In vitro molting of Dirofilaria immitis third-stage larvae derived from microfilariae collected from doxycycline-treated dogs
Yi Chu, Elyssa Campbell, Michael Dzimianski, Christopher C. Evans, Cassan Pulaski, Kaori Sakamoto, Andrew R. Moorhead

TL;DR
This study examines how doxycycline affects the development of heartworm larvae, finding that it reduces Wolbachia levels but does not prevent larval molting.
Contribution
The study identifies that doxycycline reduces Wolbachia in heartworm larvae without affecting their ability to molt.
Findings
Doxycycline treatment significantly reduces Wolbachia levels in microfilariae and L3 larvae.
Larval molting from L3 to L4 is not impacted by doxycycline-induced Wolbachia reduction.
Wolbachia levels remain below five percent of controls throughout doxycycline treatment.
Abstract
Dirofilaria immitis, also known as canine heartworm, contains an endosymbiont, Wolbachia, in all life stages. The antibiotic, doxycycline, has been incorporated into heartworm treatment protocols to eliminate Wolbachia. Previous studies indicate that subsequent infection cannot be established using viable third-stage larvae (L3) developed from doxycycline-treated microfilariae (mf). The stages in which the development of larvae is impacted by doxycycline remain unknown. We examined the impact of doxycycline on the third-stage to fourth-stage larval molt, as it is the first molt of D. immitis after it invades the vertebrate host. Microfilaremic blood was collected weekly from D. immitis–infected dogs with or without doxycycline treatment at 10 mg/kg as recommended by the American Heartworm Society. Blood was collected weekly until the end of doxycycline treatment. The blood was used for…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1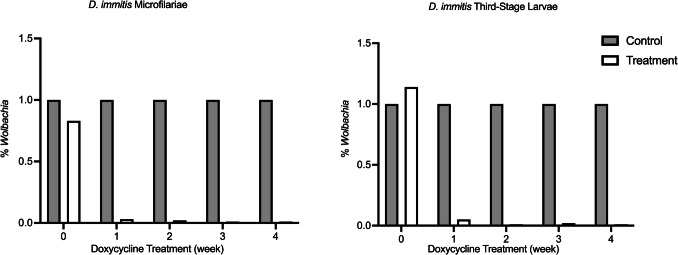 Figure 2
Figure 2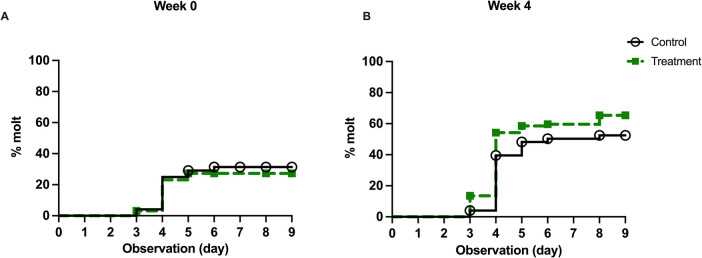 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasitic Diseases Research and Treatment · Insect symbiosis and bacterial influences
Background
Heartworm disease (HWD), caused by the parasitic nematode Dirofilaria immitis, affects multiple animal species worldwide. As of 2023, HWD has been diagnosed in all 50 states of the United States of America (USA) (American-Heartworm-Society 2023). Infected dogs harboring circulating microfilariae (mf) can be reservoirs for subsequent infection, and more than 30 species of animals, including humans, can be infected by this parasite (Nelson 2023). Incidence rates of HWD in the southeastern USA indicated an overall increase of 21.7% with minimal change in the proportion of dogs receiving heartworm prevention between the years 2013 and 2016 (Drake & Wiseman 2018). The increase in heartworm incidence rates demands additional efforts control and treat the disease.
