Prevalence and characteristics of tigecycline- and carbapenem-resistant adeN-truncated Acinetobacter baumannii: a genomic epidemiological analysis
Ying Zhang, Beibei Zhou, Jingchun Kong, Panjie Hu, Haifeng Liu, Deyi Zhao, Jianzhong Ye, Qingxia Fu, Tieli Zhou, Changrui Qian

TL;DR
This study investigates the spread and characteristics of tigecycline- and carbapenem-resistant Acinetobacter baumannii with a specific genetic mutation in a hospital setting.
Contribution
The study provides new insights into the genomic epidemiology and clinical impact of adeN-truncated Acinetobacter baumannii isolates.
Findings
AdeN-truncated Acinetobacter baumannii isolates showed high prevalence and resistance to tigecycline and carbapenems.
These isolates were associated with higher mortality rates and specific risk factors like ICU admission and immunosuppressant use.
Genomic analysis revealed clonal transmission and limited capsular diversity in adeN-truncated isolates.
Abstract
adeN-truncated Acinetobacter baumannii (ATAB) isolates are associated with elevated tigecycline resistance and enhanced virulence, yet its epidemic dynamics and genomic features remain poorly understood. This study aimed to investigate the epidemiology of ATAB isolates, identify infection risk factors, and assess their impact on patient prognosis. The prevalence of ATAB isolates in a tertiary care teaching hospital (Wenzhou, China) from January 2018 to December 2022 was determined via polymerase chain reaction (PCR) screening. Whole-genome sequencing and genomic analysis were conducted to explore the epidemiology and