Plasmodium falciparum Kelch-13 artemisinin partial resistance markers in Fort Portal, Western Uganda, 2024
Welmoed van Loon, Emma Schallenberg, Emmanuel Mande, Patrick Musinguzi, Paul Ngobi, Sharon Atukunda, John Rubaihayo, Frank P. Mockenhaupt

TL;DR
The study found early signs of artemisinin resistance in malaria parasites in Fort Portal, Uganda, near known resistance hotspots.
Contribution
The study reports the presence of Kelch13 mutations in Uganda, suggesting the incipient emergence of artemisinin resistance.
Findings
Artemisinin resistance-associated mutations in Kelch13 were present in 4.8% of samples.
A mutation of unknown relevance (490T) occurred in 9.5% of samples.
PfMDR1 variants suggested increased lumefantrine tolerance (N86, 100%).
Abstract
In Fort Portal, Uganda, artemisinin resistance-associated mutations in Plasmodium falciparum Kelch13 (n = 126) were present in 4.8% (675V, 561H, and 441L). A mutation of unknown relevance, 490T, occurred in 9.5%. PfMDR1 variants suggested increased lumefantrine tolerance (N86, 100%). Mutation 500N was absent, and 199S occurred in 12.8%. The latter is of unknown relevance. These data indicate an incipient emergence of artemisinin resistance in a crucial location between Rwandan and Ugandan resistance hotspots and hardly affected DR Congo.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
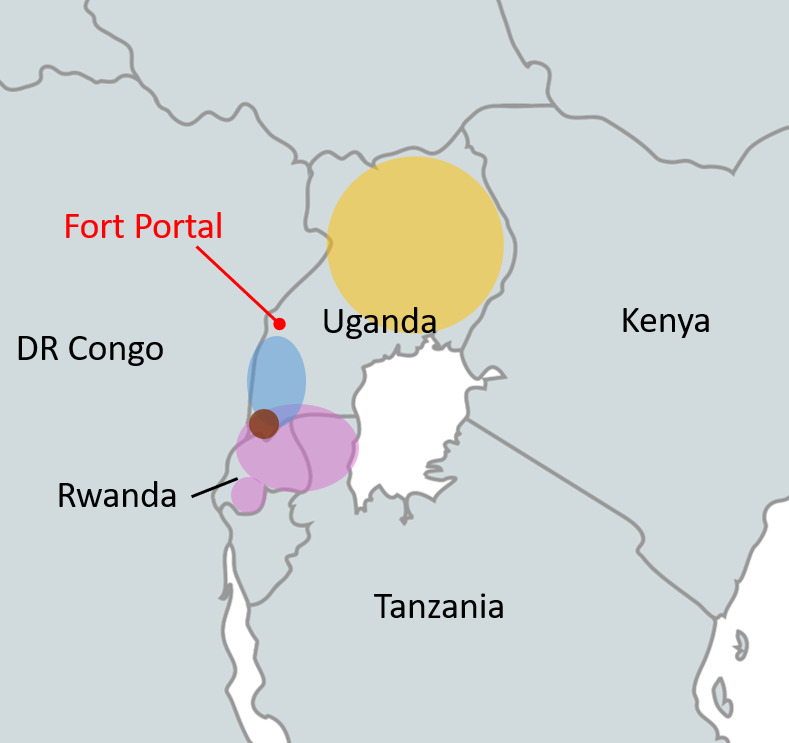 Fig 1
Fig 1| Gene | Mutation | No. reads | Prevalence mutant or mixed | Mixed | |
|---|---|---|---|---|---|
| (%, 95% CI) | (n) | (n) | |||
|
| 441L | 126 | 1.6 (0.4–5.6) | 2 | 0 |
| 490T | 9.5 (5.5–15.9) | 12 | 6 | ||
| 561H | 1.6 (0.4–5.6) | 2 | 0 | ||
| 675V | 1.6 (0.4–5.6) | 2 | 0 | ||
| 86Y | 102 | 0 | 0 | 0 | |
| 184Y | 62.8 (53.1–71.5) | 64 | 13 | ||
| 199S | 12.8 (7.6–20.6) | 13 | 2 | ||
| 500N | 125 | 0 | 0 | 0 | |
| 1246Y | 47 | 4.3 (0.5–14.5) | 2 | 0 | |
| N86-184F-D1246 (NFD) | 43 | 60.5 (44.4–75.0) | 26 | N/A | |
| Fort Portal regional referral hospital (0° 39' 10.79" N, 30° 16' 33" E) | Rukunyu Hospital (0° 15' 31" N, 30° 26' 58" E) | Kibiito Health Centre IV (0°28'39.45" N, 30°11'43.1" E) (%, n/N) | Karugutu Health Centre IV (0°47'31.64" N, 30°13'36.7" E) (%, n/N) | |
|---|---|---|---|---|
| WT | 71.4, 15/21 | 90.9, 20/22 | 92.3, 36/39 | 84.1, 37/44 |
| 441L | 4.8, 1/21 | 4.5, 1/22 | 0, 0/39 | 2.3, 1/44 |
| 490T | 23.8, 5/21 | 4.5, 1/22 | 0, 0/39 | 13.6, 6/44 |
| 561H | 0, 0/21 | 0, 0/22 | 2.6, 1/39 | 0, 0/44 |
| 675V | 0, 0/21 | 0, 0/22 | 5.1, 2/39 | 0, 0/44 |