Dirofilaria immitis is a filarial parasite related to the human pathogens Onchocerca volvulus, Brugia malayi, and Wuchereria bancrofti. Dirofilaria immitis requires a vertebrate host and an invertebrate host to complete its life cycle. A mosquito takes a blood meal from an infected host containing circulating mf; the ingested mf then develop into third-stage infective larvae (L3) in the mosquito. When the mosquito takes another blood meal from a susceptible host, the L3 migrate to the mouth parts of the mosquito and enter the host through the bite wound created by the feeding activity (Abraham 1988; McCall et al. 2008). In the subsequent host, the L3 develop and molt into fourth-stage larvae (L4) three to 12 days after infection. The L4 eventually molt into immature adults and are present in the pulmonary artery by 120 days post-infection, where the adult male and female worms mate and produce mf (Abraham 1988; Kotani & Powers 1982; McCall et al. 2008). Transmission and the life cycle continue when another mosquito takes a blood meal from the infected host.
The American Heartworm Society (AHS) recommends using three doses of melarsomine dihydrochloride via intramuscular injection as the adulticidal treatment of HWD. Prior to the first dose of melarsomine, AHS recommends a 28-day course of doxycycline, a bacteriostatic agent belonging to the tetracycline family, at 10 mg/kg BID. Doxycycline can effectively reduce the bacterial endosymbiont Wolbachia in the parasite and subsequently reduce pathology and complications that may occur in the host following adulticidal treatment (Foster et al. 2005; Kramer et al. 2008; Nelson et al. 2017). Doxycycline has been shown to be effective against early infection of D. immitis in dogs by McCall et al. (2011) in a Brugia pahangi and D. immitis co-infection model. Experimental dogs were inoculated with 200 B. pahangi L3 and 50 D. immitis L3 via subcutaneous injection. There were no D. immitis and B. pahangi recovered at the end of the study (218–222 days post-infection) in the group that received doxycycline 0–29 days post-infection (dpi), which spans over the L3 and L4 molts of the parasites (McCall et al. 2011).
Wolbachia is a Rickettsia-like, intracellular, bacterial endosymbiont that is found in a variety of filarial nematodes that cause filariasis and dirofilariasis (Taylor et al. 2013). Though the roles of this endosymbiont require further investigation, some studies have provided insights into the functions of this bacterium, indicating that five metabolic biosynthetic pathways are only found in Wolbachia and not in its nematode hosts: heme, riboflavin, flavin adenine dinucleotide (FAD), glutathione, and nucleotide synthesis pathways (Darby et al. 2012; Foster et al. 2005). Brugia malayi does not harbor six out of seven genes required for heme de novo synthesis and lacks all five genes required for riboflavin biosynthesis. However, the de novo biosynthesis pathways of purines and pyrimidines are found in Wolbachia from B. malayi, which suggests that Wolbachia may support nucleotide synthesis for the host, especially in oogenesis and embryogenesis during which DNA synthesis is in high demand (Foster et al. 2005). The depletion of Wolbachia sterilizes several filarial nematode species, including B. pahangi, B. malayi, and D. immitis, which adversely impacts the production of mf (Bandi et al. 1999; Rao & Weil 2002).
Wolbachia has been suggested to be required for the molting activities in B. malayi and B. pahangi (Casiraghi et al. 2002; Quek et al. 2022). Casiraghi et al. (2002) assessed the impact of different administration times of tetracycline treatment on B. pahangi using the gerbil (Meriones unguiculatus) model focused on the fourth-stage to immature adult molt. The gerbils were infected with 100 B. pahangi L3 and were euthanized at 54 dpi. The outcome was evaluated based on the number of worms recovered and the levels of Wolbachia persisted. Their results indicate that tetracycline treatment impacted the B. pahangi L4 to immature adult molt in vivo. All groups of worm recovered showed a significant decrease in Wolbachia levels. The gerbils that received tetracycline before the molt of both sexes showed a reduced worm recovery. The gerbils received treatment after the male molted but before the molt of females had sex-ratio distorted worm recovery (Casiraghi et al. 2002).