genomic characteristics of 254 ATAB isolates. Clinical data analysis was performed to identify risk factors for ATAB infection and its correlation with patient prognosis. The results of local sample analysis showed that adeN truncation was identified in 26.5% (486/1834) of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
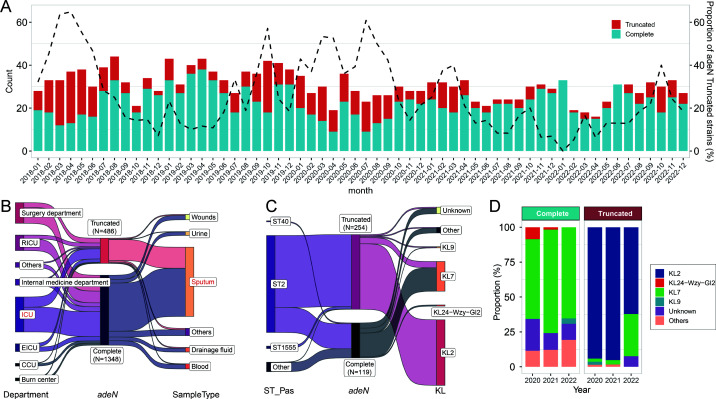 Fig 1
Fig 1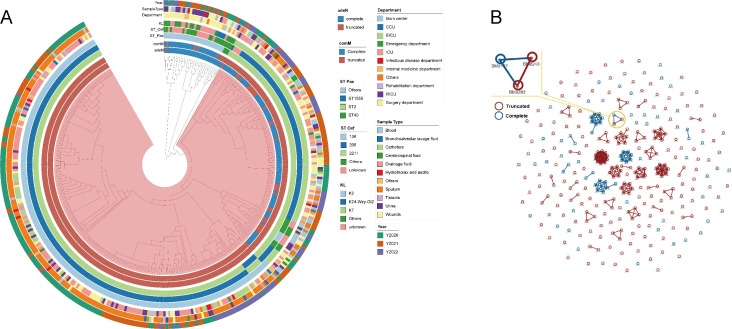 Fig 2
Fig 2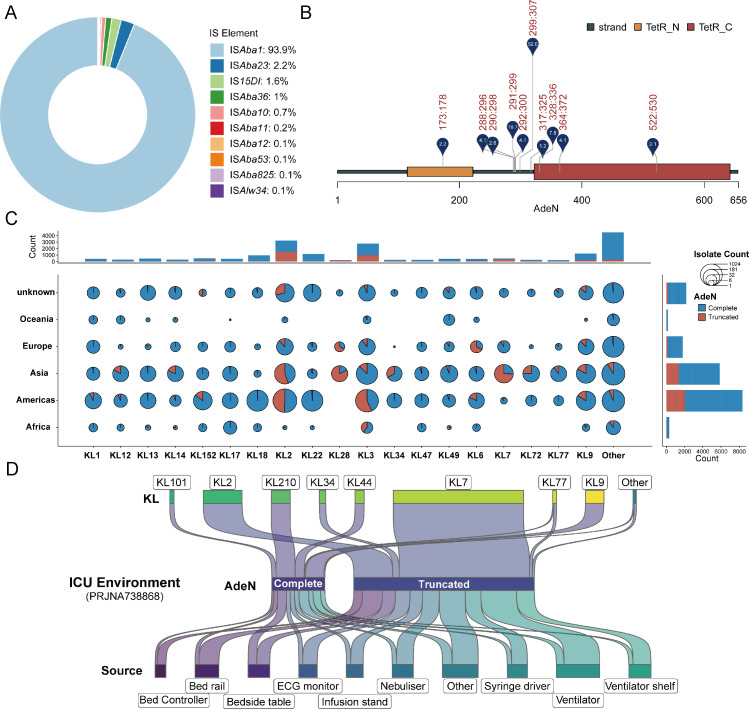 Fig 3
Fig 3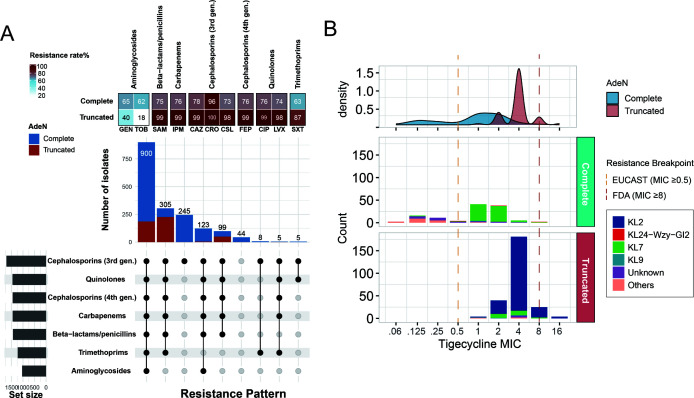 Fig 4
Fig 4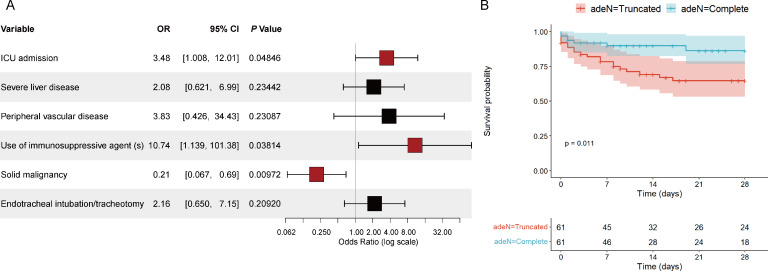 Fig 5
Fig 5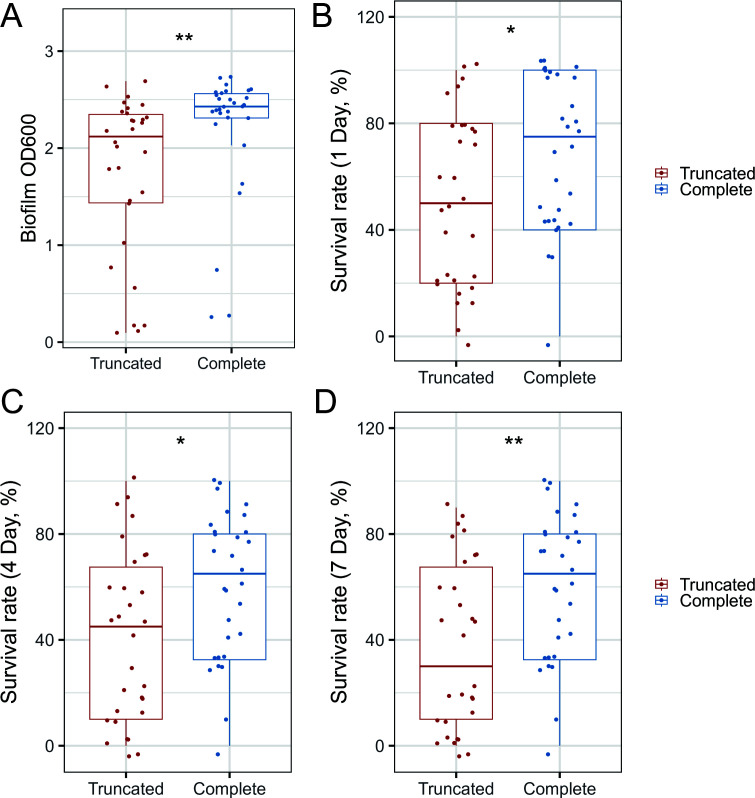 Fig 6
Fig 6- —Key Laboratory of Clinical Laboratory Diagnosis and Translational Research of Zhejiang Province
- —Zhejiang Province Medical and Health Technology Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Antibiotics Pharmacokinetics and Efficacy · Antibiotic Use and Resistance
INTRODUCTION
Acinetobacter baumannii is a major nosocomial pathogen, frequently causing life-threatening infections such as pneumonia, meningitis, and bloodstream infections (1). Its ability to develop resistance to multiple antimicrobial agents, particularly carbapenems, has made treatment options increasingly limited (2, 3). Tigecycline is often regarded as one of the last lines of defense against carbapenem-resistant A. baumannii (CRAB). However, the emergence of tigecycline-resistant isolates has been widely reported, primarily driven by the overexpression of RND efflux pumps, such as AdeABC and AdeIJK, which can reduce tigecycline susceptibility to varying degrees (4–7).