- —Else Kroener-Fresenius-Stiftunghttp://dx.doi.org/10.13039/501100003042
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Computational Drug Discovery Methods · Parasites and Host Interactions
INTRODUCTION
Uganda, among the top three countries in malaria burden globally (1), is a hotspot of emerging partial artemisinin resistance (AR) of Plasmodium falciparum in East Africa (2). Malaria control relies on effective treatment with artemisinin combination therapy (ACT), composed of a fast-acting artemisinin derivative and a slower-acting partner drug, for example, lumefantrine or amodiaquine. Combination treatment is meant to prevent resistance. However, experience from Southeast Asia shows that once AR has emerged, resistance against the long half-life partner drugs can develop within years, resulting in clinical ACT treatment failure (3, 4). So far, this has not been observed in Africa. However, in Uganda, lumefantrine tolerance is increasing (5).
Molecular surveillance of resistance markers has a central role in the response to AR, helping identify hotspots and geospatial patterns. Certain mutations in the P. falciparum Kelch-13 Propeller Domain (PfK13) associate with AR, referred to as validated or candidate PfK13 markers. Tolerance to lumefantrine and amodiaquine is mediated by mutations in the P. falciparum multi-drug resistance 1 gene (PfMDR1), each drug exerting opposing selective effects (6–8).
The aim of the present study was to contribute to unveiling the current situation of AR in a region of Uganda that has been little researched to date. The study area of Fort Portal, far western Uganda, is located between Ugandan and Rwandan AR hotspots (2, 9, 10), and so far hardly affected eastern DR Congo (10, 11). This area may potentially bridge P. falciparum populations with and without AR (Fig. 1). We genotyped PfK13 and PfMDR1 in samples from malaria patients presenting at health facilities in Fort Portal, from January to February 2024.
Occurrence of validated and associated PfK13 markers of artemisinin partial resistance in East Africa, 2020–2023, color-coded by majority markers. Colored circles indicate regions where validated or candidate PfK13 markers have been identified at prevalence >10% in 2020–2023. Yellow, majority of PfK13 mutations found are 675V and/or 469Y (2); purple, majority PfK13-561H (2, 10, 12, 13); blue, majority PfK13-441L (2); and brown, majority PfK13-469F (2). Background map created with mapchart.net.