McCall et al. (2014, 2023) investigated the ability of D. immitis L3 to complete normal development in naïve dogs when the inoculated L3 were produced from the blood of doxycycline-treated microfilaremic dogs (McCall et al. 2023, 2014). In the study published in 2014, the microfilaremic dogs received doxycycline at 10 mg/kg, BID, for 30 days. The blood was collected on days 73 to 77 post-doxycycline treatment and fed to mosquitoes. The authors then harvested L3 from the mosquitoes fed from the blood of the doxycycline-treated dogs, which were used to infect naïve dogs. Dogs infected with the L3 received blood examinations for mf and antigen beginning 5 months after inoculation until 302 dpi, when they were necropsied and examined for adult worms. No mf or detectable antigens were observed in the blood, and no live or dead adult D. immitis were recovered. Recovery of worms from dogs infected with L3 developed from the blood of untreated microfilaremic dogs averaged ten live male and 17 live female heartworms (McCall et al. 2014).
In the study published in 2023 (McCall et al. 2023), the authors investigated the ability of D. immitis L3 to develop from mf in the blood of dogs treated with doxycycline (10 mg/kg, once daily for 30 days) and ivermectin (6 µg/kg, given on days 0 and 30) at different time points throughout the treatment regimen. Blood was collected on days 22, 29, and 42 after the beginning of the treatment, and L3 produced by mosquitoes fed this blood were inoculated into naïve dogs (30–50 L3 per dog). The subsequently infected dogs were necropsied 163–183 dpi, and no adult worms were recovered from dogs that had received L3 developed from doxycycline/ivermectin-treated mf. However, dogs that received L3 developed from untreated mf had a recovery of 26–43 adult worms from each dog (McCall et al. 2023). However, no measurement of Wolbachia DNA in either mf or L3 was performed in the above two studies, leaving a gap in knowledge regarding the correlation between Wolbachia and the ability of L3 to develop into adult worms in subsequent hosts. We also do not know at which point the worms stopped developing in the host (L3 to L4 molt, L4 to immature adult molt), or whether the depletion of Wolbachia affected the immunomodulatory effect that the worms have against the host.
Our study explores the impact of doxycycline on mf and subsequently produced L3 through the measurement of Wolbachia DNA in mf and L3 at different times throughout the AHS-recommended doxycycline treatment duration of 28 days. Previous studies have demonstrated an inhibition of molting in Brugia spp. with tetracycline treatment (Casiraghi et al. 2002; Quek et al. 2022; Rao & Weil 2002; Smith & Rajan 2000), so we wanted our study to focus on the first molt after the D. immitis L3 enter the host. Due to limitations in the ability to track D. immitis L3 to L4 molting in vivo, we established a D. immitis in vitro culture/molting system to observe the impact of Wolbachia on molting.
Method
A summary of the experimental design is shown in Fig. 1.Fig. 1. Experimental design flow chart. The animals received sham treatment or doxycycline treatment at 10 mg/kg for 28 consecutive days (4 weeks). The blood was collected weekly for L3 production and mf isolation. L3 were set up for in vitro culture and observed for 9 days. The Wolbachia levels were measured in mf and L3
Study animals
All experiments with purpose-bred dogs were performed according to the University of Georgia Institutional Animal Care and Use Committee guidelines using approved Animal Use Protocol A2022 04–006.
Three dogs were experimentally infected with 50 D. immitis Missouri (MO) isolate and housed in a specific pathogen-free environment. After the dogs became microfilaremic (9 months post-infection), they were randomly sorted into two groups: one dog served as the negative control, and two dogs received doxycycline hyclate (Epic Pharma, LLC, Laurelton, NY) orally at 10 mg/kg twice daily for 28 consecutive days. Blood was collected from the jugular vein before the start of doxycycline treatment and weekly after the treatment began.
Microfilaria isolation from whole blood
The mf in blood were collected for DNA isolation and subsequent Wolbachia quantification. Whole blood was diluted with saponin solution, consisting of 0.2% saponin (Tokyo Chemical industry, Tokyo, JPN) and 0.85% sodium chloride (Sigma-Aldrich, St. Louis, MO) in distilled water at a 1:11 ratio (i.e., 3 mL blood with 33 mL solution). The mixture was incubated for 15 min at 37 °C for hemolysis. The hemolyzed sample was centrifuged at 850 × g for 10 min at room temperature, and the supernatant was discarded. The mf were washed twice by filling the tubes with phosphate-buffered saline (PBS) and repeating the centrifugation step. After the second wash, mf were resuspended with 10 mL PBS and quantified by averaging the count from two 10-µL aliquots and observing at 10X magnification via light microscopy. Each sample was diluted to approximately 1000–3000 mf per mL. Microfilariae were stored at − 80 °C until DNA isolation was performed.