AdeN, a TetR family transcriptional regulator, critically regulates the expression of AdeIJK, the most conserved RND efflux pump (8). In 2017, Rajagopalan et al. first reported an ISAba1 insertion in the adeN gene of clinical A. baumannii isolates, demonstrating that this genetic alteration increases the lethality of A. baumannii ATCC 17978 against A549 lung cells (9). ISAba1-mediated truncation of adeN has also been identified in multiple studies as a key resistance-associated genetic alteration, linked to tigecycline resistance (5–7). Despite these insights, the epidemiological characteristics of adeN-truncated A. baumannii (ATAB) isolates are not yet well understood. Moreover, the prognostic outcomes of patients infected with ATAB isolates, compared to those with adeN-complete isolates, remain to be further elucidated. To address these gaps, this study retrospectively screened A. baumannii isolates collected over a 5-year period from a large tertiary care hospital for adeN truncations. Through whole-genome sequencing and genomic database mining, we systematically investigated the epidemiological characteristics of ATAB isolates and analyzed potential infection risk factors and clinical outcomes using comprehensive clinical data.
RESULTS
ATAB isolates are prevalent in hospital patients
A total of 1,834 A. baumannii isolates from patients were collected between January 2018 and December 2022 from a large tertiary care hospital to investigate the prevalence of ATAB isolates. These isolates were derived from various clinical specimens collected from patients, including sputum (72.8%, 1,336/1,834), wounds (5.5%, 101/1,834), and blood (5.3%, 97/1,834) (Fig. S1). The polymerase chain reaction (PCR) screening result showed that 26.5% (486/1,834) of A. baumannii isolates harbored truncated adeN. The proportion of ATAB isolates showed considerable fluctuations over the years, peaking at 64.9% (24/37) in April 2018 (Fig. 1A). The ATAB isolates were primarily derived from sputum specimens of various ICU patients (Fig. 1B).
Epidemiological characteristics and distribution of adeN-truncated and adeN-complete A. baumannii isolates in the hospital. (A) Monthly distribution of adeN-truncated and adeN-complete isolates over time. (B) Distribution of adeN-truncated and adeN-complete isolates in different hospital departments and sample types. (C) Molecular typing of isolates with different adeN statuses. (D) Temporal distribution of KL types of adeN-truncated and adeN-complete isolates.
WGS was performed on all ATAB isolates (N = 254) from 2020 to 2022, alongside randomly selected adeN-complete strains (N = 119) from the same period. The metadata for these strains have been provided as the supplementary table (Table S1). These ATAB isolates predominantly belonged to the IC2/ST2 clone (96.5%, 245/254), and the main capsule type was KL2 (87.8%, 223/254) (Fig. 1C). In contrast, adeN-complete strains exhibited greater diversity in MLST and capsule types, with ST2-KL7 being most common (66.4%, 79/119) and no KL2 clones identified. By analyzing the capsule-type distribution over time, we found that adeN-complete KL7 strains maintained a consistent prevalence, whereas adeN-truncated KL7 strains expanded significantly in 2022 (Fig. 1D).
Frequent clonal dissemination of ATAB isolates in nosocomial settings
To investigate the relationships between ATAB and adeN-complete isolates, we constructed a cgSNP-based phylogenetic tree (Fig. 2A). We found that the ST2^Pas^/ST208^Oxf^-KL2 lineage formed the largest ATAB branch, widely distributed across multiple departments. The adeN-complete isolates were mainly composed of the ST2^Pas^/ST208^Oxf^-KL7 lineage. We clustered all ST2^Pas^/ST208^Oxf^-KL2 and ST2^Pas^/ST208^Oxf^-KL7 isolates using a cgSNP distance of 0 to search for potential clonal transmission (Fig. 2B). Interestingly, we found that ATAB isolates formed 17 clusters, involving up to 35.2% (83/236) of the isolates, while adeN-complete isolates formed only three genetic clusters, involving 30.7% (23/75) of the isolates. This indicated ATAB isolates have driven significant clonal transmission in the hospital. Notably, within one cluster, adeN-complete isolates BM8137 evolved to ATAB BM8243 and BM8283, suggesting adeN truncation occurs in the nosocomial environment.
