transmission-blocking efficacy of DHA for ART-R isolates.
Transmission reduction in the presence of DHA. (A) Oocyst densities observed upon exposing mature gametocytes to 700 and 7,000 nM DHA prior to being fed to mosquitoes in a blood meal. The dots represent the average oocysts per infected mosquito from a single cage. (B) Relative reductions in oocyst density compared with the no drug control. All error bars represent the confidence intervals.
Correlating resistance and transmission stages
Although the number of parasite isolates examined was modest, our data on asexual parasite survival under DHA pressure, combined with findings on sexual conversion and mosquito transmission, provide an opportunity to investigate whether ART-R confers a transmission advantage. We initially hypothesized that parasites with high levels of in vitro resistance to ART at the ring stage would show enhanced transmission to mosquitoes under drug pressure. However, no correlation was observed between parasite survival in the RSA and the reduction in oocyst density under DHA pressure (Fig. 4A). In contrast with our initial hypothesis, we observed a weakly negative correlation between parasite survival rates in the RSA and gametocyte conversion rates upon induction (Fig. 4B).
Impact of resistance on transmission and commitment. Transmission (A) and conversion (B) plotted against survival rates from the RSA for each parasite line. The underlying detailed data on in vitro parasite resistance in the RSA and gametocyte commitment are presented in Fig. 1; data on transmission under DHA exposure are presented in Fig. 3.
DISCUSSION
Understanding the intricacies of transmission fitness of parasites that survive ART-based treatment is important for developing strategies that aim to prevent or slow down the spread of resistance. Some reports have suggested that K13 mutant parasites may have higher intrinsic gametocyte production (4, 46), yet some in vivo evidence suggests that following ART treatment in a Vietnam cohort, the conversion marker ap2g decreases—suggesting a decrease in gametocyte production linked to K13 mutations (47). In contrast, an in vitro study observed increased survival of specifically male gametocytes of K13 mutants under DHA exposure (32).
We selected three parasite isolates from Southeast Asia and three from Sub-Saharan Africa, each with distinct K13 genotypes and genetic backgrounds. The Asian ART-R lines ARN1G and 3815 exhibited higher parasite survival in the RSA, consistent with previous studies (16, 32, 35). PAT-023, a newly characterized Ugandan ART-R line that is wild-type for K13, demonstrated a high level of survival in the in vitro RSA. Interestingly, we also observed that NF135, a line previously associated with treatment failure following artemether-lumefantrine therapy in vivo (48), showed erratic and occasionally increased DHA survival rates.
This study focuses on the transmission potential of P. falciparum isolates, beginning with their commitment to sexual stages. Fatty acids play a key role in signaling the parasite to commit to sexual conversion (36, 37); we used two methods targeting this signaling pathway to robustly compare sexual conversion rates. From all media conditions tested, minimal fatty acid media consistently produced the highest sexual conversion rates across all lines. Among the two ART-S parasite isolates, NF54 and NF135 consistently exhibited high conversion levels. The newly introduced NF180 line, which has not been characterised in detail previously, demonstrated high conversion rates, comparable to NF54 and higher than NF135. Among the ART-R parasite lines, PAT-023 displayed the lowest commitment to gametocyte production, while ARN1G showed commitment levels similar to NF135. The Cambodian 3815 line exhibited an inconsistent commitment to gametocyte production, with occasional high levels that decreased with prolonged culture, potentially reflecting epigenetic silencing during in vitro cultivation.
We observed no evidence of increased sexual conversion in the ART-R isolates in the absence of treatment. Of note, we did not explore the possible impact of DHA on conversion rates in these ART-R lines. An earlier study reported that the ART-S NF54 line was associated with higher sexual conversion when trophozoites were treated with DHA (49). Future studies that investigate whether DHA-treated ART-R lines may give rise to an elevated conversion rate can be of value. Such studies would allow us to disentangle the contributions to the overall transmission competence; if this is linked to the increased conversion to or survival of the sexual stages after ART treatment or an inherent increased conversion rate in the K13 mutant lines.
While sexual conversion is a critical step for transmission, high commitment rates do not always correlate with high mosquito infectivity (25, 29). The presence of mature ookinetes is a direct indicator of fertilisation in the mosquito environment. In our study, we fed the same gametocyte material to the three mosquito species. We did not normalize for gametocyte density prior to feeding; instead, we had a uniform starting parasite concentration prior to gametocyte induction. We are thus able to compare the composite product of gametocyte production and infectivity between lines. When examining the impact of individual mutations on transmission competence in matched genetic backgrounds, such normalization at different points in the transmission cycle may be beneficial. We reproducibly observed low transmission potential of the 3815 line, which consistently formed very few ookinetes in all species and exhibited low and sporadic oocyst infections. We also observed a tendency for African parasites to fare better in the African mosquitoes, illustrated by both the numbers of mature ookinetes and oocyst densities. Although our sample size is too small for a comprehensive analysis of the underlying biological mechanisms, one could speculate that genes influencing the parasite’s ability to evade the mosquito immune system may play a role. Pfs47 is essential for P. falciparum infection in An. gambiae but not for An. stephensi (50, 51). Contrary to this observation for African parasite lines, all three Southeast Asian parasite lines were able to infect Asian and African mosquito vectors. Interestingly, ARN1G achieved slightly higher ookinete numbers in An. coluzzii, but this did not translate into higher oocyst densities. Similar to our observations on sexual conversion rates, we found no significant transmission advantage for ART-R parasite lines with reduced ART susceptibility in the RSA.