Mosquito feeding and L3 collection
Blood samples from three dogs were used for the production of L3 and Wolbachia DNA analysis in this study. Laboratory-raised Aedes aegypti, black-eyed, Liverpool strain were cultured and fed with blood collected from control and treatment groups (the blood from each animal was fed separately at each time point). Mosquitoes were harvested, and L3 were collected 15 days post-feeding per standard FR3 procedures (SOP numbers 8.2–8.4) (FR3, accessed 2024) with minor alterations. Briefly, mosquitoes were stunned at low temperatures, gently crushed in a mortar with a pestle, transferred to a 32-µm mesh sieve, quickly washed, and then soaked with 2% ciprofloxacin (Sigma-Aldrich, St. Louis, MO)-supplemented Hanks’ balanced salt solution (instead of Hanks’ balanced salt solution supplemented with penicillin–streptomycin as described in SOP 8.4). The L3 were manually quantified under light microscopy.
DNA isolation and quantitative real-time PCR
DNA samples were isolated from mf (see additional file Table S2 for details) and L3 of the control and treatment groups. DNeasy Blood & Tissue kit (QIAGEN, Valencia, CA) was used for DNA isolation. All samples were processed according to the manufacturer’s instructions. Isolated DNA was stored at − 20 °C for qPCR analysis.
Quantitative PCR was performed to determine the ratio of Wolbachia ftsZ DNA to D. immitis 18S ribosome DNA using the primer set listed (Table 1). SsoAdvanced™ Universal SYBR® Green Supermix (Bio-Rad, Hercules, CA) with Bio-Rad CXF96 Touch Real-Time PCR Detection System was used. The reactions were designed based on the protocol provided with the SYBR Green Supermix mentioned above. Briefly, the reaction was started at 98 °C for 2 min, followed by 40 cycles of 95 °C for 15 s, 54 °C for 30 s, 60 °C for 30 s, signal detection, and melting curve generation (from 60 to 90 °C, at 0.5 °C interval). Primers were diluted to 500 nM for all reactions in each well, and the loading amount of DNA template ranged from 1 to 10 ng. Raw data were collected with CFX Maestro Software version 3.1.1517.0823 (Bio-Rad). Table 1. Primers used for Wolbachia DNA quantificationGene namePrimer sequence (5’–3’)ReferenceD. immitis 18S rRNAForwardTGAGAAACGGCTACCACATCGenBank: AF036638ReverseGATAACCGGCCTCATAGAGAACD. immitis Wolbachia FtsZForwardGCTGGTGCCTTACCTGATATTGenBank: AJ495000 (Savadelis et al. 2018)ReverseCCACCCATTCCTGCTGTTAT
In vitro molting assessment
Third-stage larvae were washed according to a protocol from the Zamanian lab at the University of Wisconsin—Madison (Mostafa 2024) (Protocols – parasitic nematodes – filarial nematodes – Dirofilaria immitis L3 to L4 molt) with a few changes. Briefly, we used a different media and antibiotic mix compared to the protocol mentioned above. L3 were transferred to a 1.5-mL microcentrifuge tube with 500 µL of wash media RPMI-1640 with L-glutamine (Lonza Bioscience, Walkersville, MD) supplemented with 100 U/mL penicillin and 100 µg/mL streptomycin (Penicillin–Streptomycin solution, Thermo Fisher Scientific, Waltham, MA), 10 µg/mL ciprofloxacin, and 100 µg/mL gentamycin (Sigma-Aldrich, St. Louis, MO) and centrifuged at 1000 × g for 10 min at room temperature. The concentrated L3 were transferred to new microcentrifuge tubes pre-filled with 500 µL washing media. The centrifugation and transfer steps were repeated twice. The L3 were transferred to a Petri dish containing culture media consisting of the washing medium supplemented with 10% heat-inactivated fetal bovine serum (FBS; Thermo Fisher Scientific, Waltham, MA). The L3 were examined under a dissecting microscope, and individual L3 were aliquoted to single wells of a 96-well plate. An additional 200 µL of culture medium was added to each well containing a single L3. The L3 were cultured in an incubator at 37 °C and 5% CO_2_. A pan of autoclaved water was placed in the incubator throughout the culture period.