Phylogenetic analysis of 373 sequenced A. baumannii isolates (A) Maximum-likelihood cgSNP-based phylogenetic tree. The rings from inside to outside represent the isolate’s adeN status, comM status, sequence type of the Pasteur scheme, sequence type of the Oxford scheme, K type, department, sample type, and year, respectively. (B) Genetic clusters based on cgSNPs with an SNP distance of 0. Red circles represent ATAB isolates, blue circles represent adeN-complete isolates, and the connecting line represents that the cgSNP distance between isolates is 0.
Global emergence and prevalence of ATAB isolates
To investigate the global prevalence of ATAB isolates, we analyzed 18,946 high-quality A. baumannii genomes from the GenBank database. Our results revealed that these ATAB isolates were widespread across numerous regions (Fig. S2). ISAba1 was the dominant IS element causing truncation (93.9%), followed by ISAba23 (2.2%) (Fig. 3A). Notably, 52.6% of insertions occurred between positions 299 and 307 of the adeN gene (Fig. 3B). Consistent with local observations, the IC2 clone was the predominant genotype of ATAB isolates across continents (Fig. S3). Among the five major IC2-associated capsule types (KL2, KL3, KL7, KL9, and KL22), KL22 primarily comprised adeN-complete strains, while the others showed varying degrees of adeN truncation (Fig. S4). Notably, adeN-truncated KL2 strains were widespread globally, KL3 strains were predominant in the Americas, and KL7 strains were mainly found in Asia (Fig. 3C). Interestingly, ATAB strains were first identified in Europe in 2003 and have since spread widely across America and Asia (Fig. S5). These findings underscore the global prevalence of ATAB isolates and their strong association with specific genetic traits.
Global distribution and genomic characteristics of adeN-truncated and adeN-complete A. baumannii isolates. (A) Prevalence of different IS elements associated with truncation of the adeN gene. (B) Schematic of the adeN gene with truncation sites caused by IS elements. Only sites with a frequency higher than 2% are shown. The numbers next to the markers indicate the proportion of strains with truncations at each site. (C) Distribution of KL types among adeN-truncated and adeN-complete isolates across different continents. Bubble sizes represent the number of strains, with pie chart segments indicating the proportions of truncated versus complete adeN isolates. (D) Source distribution of adeN-truncated and adeN-complete isolates in the ICU environment (PRJNA738868).
Importantly, we expanded our analysis to include a recently published large-scale genomic data set from another hospital in Zhejiang Province. This data set comprised 432 strains isolated from various medical device surfaces in the ICU ward between August and October 2019 (10). Surprisingly, we found that ATAB isolates were highly prevalent (73%, 334/432) across all types of surfaces in the ICU (Fig. 3D). The capsule type of these ATAB isolates was predominantly KL7 (72.2%, 241/334), followed by KL2 (21.6%, 72/334). This finding highlights the widespread presence of ATAB isolates in ICU environments.
High correlation of ATAB isolates with carbapenem resistance
We compared the drug resistance profiles of 1,765 isolates with complete susceptibility data for 11 antibiotics (Fig. 4A). ATAB isolates showed significantly higher resistance to cephalosporins, carbapenems, and quinolones, with rates approaching or exceeding 99% (P < 0.001). However, ATAB isolates had lower resistance to aminoglycosides (P < 0.001). Further analysis of resistance patterns revealed that 65% of ATAB isolates exhibited multidrug-resistant phenotypes, while 35% were extensively drug-resistant. Carbapenem resistance in A. baumannii is primarily mediated by carbapenem-hydrolyzing class D β-lactamases, such as blaOXA-23, which are typically located within AbaR-type resistance islands (AbaRs) (11, 12). Previous studies have demonstrated that AbaRs are predominantly inserted into the comM gene, which encodes a helicase involved in DNA recombination during natural transformation (13, 14). Interestingly, we observed that most of the ATAB isolates in both local and global data sets also exhibited truncations in the comM gene (Fig. 2A; Fig. S6). This suggested that adeN truncation predominantly occurs in strains where the AbaR island has integrated into comM. Overall, these findings highlight a strong association between ATAB and antimicrobial resistance, emphasizing the urgent need for targeted interventions in clinical settings.
Antimicrobial susceptibility phenotypes of adeN-truncated and adeN-complete A. baumannii isolates. (A) Resistance rates and pattern of ATAB and adeN-complete isolates across different antimicrobials. The heatmap at the top indicates the resistance percentage, while the bar plot in the middle shows the number of isolates resistant to each antibiotic. The bottom section presents an UpSet plot of resistance patterns across different antibiotic classes. (B) Distribution of tigecycline MICs for adeN-truncated and adeN-complete isolates. The density plot at the top illustrates the distribution of MIC values, while the bar plot below shows the count of isolates. The vertical lines represent the tigecycline resistance breakpoints according to EUCAST and FDA guidelines.