Malaria patients attending four health facilities in the area of Fort Portal located approximately 100 km from the Eastern DRC border with Uganda were invited to participate in this descriptive study from January to February 2024, following informed written consent. The study protocol was approved by the Mbarara University of Science and Technology Research Ethics Committee. Malaria was diagnosed by available rapid diagnostic tests for malaria at the health centers, and finger-prick blood samples were collected on Whatman 3 mm paper. DNA was extracted using a commercial kit (Qiagen, Hilden, Germany). A single PCR for PfK13 spanning codons 426–709 (14) confirmed P. falciparum infection and generated amplicons for Sanger sequencing. Genotyping of PfMDR1 region 1 and region 2, including markers at codons 86, 184, and 1,246, was done by PCR and sequencing (15) on a random selection of P. falciparum-positive samples. Sequences were aligned to reference sequences from laboratory strain 3D7 (PlasmoDB.org), using CodonCode Aligner. In addition, a novel putative marker for lumefantrine tolerance, PfMDR1 Y500N (5), was typed by a high-resolution melting curve with a commercial SimpleProbe Assay (TIB MOLBiol, Berlin, Germany) on a Lightcycler 480 II (Roche). Positive and negative controls were included in all assays; mixed signals were counted as mutants.
Samples from 200 patients were analyzed, and 126 were confirmed by PfK13 PCR to contain P. falciparum. Validated or candidate PfK13 markers were present in 4.8% (6/126; 95% CI, 2.2%–10.0%). In addition, a PfK13 mutation of so far unknown relevance, 490T, was observed in almost one in ten isolates (Table 1). One health center differed by mutation pattern compared with the others, as two validated markers appeared exclusively there (Table 2). No clear geographic distinction or patient population was identified for this center. For PfMDR1, 102 samples were assessed, wildtype allele N86 was present in all samples, and mutation 184Y slightly dominated over the wildtype allele. A PfMDR1 mutation of unknown relevance, 199S, was present in more than 10%. The PfMDR1 Y500N mutation was absent. In a random 47 samples sequenced for the PfMDR1 region 2, wildtype allele D1246 prevalence was below 5% (Table 1).
Molecular surveillance data for AR in Uganda are relatively abundant, but not for the far western Fort Portal area, located between AR hotspots and eastern DR Congo (Fig. 1). We found diverse AR-associated PfK13 markers at a rather low combined prevalence of 4.8%. This contrasts with the situation in neighboring Rwanda and elsewhere in Uganda, where single AR markers reach 20%–40% (2, 10). PfK13 561H prevails in central Rwanda, 675V across Uganda (initially in the north), 441L in southern Uganda, and 469Y in northern Uganda (2). None of the markers dominates in Fort Portal, and 469Y is absent. As for lumefantrine, the PfMDR1 pattern, particularly the fixation of the wildtype allele N86, suggests increased tolerance (6–8). PfMDR1 500N, recently suggested as a novel marker for lumefantrine tolerance, was absent (5). The PfMDR1 mutation 199S (12.8%) is of unknown relevance, and it was previously seen at approximately 30% in northern Uganda in 2016/17, and at 2% in southern Rwanda in 2018/19 (16, 17). Its relevance needs to be evaluated in vitro or ex vivo. The same applies to a PfK13 polymorphism of unclear significance, 490T, which we detected at almost 10%. The prevalence of these variants of unknown significance might indicate positive selection, although temporal follow-ups are required to confirm this. The molecular signatures in the Fort Portal area suggest that AR is starting to appear alongside reduced lumefantrine susceptibility.
AR is a multifactorial phenomenon, and the genetic background of P. falciparum plays a major part in the level of resistance and fitness (18, 19). Fort Portal appears to be a melting pot of various P. falciparum strains from several regions, where different AR mutations prevail at high prevalence. In these hotspot regions, mutated strains have been successfully selected for beneficiary factors, such as survival advantage under drug pressure and fitness. The overlapping geographical occurrence of these strains in the Fort Portal area may facilitate an accelerated reassembling of genetic factors favoring AR. Multi-drug-resistant P. falciparum in Southeast Asia likely developed through the recombination of strains with each their own drug resistance genotype (20). In Uganda, increased lumefantrine tolerance has already been observed (5). Particularly in regions like the study area with low PfK13 mutation prevalence (bordering on so far hardly affected eastern DR Congo), the local selection of resistant parasite haplotypes should be rapidly monitored and targeted with scaled-up control measures. Facing a lack of alternatives to the ACTs and preventing the aggravation of AR in East Africa and its spread to other regions should be prioritized.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1World Health O. 2023. World malaria report 2023. World Health Organization, Geneva.