An optical microscope was used to observe the L3 at 400X magnification for nine consecutive days. The presence of a fully detached cuticle was defined as a successful molt.
Data analysis
The qPCR results were collected as Ct values. All Ct values were processed by CFX Manager software (Bio-Rad, Hercules, CA, USA, 3.1.1517.0823), and 2^−ΔΔCt^ (fold change) was calculated using the Livak method (Livak & Schmittgen 2001). The readouts for the Wolbachia ftsZ gene for each sample were normalized to the internal control (D. immitis 18S rRNA gene) and then to the control at each time point.
Fisher’s exact test was applied to determine the dependency of doxycycline treatment versus in vitro L3 to L4 molting rate. To calculate whether the molting event significantly differed between the control and treated groups, the Log-rank (Mantel-Cox) test was compared to the survival curves. Bonferroni correction was applied to both analyses.
Results
Quantitative real-time PCR
The Wolbachia levels in mf and L3 demonstrated a similar trend (Fig. 2). Following 7 days of treatment, the Wolbachia levels in treatment groups decreased to < 5% of the control group in both mf and L3 and continued to decrease as the treatment progressed.Fig. 2. Fold-change of Wolbachia DNA in D. immitis mf and L3. Wolbachia DNA were measured using qPCR with ftsZ/Di 18 s rRNA DNA ratio in both mf (A) and L3 (B). The results indicate a substantial decrease after 7 days of doxycycline (< 5% of the control group) and remained at low levels throughout the treatment
In vitro L3 to L4 molting
We tested different culture conditions for in vitro molting with non-treated D. immitis L3. (additional file Text S1). Based on the observation (molting rate, death rate, and the time-to-molt event, data not shown), we decided that 200 µL RPMI-1640 media containing 10% FBS is sufficient to support the in vitro survival and molting of one D. immitis L3 for at least 9 days without media change, and the 9-day observation could include most of the molting events.
We defined a completed, successful molt by the identification of a fully detached cuticle in the culture plate. Using Fisher’s exact test, we analyzed whether the successful molting correlated with doxycycline treatment by examining the number of L3 that had molted by the end of the observation period. There was no significant difference in the number of L3 that molted between the control and the doxycycline-treated parasites, with the exception of week 2 (Fisher’s exact test, p = 0.008) (Table 2). The L3 that molted in week 2 from the control group reached a molting rate of 83.3%, while the molting rates of the other weeks were 31.3%, 72.9%, 51.2%, and 51% for weeks 0, 1, 3, and 4, respectively. The molting rate for the treatment group reached 60.8% on week 2, while the molting rates of the other weeks were 26.8%, 82.3%, 63%, and 64.6% for weeks 0, 1, 3, and 4, respectively. Table 2. Number of molted control and doxycycline-treated L3DoxycyclineWeek 0Week 1Week 2Week 3Week 4- + - + - + - + - + MoltYes15263579405921582562No3371131783820342434p#0.69540.19940.0076**0.25170.1515#Fisher’s exact test
The time-to-molt event was analyzed by recording the day that the L3 molted and excluding any L3 that died before the observation endpoint. We did not observe any significant impact of doxycycline on the time-to-molt events. We used the log-rank (Mantel-Cox) test in Kaplan–Meier survival curves to compare molting activity based on doxycycline treatment. The molting pattern was not significantly affected by the doxycycline treatment (Mantel-Cox test; week 0, df = 1, p = 0.622; week 4, df = 1, p = 0.085) (Fig. 3).Fig. 3. Time-to-molt events of D. immitis L3 through doxycycline treatment. The observation of a fully detached cuticle indicated the success of the molt event. Deaths of L3 that occurred before molt were excluded from the group. The percent molt represents the ratio of successfully molted L3 on a given day after the start of the culture to the total number of L3 included in the experiment. The doxycycline treatment in all weeks does not significantly impact the time-to-molt event
Discussion
Heartworm disease affects multiple hosts (e.g., felids, bears, ferrets, seals, sea lions, and humans) and is diagnosed in all continents except Antarctica (Nelson 2023). Doxycycline is an important component of heartworm treatment plan due to its effect on the bacterial endosymbiont Wolbachia (Bazzocchi et al. 2008; Taylor et al. 2013). Previous studies by McCall et al. (2014, 2023) (McCall et al. 2023, 2014) indicated that D. immitis L3 cultured from doxycycline-treated mf can develop into viable L3 in mosquitoes but cannot develop into adults in subsequent vertebrate hosts. Based on the results of these previous studies, we inferred that the removal of Wolbachia leads to the inability of D. immitis to complete its life cycle. However, the mechanism behind this remains to be determined. Hypotheses include that the inhibition of L3 to L4 molt, the inhibition of L4 to immature adult molt, and the depletion of Wolbachia lead to altered host immune reactions against the worms.