Genetic background affects tigecycline resistance of ATAB isolates
Given the association between adeN truncation and reduced tigecycline susceptibility, we further analyzed tigecycline MICs in 373 sequenced isolates. The results showed that tigecycline MICs for ATAB isolates were generally high, ranging from 1 to 16 mg/L, with an MIC_50_ value of 4 mg/L (Fig. 4B). Based on EUCAST breakpoints, all ATAB isolates were tigecycline-resistant, whereas FDA standards classified only 11.4% (29/254) ATAB isolates as resistant. In contrast, tigecycline MICs for adeN-complete isolates were more widely distributed, ranging from 0.06 to 8 mg/L, and were correlated with the capsular types of isolates. Notably, adeN-complete KL7 strains exhibited relatively high MIC values, with an MIC_50_ of 2 mg/L, approaching that of adeN-truncated KL7 strains (MIC_50_ of 4 mg/L). Other capsular-type strains displayed lower tigecycline MIC values, with an MIC_50_ of 0.125 mg/L. These results update our understanding of the association between adeN truncation and tigecycline resistance phenotype.
Infection of ATAB isolates is associated with worse patient outcomes
To evaluate the specific impact of infection with ATAB isolates on patient clinical outcomes, this study included 61 patients from whom ATAB isolates were isolated from sterile body fluids. As a control, 61 patients with adeN-complete strains isolated were randomly selected through case-matching. There were no significant differences between the two groups in terms of gender, age distribution, and Charlson Comorbidity Index (Table S2). Notably, we observed that patients in the ATAB group had a significantly longer pre-infection hospital stay (median 42 days vs 27 days, P < 0.05) and higher ICU admission rate (75.4% vs 39.3%, P < 0.001). Additionally, the ATAB group was more likely to suffer from severe liver disease (29.5% vs 11.5%, P < 0.05), peripheral vascular disease (16.4% vs 1.6%, P < 0.01), immunosuppressant use (16.4% vs 1.6%, P < 0.01), and receiving tracheal intubation or tracheostomy (73.7% vs 44.3%, P < 0.001). In contrast, the incidence of solid malignancies was higher in the adeN-complete group (31.1% vs 11.5%, P<0.01) (Table S2). Further logistic regression analysis of these significant factors identified ICU admission and immunosuppressant use as independent risk factors for infection with ATAB isolates (Fig. 5A). Patients infected with ATAB exhibited higher 14-day (29.5%, 18/61) and 28-day (32.8%, 20/61) mortality rates compared to those infected with adeN-complete A. baumannii (P < 0.01) (Table S2). Furthermore, the Kaplan–Meier curves with the log-rank test revealed a significant difference in 28-day survival between the two groups (P < 0.05) (Fig. 5B).
Clinical data analysis of patients infected with adeN-truncated and adeN-complete isolates. (A) Forest plot of risk factors associated with infection by adeN-truncated A. baumannii isolates. Odds ratios (OR) with 95% confidence intervals (CI) are displayed on a logarithmic scale. The red squares indicate variables with significant P-values (P < 0.05). (B) Kaplan–Meier survival curves in patients with A. baumannii isolate infection.
ATAB has higher virulence and reduced biofilm formation ability
To assess the virulence characteristics of patient-isolated adeN-truncated strains, we randomly selected 30 adeN-truncated and 30 adeN-complete A. baumannii clinical isolates for subsequent experiments. Biofilm formation was compared between the two groups using crystal violet staining, revealing that adeN-truncated A. baumannii strains exhibited significantly weaker biofilm formation ability compared to the adeN-complete strains (Fig. 6A). Furthermore, we assessed the in vivo pathogenicity of the two group isolates using the G. mellonella larvae infection model. The results demonstrated that the 1-day, 4-day, and 7-day survival rates of G. mellonella larvae infected with the adeN-truncated strains were lower compared to those infected with the adeN-complete strains (Fig. 6B through D).
Comparison of virulence traits between the adeN-truncated and adeN-complete isolates. (A) Biofilm formation. (B–D) Survival plot of Galleria mellonella larvae infected with the adeN-truncated group compared to the adeN-complete group. (B) Observed for 1 day. (C) Observed for 4 days. (D) Observed for 7 days.
DISCUSSION
This study comprehensively explored the genomic epidemiological characteristics of adeN-truncated A. baumannii at both local and global levels. Our results reveal a widespread prevalence of adeN-truncated A. baumannii in hospital settings. This strain often exhibits more severe antimicrobial resistance characteristics and is closely associated with poor prognoses in infected patients. Our findings emphasize the potential value of adeN truncation in guiding clinical treatment and prognosis assessment for infected patients. Furthermore, our study highlights a close association between adeN-truncated A. baumannii and a specific global epidemic lineage, which enriches our understanding of the epidemic dynamics of adeN-truncated A. baumannii.