- 2Conrad MD, Asua V, Garg S, Giesbrecht D, Niaré K, Smith S, Namuganga JF, Katairo T, Legac J, Crudale RM, Tumwebaze PK, Nsobya SL, Cooper RA, Kamya MR, Dorsey G, Bailey JA, Rosenthal PJ. 2023. Evolution of partial resistance to artemisinins in malaria parasites in Uganda. N Engl J Med 389:722–732. doi:10.1056/NEJ Moa 221180337611122 PMC 10513755 · doi ↗ · pubmed ↗
- 3Phyo AP, Ashley EA, Anderson TJC, Bozdech Z, Carrara VI, Sriprawat K, Nair S, White MM, Dziekan J, Ling C, Proux S, Konghahong K, Jeeyapant A, Woodrow CJ, Imwong M, Mc Gready R, Lwin KM, Day NPJ, White NJ, Nosten F. 2016. Declining efficacy of artemisinin combination therapy against P. falciparum malaria on the Thai-Myanmar Border (2003-2013): the role of parasite genetic factors. Clin Infect Dis 63:784–791. doi:10.1093/cid/ciw 38827313266 PMC 4996140 · doi ↗ · pubmed ↗
- 4van der Pluijm RW, Tripura R, Hoglund RM, Pyae Phyo A, Lek D, Ul Islam A, Anvikar AR, Satpathi P, Satpathi S, Behera PK, et al.. 2020. Triple artemisinin-based combination therapies versus artemisinin-based combination therapies for uncomplicated Plasmodium falciparum malaria: a multicentre, open-label, randomised clinical trial. Lancet 395:1345–1360. doi:10.1016/S 0140-6736(20)30552-332171078 PMC 8204272 · doi ↗ · pubmed ↗
- 5Tumwebaze PK, Conrad MD, Okitwi M, Orena S, Byaruhanga O, Katairo T, Legac J, Garg S, Giesbrecht D, Smith SR, Ceja FG, Nsobya SL, Bailey JA, Cooper RA, Rosenthal PJ. 2022. Decreased susceptibility of Plasmodium falciparum to both dihydroartemisinin and lumefantrine in northern Uganda. Nat Commun 13:6353. doi:10.1038/s 41467-022-33873-x 36289202 PMC 9605985 · doi ↗ · pubmed ↗
- 6Veiga MI, Dhingra SK, Henrich PP, Straimer J, Gnädig N, Uhlemann AC, Martin RE, Lehane AM, Fidock DA. 2016. Globally prevalent Pf MDR 1 mutations modulate Plasmodium falciparum susceptibility to artemisinin-based combination therapies. Nat Commun 7:11553. doi:10.1038/ncomms 1155327189525 PMC 4873939 · doi ↗ · pubmed ↗
- 7Okell LC, Reiter LM, Ebbe LS, Baraka V, Bisanzio D, Watson OJ, Bennett A, Verity R, Gething P, Roper C, Alifrangis M. 2018. Emerging implications of policies on malaria treatment: genetic changes in the Pfmdr-1 gene affecting susceptibility to artemether-lumefantrine and artesunate-amodiaquine in Africa. BMJ Glob Health 3:e 000999. doi:10.1136/bmjgh-2018-000999 PMC 620299830397515 · doi ↗ · pubmed ↗
- 8Venkatesan M, Gadalla NB, Stepniewska K, Dahal P, Nsanzabana C, Moriera C, Price RN, Mårtensson A, Rosenthal PJ, Dorsey G, et al.. 2014. Polymorphisms in Plasmodium falciparum chloroquine resistance transporter and multidrug resistance 1 genes: parasite risk factors that affect treatment outcomes for P. falciparum malaria after artemether-lumefantrine and artesunate-amodiaquine. Am J Trop Med Hyg 91:833–843. doi:10.4269/ajtmh.14-003125048375 PMC 4183414 · doi ↗ · pubmed ↗