We focused on the L3 to L4 molt in this study, which occurs days 3–12 post-infection in the host (Kotani & Powers 1982). Due to the lack of an in vivo tracking method of L3, we evaluated and established an in vitro model and observed the L3 for nine consecutive days. We hypothesized that L3 developed from doxycycline-treated mf would be unable to molt to L4 in vitro. Our results failed to support our hypothesis. The results demonstrate that the amount of Wolbachia DNA in mf and L3 was reduced substantially 7 days (week 1) after the beginning of the AHS recommended 28-day (4 weeks) doxycycline treatment and remained < 5% of the control group throughout the treatment. However, the successful molting rate and time-to-molt event were not impacted (Table 2, Fig. 3) with the exception of the successful molting rate for week 2 (p = 0.0076) (Fig. S1). The significance of the molting rate at week 2 may be due to biological variation from different batches of L3. A previously published study (Reaves et al. 2018) on Brugia malayi mf also found significant biological variation between different batches of mf. In the study, researchers obtained B. malayi mf from the gerbils’ peritoneal cavity and co-cultured them with human neutrophils and monocytes to observe the killing effect. Despite no other treatment having been done, the results of the replicates of these experiments were significantly different. Our study focused on whether the absence of Wolbachia impacts the molting of D. immitis L3 to L4 in vitro, and the results that Wolbachia-depleted L3 could still finish the molt indicated a pronounced trend towards our conclusion—the molting was not impacted.
Our study investigated the impact of doxycycline without ivermectin at the preventive dose to reduce potential variations due to other factors besides Wolbachia elimination, which is microfilaricidal over time and may impact the general health of the mf (Bowman & Mannella 2011). Our data demonstrate that Wolbachia DNA is almost non-detectable in mf and L3 after doxycycline treatment. While Wolbachia DNA had been examined in mf in previous studies, it had not been examined in L3 derived from mosquitoes fed blood from doxycycline-treated dogs. These results suggest that the inability of mf from doxycycline-treated dogs to complete their life cycle in subsequent hosts is due to disruption in the life cycle at a developmental stage beyond the L3 molt.