The IC2 lineage of A. baumannii is a globally successful, carbapenem-resistant clonal group that has caused numerous outbreaks of hospital-acquired infections worldwide (15–17). Our study demonstrated that adeN truncation predominantly occurs in the IC2 clonal group, with KL2 and KL3 being the primary capsular types in the Americas and KL2 and KL7 in Asia. Notably, we also observed the expansion of KL7-type ATAB strains, which is consistent with our previous studies (18). Several studies have shown that KL7 has a higher virulence than other capsule types and has caused outbreaks in multiple hospitals, suggesting that it has a good fitness advantage in hospitals (19, 20). Although the driving factors behind ATAB isolate dissemination remain unclear, previous studies have shown that disinfectants significantly increase the frequency of adeN inactivation in A. baumannii (21, 22). Taking these factors into consideration, we speculate that the differences in genotypes (such as capsule types) of ATAB strains may be related to the frequent exposure of local hospital epidemic lineages to disinfectant-induced selection pressure. Based on the phylogenetic analysis, we also observed that ATAB evolved within the hospital and led to multiple clonal transmissions, which may also be the driving factor for its expansion. Notably, adeN truncation may impose an adaptive burden on A. baumannii due to overexpression of the AdeIJK efflux pump, potentially limiting the competitive advantage of ATAB isolates in environments without selective pressures (23, 24). These multifaceted factors likely contribute to the periodic prevalence of ATAB isolates in hospital settings.
We also observed an intriguing association between ATAB isolates and specific resistance patterns, particularly resistance to cephalosporins, carbapenems, fluoroquinolones, and tigecycline. Although AdeIJK efflux pump substrates include β-lactams, fluoroquinolones, and tetracyclines, its contribution to resistance appears limited (21, 23). We hypothesize that genomic differences in resistance determinants within ATAB lineages also play a significant role. Notably, for tigecycline resistance, we found that the adeN-complete KL7 lineage itself has a high MIC value for tigecycline (MIC_50_ = 1). Studies have demonstrated that high-level tigecycline resistance in A. baumannii is frequently associated with the overexpression of the AdeIJK efflux pump or the presence of plasmid-borne tigecycline-inactivating enzyme TetX variants (5, 25). Furthermore, specific mutations within chromosomal genes, such as trm and plsC, may contribute to diminished tigecycline susceptibility (26, 27). We speculate that the accumulation of tigecycline resistance-associated mutations in the KL7-type A. baumannii isolates resulted in reduced sensitivity to tigecycline. Therefore, when adeN truncation occurs, it leads to clinical-level resistance in strains. For other lineages, however, the occurrence of adeN truncation may not be sufficient to induce clinical resistance to tigecycline. This underscores the importance of considering the genetic background of the strains when interpreting the relationship between adeN truncation and clinical tigecycline resistance.
A. baumannii primarily infects immunocompromised hospitalized patients, with infection-related mortality rates ranging from 34% to 44.5% (28). Previous studies have shown that the mortality rate of patients infected with A. baumannii is closely related to the severity of infection, antimicrobial resistance, timeliness of treatment, and the lineage of the infecting strain (29, 30). In the present study, a statistically significant elevation in mortality risk was observed among patients infected with ATAB isolates. Consistent with previous reports, we demonstrated enhanced lethality of ATAB strains in the Galleria mellonella larval infection model (9). Considering the multifaceted characteristics of ATAB strains, we hypothesize that the increased mortality risk may be attributed to a confluence of factors, including (i) virulence enhancement due to adeN gene truncation; (ii) complex antimicrobial resistance profiles, potentially contributing to clinical treatment failure; and (iii) intrinsic pathogenic properties associated with the prevalent ATAB lineage. Collectively, these findings underscore the potential clinical utility of adeN truncation as an early indicator for guiding therapeutic interventions and prognostic assessments in A. baumannii infections.
This study has several limitations. First, as a single-center analysis, the generalizability of conclusions regarding the genetic characteristics and prevalence of ATAB isolates may be limited. Second, the small number of isolates from sterile sites constrained our ability to fully assess the potential impact of factors beyond the adeN status on patient outcomes. Furthermore, the inherent challenge of differentiating between true infection and colonization represents a notable limitation. While our study included only patients with isolates from sterile body fluids, the diagnosis of pneumonia was often based on clinical criteria and microbiological findings from nonsterile sites such as sputum or bronchoalveolar lavage. Furthermore, selection bias and the number of adeN-complete isolates used for WGS may limit the validity of some comparisons.