To confirm the ability of the parasites to complete sporogony, we quantified sporozoite numbers in mosquito salivary glands. Previous studies have suggested that K13 mutant lines might produce larger oocysts (32), potentially indicating increased sporozoite production. We did not observe a clear increase in oocyst size (data not shown), while sporozoite production varied between parasite lines. The two Asian ART-R lines, each with a unique K13 mutation, exhibited significantly higher sporozoite loads in at least one African mosquito species. This effect was most pronounced for ARN1G that showed significantly higher sporozoite numbers in An. coluzzii, even after adjusting for oocyst density within the same mosquito batch. Further studies are needed to confirm these findings and to explore the underlying biological mechanisms. However, our results are consistent with the suggestion that the K13 mutation may lead to larger oocysts (32), which could lead to increased sporozoite production.
Finally, we examined the impact of DHA exposure on transmission efficiency, hypothesizing that parasite lines with increased survival rate of asexual parasites under DHA exposure may show similar survival advantages when gametocytes are exposed. Previous work has suggested that male gametocytes with K13 mutations may have partial protection against DHA (32, 52). Using An. coluzzii, we tested two DHA concentrations and found no evidence that DHA was less effective against mature gametocytes and subsequently reducing transmission in ART-R parasites. Immature gametocytes are known to be more susceptible to artemisinins than mature gametocytes (32, 53). Since we did not examine the impact of DHA on immature gametocytes, we cannot rule out preferential survival of immature ART-R gametocytes. Even at the highest DHA concentrations, both ART-S and ART-R African parasite lines occasionally infected mosquitoes. This observation is consistent with previous ex vivo experiments (45) and aligns with clinical studies showing that treatment with DHA-piperaquine fails to fully prevent transmission in the first weeks after treatment (54).
In this study, we examined a limited number of ART-S and ART-R parasite lines across three mosquito species. We observed no consistent evidence of increased gametocyte production or increased mosquito infectivity. While the transmission-reducing effect of DHA was imperfect, its efficacy was not reduced in gametocyte-producing lines with partial resistance to ARTs. To fully uncover the discrete impacts of individual K13 mutations on gametocyte production, infectivity, and gametocyte resistance to ARTs, future studies using genetically engineered lines in controlled isogenic backgrounds are required.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1WHO. 2024. World malaria report.
- 2Noedl H, Se Y, Schaecher BL, Socheat D, Fukuda MM. 2008. Evidence of artemisinin-resistant malaria in Western Cambodia. N Engl J Med 359:2619–2620. doi:10.1056/NEJ Mc 080501119064625 · doi ↗ · pubmed ↗
- 3Dondorp AM, Nosten F, Yi P, Das D, Phyo AP, Tarning J, Lwin KM, Ariey F, Hanpithakpong W, Lee SJ, Ringwald P, Silamut K, Imwong M, Chotivanich K, Lim P, Herdman T, An SS, Yeung S, Singhasivanon P, Day NPJ, Lindegardh N, Socheat D, White NJ. 2009. Artemisinin resistance in Plasmodium falciparum malaria. N Engl J Med 361:455–467. doi:10.1056/NEJ Moa 080885919641202 PMC 3495232 · doi ↗ · pubmed ↗
- 4Amaratunga C, Lim P, Suon S, Sreng S, Mao S, Sopha C, Sam B, Dek D, Try V, Amato R, Blessborn D, Song L, Tullo GS, Fay MP, Anderson JM, Tarning J, Fairhurst RM. 2016. Dihydroartemisinin-piperaquine resistance in Plasmodium falciparum malaria in Cambodia: a multisite prospective cohort study. Lancet Infect Dis 16:357–365. doi:10.1016/S 1473-3099(15)00487-926774243 PMC 4792715 · doi ↗ · pubmed ↗
- 5Mairet-Khedim M, Leang R, Marmai C, Khim N, Kim S, Ke S, Kauy C, Kloeung N, Eam R, Chy S, Izac B, Mey Bouth D, Dorina Bustos M, Ringwald P, Ariey F, Witkowski B. 2021. Clinical and in vitro resistance of Plasmodium falciparum to artesunate-amodiaquine in Cambodia. Clin Infect Dis 73:406–413. doi:10.1093/cid/ciaa 62832459308 PMC 8326543 · doi ↗ · pubmed ↗
- 6Phyo AP, Ashley EA, Anderson TJC, Bozdech Z, Carrara VI, Sriprawat K, Nair S, White MM, Dziekan J, Ling C, Proux S, Konghahong K, Jeeyapant A, Woodrow CJ, Imwong M, Mc Gready R, Lwin KM, Day NPJ, White NJ, Nosten F. 2016. Declining efficacy of artemisinin combination therapy against P. falciparum malaria on the Thai-Myanmar border (2003-2013): the role of parasite genetic factors. Clin Infect Dis 63:784–791. doi:10.1093/cid/ciw 38827313266 PMC 4996140 · doi ↗ · pubmed ↗
- 7Ashley EA, Dhorda M, Fairhurst RM, Amaratunga C, Lim P, Suon S, Sreng S, Anderson JM, Mao S, Sam B. 2014. Spread of artemisinin resistance in Plasmodium falciparum malaria. N Engl J Med 371:411–423. doi:10.1056/NEJ Moa 131498125075834 PMC 4143591 · doi ↗ · pubmed ↗
- 8Birnbaum J, Scharf S, Schmidt S, Jonscher E, Hoeijmakers WAM, Flemming S, Toenhake CG, Schmitt M, Sabitzki R, Bergmann B, Fröhlke U, Mesén-Ramírez P, Blancke Soares A, Herrmann H, Bártfai R, Spielmann T. 2020. A Kelch 13-defined endocytosis pathway mediates artemisinin resistance in malaria parasites. Science 367:51–59. doi:10.1126/science.aax 473531896710 · doi ↗ · pubmed ↗