Multiple studies have been performed examining the effect of tetracycline treatment on the molting of filarial nematodes, including B. malayi, B. pahangi,* O. volvulus*, and D. immitis (Albers et al. 2012; Quek et al. 2022; Rao et al. 2002; Smith & Rajan 2000). Our data indicate that the presence of Wolbachia may not be required for D. immitis L3 to molt to L4 in vitro*.* Smith et al. (2000) examined the in vitro impact of tetracycline against the development of several filarial nematodes, and their results indicate inhibition of molting. They cultured B. malayi and B. pahangi with Rhodotorula minuta supplemented by arachidonic acid, while D. immitis was cultured without cellular coculture. Tetracycline (10 µg/ml) was added at the beginning of the culture (10 days for Brugia spp., 3 days for D. immitis). Tetracycline treatment inhibited B. malayi L3 to L4 molting by 93.9% of the controls on day 10, and the D. immitis L3 to L4 molt reached inhibition at 57.5% of the controls on day 3. They performed PCR to determine the amount of Wolbachia DNA in each sample, and samples collected from tetracycline-treated larvae still showed amplified bands on gel electrophoresis (Smith & Rajan 2000). There are several potential explanations for the discrepancy between their study and ours. The longer duration of treatment may explain the decrease of Wolbachia in our L3 samples. The timepoint we tested for Wolbachia DNA reduction was 7 days after the beginning of the treatment (week 1), while Smith et al. (2000) tested for Wolbachia 3 days post-treatment. The treatment conditions (in vitro vs. in vivo) could contribute to the difference in molting results. The actual medication used (tetracycline vs. doxycycline, with doxycycline being more lipophilic (Barza et al. 1975) and the endpoint of molting observation (3 days vs. 9 days) may also impact the outcomes.
There are some differences in L4 obtained from in vitro or in vivo molting. Marriott et al. (2023) developed an immunodeficient mouse model and obtained in vivo–developed L4 from inoculated L3 (Marriott et al. 2023). Alternatively, they cultured L3 in vitro with DMEM supplemented with only 10% FBS or with the addition of Madin-Darby canine kidney cells or rhesus monkey kidney epithelial cells in addition to the FBS. The authors compared the morphology of L4 as well as the amount of Wolbachia DNA in L4 obtained from mice 14 dpi and after 14 days of in vitro culture. The L4 cultured in vitro were significantly smaller compared to the L4 cultured in vivo. In vitro–derived L4 also presented with microscopic degenerative phenotypes, including malformed cuticle, hypodermis, buccal cavity, esophagus, and intestine (Marriott et al. 2023). We observed some deformed L3 during the in vitro culture process in both the control and treatment groups (data not shown). However, we could not rule out whether this was due to manipulation of the worms. The amount of Wolbachia DNA in the in vivo–derived L4 expanded 66-fold on average, while the in vitro–derived L4 did not display a notable change in Wolbachia. This could suggest that the host immune system, the nutrition that L3 could receive, as well as the host-parasite interaction, contribute to the fitness of the worms and their endosymbiont, Wolbachia, thus explaining the differences in the amount of Wolbachia DNA between in vivo and in vitro–derived larvae.
Wolbachia DNA in mf were measured weekly throughout the 28-day doxycycline treatment in a clinical study (Savadelis et al. 2018). Savadelis et al. (2018) studied eight dogs diagnosed with heartworm disease with doxycycline treatment at 10 mg/kg twice daily for 28 days and a monthly dosage of ivermectin/pyrantel (Heartgard® Plus, Boehringer Ingelheim, Duluth, GA). The dogs were brought to the clinic weekly for blood collection and qPCR was performed to detect the amount of Wolbachia DNA in mf at weeks 0, 1, 2, 3, and 4 post-treatment. This previous study showed that six out of eight dogs tested positive for Wolbachia DNA in mf at week 1. All dogs were cleared of Wolbachia DNA in the mf at the end of the study (Savadelis et al. 2018). This difference may be due to several variables. The authors defined Wolbachia DNA positivity as the qPCR result that could amplify Wolbachia DNA within 38 cycles of the reaction. In contrast, our qPCR looked at the relative changes in Wolbachia DNA compared to D. immitis DNA, which focused on the difference in Ct values between the Wolbachia gene and the D. immitis gene. Also, clinical studies have limitations in terms of animal conditions, the original burden of infections, and the compliance of pet owners. Those aspects may impact the efficacy of doxycycline treatment, thus leading to a delay in Wolbachia DNA reduction.