In conclusion, this study highlights the widespread presence of ATAB isolates as a potential high-risk clone. Given its association with resistance and virulence, early identification and monitoring of ATAB as a marker for hospital infection control are crucial for developing effective management and treatment strategies. Furthermore, the role of environmental disinfectants in driving the emergence and spread of ATAB warrants careful consideration.
MATERIALS AND METHODS
Collection and screening of adeN-truncated isolates
This retrospective study was conducted at the First Affiliated Hospital of Wenzhou Medical University, a large tertiary care teaching hospital in Zhejiang, China. A. baumannii isolates collected from January 2018 to December 2022 were used as experimental strains. The isolates were identified using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF/MS; bioMérieux, Lyon, France). These isolates were preserved in 30% glycerol broth vials and stored at −80°C for future use. The integrity of the adeN gene in A. baumannii isolates was assessed using PCR. An adeN truncation was indicated if the PCR product band size after gel electrophoresis was 800 bp larger than that of A. baumannii ATCC 17978 (primer sequences are detailed in Table S3).
Antimicrobial susceptibility testing
The susceptibility of strains to clinically used antibiotics, including amikacin, ceftazidime, ciprofloxacin, ceftriaxone, cefepime, gentamicin, imipenem, levofloxacin, ampicillin/sulbactam, cotrimoxazole and tobramycin, was determined using the automated broth microdilution method with VITEK2 (bioMérieux, Marcy-l'Étoile, France). Tigecycline susceptibility was determined using the manual broth microdilution method. Results for all other antimicrobial agents were interpreted according to the latest CLSI guidelines, while tigecycline susceptibility was interpreted based on EUCAST and FDA guidelines.
Whole-genome sequencing, genome assembly, and annotation
Genomic DNA from 373 A. baumannii isolates, including both adeN-truncated and adeN-complete strains, was extracted using the AxyPrep Bacterial Genomic DNA Miniprep Kit (Axygen Scientific, USA). The DNA library was prepared using the NEBNext Ultra II DNA library preparation kit and subsequently sequenced by the Illumina NovaSeq. The quality control of raw sequence reads was performed using FastQC (https://github.com/s-andrews/FastQC) and trimmed using fastp with default parameters (31). The trimmed reads for each strain were assembled using SPAdes, and draft assemblies were annotated with Prokka (32, 33). The multi-locus sequence type (MLST) analysis of A. baumannii was performed using pymlst (34). The capsular (K-locus) and O-antigen (O-type) genotypes were determined using Kaptive (35). The antimicrobial resistance genes (ARGs) on A. baumannii were identified using AMRFinderPlus (36). The Insertion Sequence (IS) elements inserted into the adeN gene were identified using ISfinder (37).
Phylogenetic analysis
The core genome alignment of genomes was identified using Snippy (https://github.com/tseemann/snippy), with the complete genome of BM2333, an IC2/ST2 strain previously isolated in our hospital, serving as the reference for comparison. The recombination site of generated alignment was removed using Gubbins (38). SNP distances were calculated from the Gubbins-filtered polymorphic sites file using SNP-dists (https://github.com/tseemann/snp-dists). Clonal transmission clusters with a core-genome SNP (cgSNP) distance of 0 were extracted using the R library iGRAPH (39).
Analysis of A. baumannii assemblies from GenBank
A total of 18,946 completed and draft A. baumannii genome assemblies were downloaded from GenBank as of Jun 2023. The AST phenotype data of A. baumannii were retrieved from NCBI Pathogen Detection (www.ncbi.nlm.nih.gov/pathogens/). The completeness of the adeN gene in the A. baumannii genomes was checked using a self-written Python script (https://github.com/qcrcherry/comparative_genomic_analysis).
Clinical data collection and definitions
To minimize errors caused by colonization, the study focused on adult patients (aged 18 and above) with the ATAB strain isolated from sterile body fluids (e.g., blood, pleural or peritoneal fluid, and cerebrospinal fluid). These patients formed the study group, while patients with adeN-complete strains from the same sample types were randomly selected as controls. Outpatient and duplicate cases were excluded. The Charlson Comorbidity Index was used to adjust for underlying diseases, and all-cause in-hospital mortality was recorded. The infection date was defined as the first isolation of A. baumannii from sterile body fluids. Infection sites were classified according to CDC guidelines, and laboratory results from the day of strain isolation were recorded. The primary outcome was the all-cause mortality rate within 28 days after A. baumannii infection.
Biofilm formation assay
The biofilm-forming ability of A. baumannii was assessed using the crystal violet staining method. Bacterial suspensions were first adjusted to a concentration of 1.5 × 10^8^ CFU/mL. The suspensions were then diluted 100-fold with the LB medium and added to wells of a polystyrene 96-well plate (Corning, USA). After 24 hours of static incubation, the wells were stained with crystal violet (Solarbio, China), and excess stain was removed by washing with phosphate-buffered saline (PBS), followed by washing with absolute ethanol. The absorbance at 600 nm was measured to determine the average optical density of the triplicates. The experiment was repeated three times to obtain the mean value.