Answering the questions of whether the D. immitis L3 can also molt in vivo and exactly how doxycycline treatment inhibits D. immitis development in vivo requires further investigation. Tracking the L3 once they enter the host is time-consuming and cost-intensive (Kotani & Powers 1982). For our study, we optimized an in vitro D. immitis L3 to L4 molting system with nutrients supplied by only RPMI-1640 and heat-inactivated FBS, sufficient to support the overall survival for at least 9 days. Though widely used in cell culture, the components of FBS are complicated and may differ between manufacturers and batches. The genetic analysis revealed the potential metabolic role of Wolbachia in filarial nematodes, including the synthesis of heme and glutathione (Darby et al. 2012; Foster et al. 2005). Studies focused on FBS implied a likelihood of the existence of heme (Wagner et al. 2007), and glutamic acid and glutamine were found (Lee et al. 2023). We believe that FBS should not act as a replacement for Wolbachia and therefore interfere with the experimental outcome. The previous finding demonstrated that in vivo molting of tetracycline-treated B. malayi and B. pahangi was impaired (Casiraghi et al. 2002), indicating that Wolbachia contributes to molting in an environment that contains serum.
This system allows us to determine whether each cultured L3 can molt with fewer variables (i.e., cell viability if cell culture is involved). We demonstrated that doxycycline-treated D. immitis L3 can molt in vitro into L4. These results suggest that the inhibition of worm development in the host may occur after the L3 to L4 molt.
Conclusion
Our research demonstrated that doxycycline reduces Wolbachia DNA levels in mf and subsequently derived L3 1 week after the start of the AHS-recommended 28-day treatment. Our research also suggests that under in vitro conditions, the depletion of Wolbachia via in vivo doxycycline treatment does not impact the successful molting rate or the time-to-molt event of D. immitis L3 to L4.
Supplementary Information
Below is the link to the electronic supplementary material. ESM 1Supplementary file1 Time-to-molt events of D. immitis L3 through doxycycline treatment of weeks 1, 2, and 3. The observation of a fully detached cuticle indicated the success of the molt event. Deaths of L3 that occurred before molt were excluded from the group. The percent molt represents the ratio of successfully molted L3 on a given day after the start of the culture to the total number of L3 included in the experiment (PNG 227 KB)High Resolution Image (TIF 636 KB)Supplementary file2 Information of animals used in experiment (DOCX 17 KB)Supplementary file3 Microfilariae isolation and DNA isolation (DOCX 19 KB)Supplementary file4 In vitro culture condition testing (DOCX 104 KB)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Foster J, Ganatra M, Kamal I, Ware J, Makarova K, Ivanova N, Bhattacharyya A, Kapatral V, Kumar S, Posfai J, Vincze T, Ingram J, Moran L, Lapidus A, Omelchenko M, Kyrpides N, Ghedin E, Wang S, Goltsman E, . . . Slatko B (2005) The Wolbachia genome of Brugia malayi: endosymbiont evolution within a human pathogenic nematode. P Lo S Biol 3(4):e 121. 10.1371/journal.pbio.003012110.1371/journal.pbio.0030121 PMC 106964615780005 · doi ↗ · pubmed ↗
- 2FR 3. (accessed 2024). Filariasis Research Reagent Resource Center Protocols. Retrieved October, 25 from https://sites.uwosh.edu/filariasis-center/protocols/parasite-protocols/
- 3Kotani T, Powers KG (1982) Developmental stages of Dirofilaria immitis in the dog. Am J Vet Res 43(12):2199–2206. https://www.ncbi.nlm.nih.gov/pubmed/71651657165165 · pubmed ↗
- 4Mc Call JW, Genchi C, Kramer LH, Guerrero J, Venco L (2008) Chapter 4 Heartworm disease in animals and humans. In Advances in parasitology (Vol. 66, pp. 193–285). Academic Press. 10.1016/S 0065-308X(08)00204-210.1016/S 0065-308X(08)00204-218486691 · doi ↗ · pubmed ↗
- 5Mostafa Z (Accessed 2024) Zamanian Lab Docx - Lab Protocols. Retrieved October 25 from https://www.zamanianlab.org/Zamanian Lab Docs/labmanual/
- 6Nelson CT (2023) Chapter 111. Heartworm and related nematodes. In Sykes JE (Ed.), Greene's Infectious Diseases of the Dog and Cat. Elsevier, Inc. https://www.sciencedirect.com/science/article/pii/B 9780323509343001117. (Reprinted from Fifth Edition)