In vivo G. mellonella survival rate analysis
The virulence of A. baumannii was assessed using an in vivo G. mellonella infection model (40). G. mellonella larvae weighing between 250 and 300 mg were used for the infection experiment. PBS served as the negative control. Overnight bacterial cultures were washed with PBS and resuspended to an OD_600_ of 0.5. A 10 µL aliquot of a tenfold dilution of each bacterial strain was injected into the left forelimb of each larva, with 10 larvae per group. The larvae were maintained at 37°C, and their survival was monitored continuously for 7 days post-injection. Survival rates were recorded at the corresponding time points.
Statistical analysis and visualization
Data processing and statistical analyses were conducted using R v4.2. Categorical variables were reported as numbers (percentages) and compared using χ or Fisher’s exact tests, while continuous variables were presented as mean ± standard deviation or median (interquartile range) and analyzed using t-tests or Mann–Whitney U tests. A significance level of P < 0.05 was applied. A world map was built using the R package “rworldmap.” The Sankey diagram was produced by “ggsankey.” Survival analysis was performed by “survival.” The Kaplan–Meier curve was generated using “survminer.” The UpSet plot was generated using “ComplexUpset” v1.4.0. Univariate and multivariate logistic regression analyses were performed to analyze the risk factors for infection with the ATAB isolates, and the results were visualized using the “forestplot.”
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dijkshoorn L, Nemec A, Seifert H. 2007. An increasing threat in hospitals: multidrug-resistant Acinetobacter baumannii. Nat Rev Microbiol 5:939–951. doi:10.1038/nrmicro 178918007677 · doi ↗ · pubmed ↗
- 2Perez F, Hujer AM, Hujer KM, Decker BK, Rather PN, Bonomo RA. 2007. Global challenge of multidrug-resistant Acinetobacter baumannii. Antimicrob Agents Chemother 51:3471–3484. doi:10.1128/AAC.01464-0617646423 PMC 2043292 · doi ↗ · pubmed ↗
- 3Nhu NTK, Lan NPH, Campbell JI, Parry CM, Thompson C, Tuyen HT, Hoang NVM, Tam PTT, Le VM, Nga TVT, Nhu TDH, Van Minh P, Nga NTT, Thuy CT, Dung LT, Yen NTT, Van Hao N, Loan HT, Yen LM, Nghia HDT, Hien TT, Thwaites L, Thwaites G, Chau NVV, Baker S. 2014. Emergence of carbapenem-resistant Acinetobacter baumannii as the major cause of ventilator-associated pneumonia in intensive care unit patients at an infectious disease hospital in southern Vietnam. J Med Microbiol 63:1386–1394. doi:10.1099/jmm.0. · doi ↗ · pubmed ↗
- 4Isler B, Doi Y, Bonomo RA, Paterson DL. 2019. New treatment options against carbapenem-resistant Acinetobacter baumannii infections. Antimicrob Agents Chemother 63:10–1128. doi:10.1128/AAC.01110-18PMC 632523730323035 · doi ↗ · pubmed ↗
- 5Lucaßen K, Müller C, Wille J, Xanthopoulou K, Hackel M, Seifert H, Higgins PG. 2021. Prevalence of RND efflux pump regulator variants associated with tigecycline resistance in carbapenem-resistant Acinetobacter baumannii from a worldwide survey. J Antimicrob Chemother 76:1724–1730. doi:10.1093/jac/dkab 07933760099 · doi ↗ · pubmed ↗
- 6Jo J, Ko KS. 2021. Tigecycline heteroresistance and resistance mechanism in clinical isolates of Acinetobacter baumannii. Microbiol Spectr 9:e 0101021. doi:10.1128/Spectrum.01010-2134523993 PMC 8557860 · doi ↗ · pubmed ↗
- 7Gerson S, Nowak J, Zander E, Ertel J, Wen Y, Krut O, Seifert H, Higgins PG. 2018. Diversity of mutations in regulatory genes of resistance-nodulation-cell division efflux pumps in association with tigecycline resistance in Acinetobacter baumannii. J Antimicrob Chemother 73:1501–1508. doi:10.1093/jac/dky 08329554339 · doi ↗ · pubmed ↗
- 8Rosenfeld N, Bouchier C, Courvalin P, Périchon B. 2012. Expression of the resistance-nodulation-cell division pump Ade IJK in Acinetobacter baumannii is regulated by Ade N, a Tet R-type regulator. Antimicrob Agents Chemother 56:2504–2510. doi:10.1128/AAC.06422-1122371895 PMC 3346617 · doi ↗ · pubmed ↗