Investigating interspecific mating in the thelytokous predatory mite Amblyseius herbicolus (Chant) (Acari: Phytoseiidae), with comparative observations from three sexually reproducing phytoseiid species
Keshi Zhang, Junlin Cao, Xintong Li, Zhi-Qiang Zhang

TL;DR
This study explores whether a thelytokous mite species can mate with males from other species, shedding light on its reproductive mechanisms.
Contribution
The study provides new insights into the mating behavior of a thelytokous mite species through comparative interspecific mating trials.
Findings
A. herbicolus females did not mate with heterospecific males within 24 hours.
Mating behaviors varied across species, with differences in pre-mating duration and spermatophore deposition.
No correlation was found between male body size, mating duration, and endospermatophore volume.
Abstract
Asexual reproduction can be advantageous in the short term but is generally considered an evolutionary dead end due to the lack of genetic diversity and the accumulation of deleterious mutations. The thelytokous predatory mite Amblyseius herbicolus (Chant) (Acari: Mesostigmata) shows potential for biological pest control, but its reproductive mechanism remains poorly understood. This study examined whether A. herbicolus females have retained mating ability by exposing them to morphologically similar heterospecific males from three sexually reproducing phytoseiid species: Amblydromalus limonicus (Garman & McGregor), Amblyseius lentiginosus Denmark & Schicha, and Neoseiulus cucumeris (Oudemans). No mating or insemination occurred between A. herbicolus females and heterospecific males within 24 h. In contrast, nearly all males successfully inseminated conspecific females in a follow-up…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
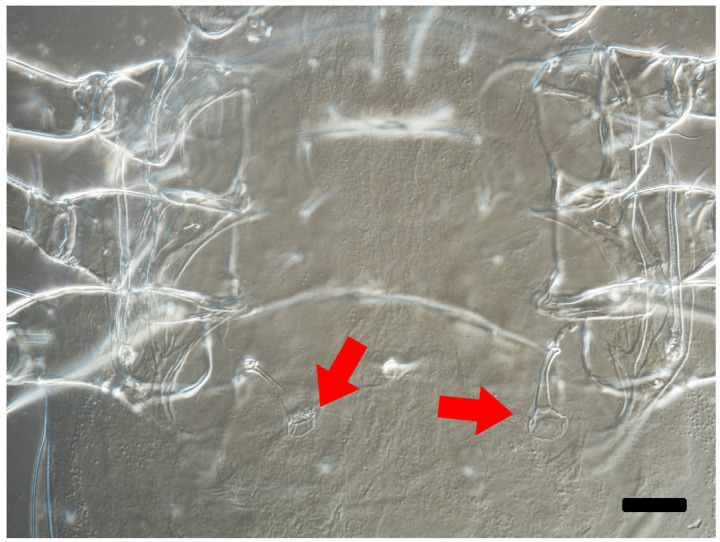 Figure 1
Figure 1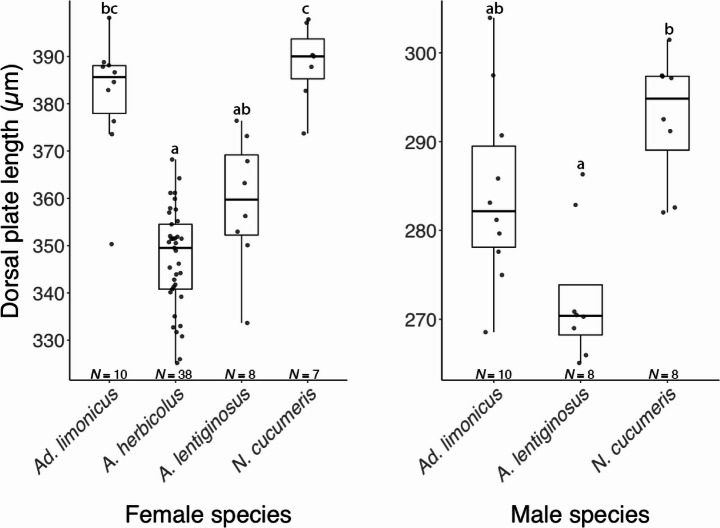 Figure 2
Figure 2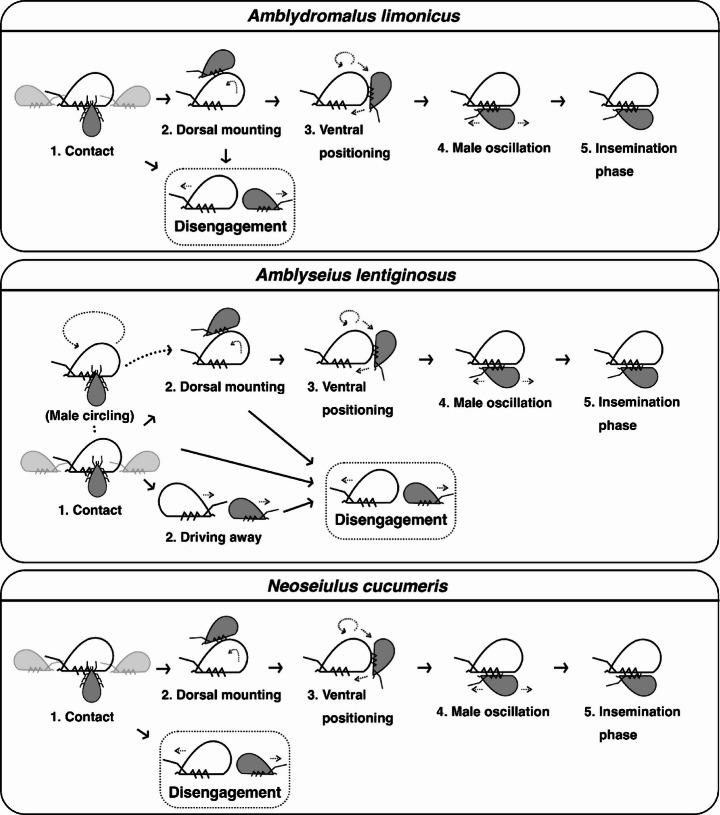 Figure 3
Figure 3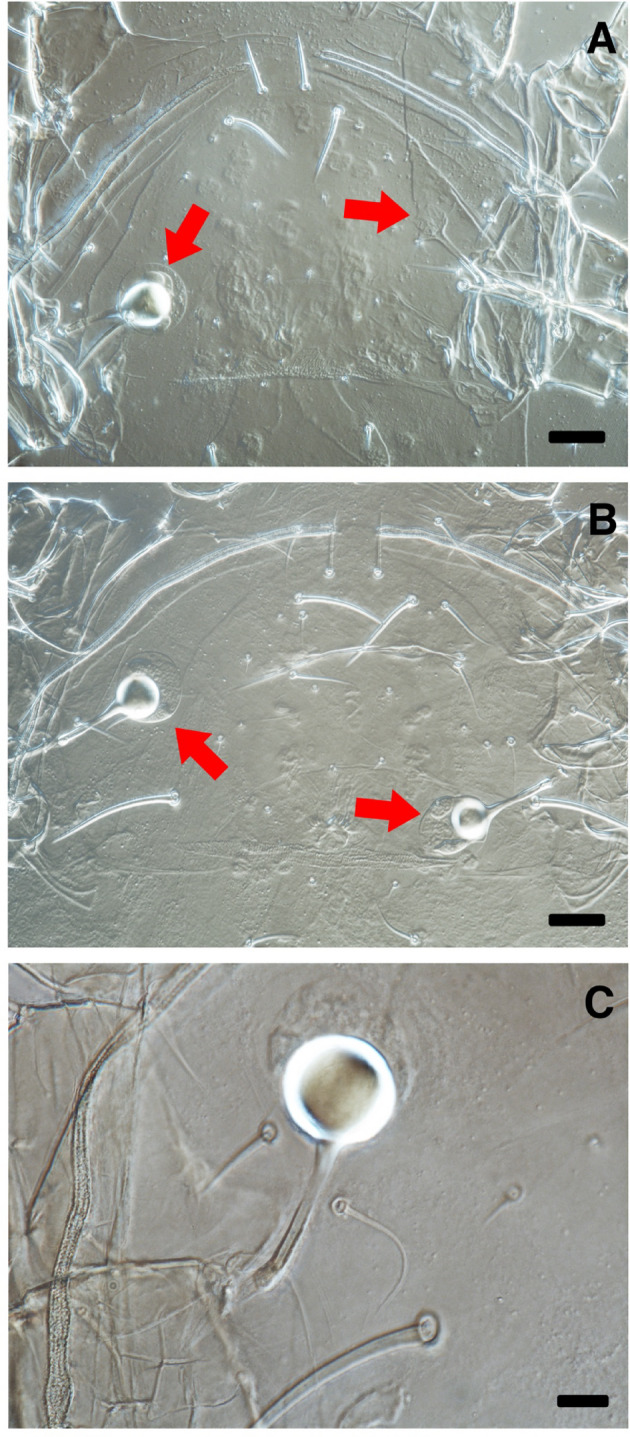 Figure 4
Figure 4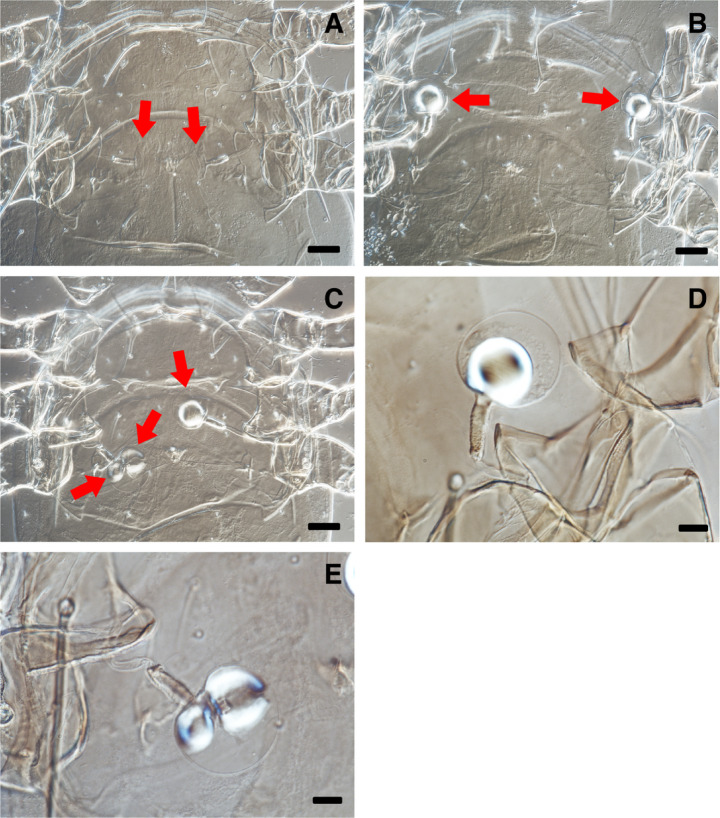 Figure 5
Figure 5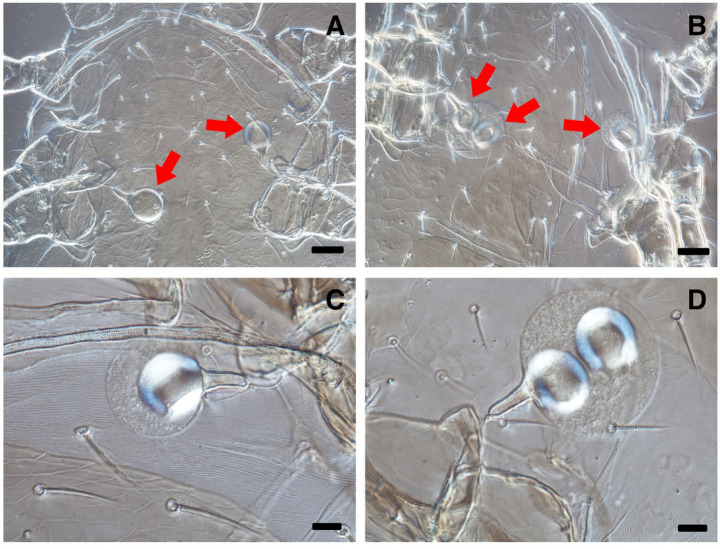 Figure 6
Figure 6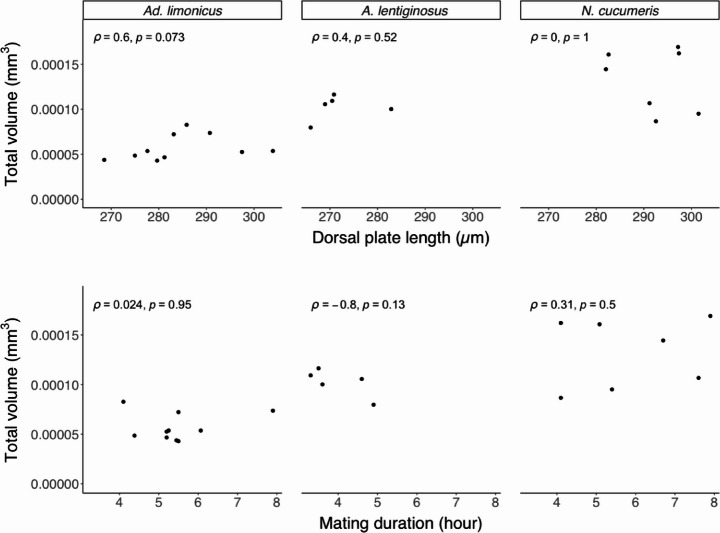 Figure 7
Figure 7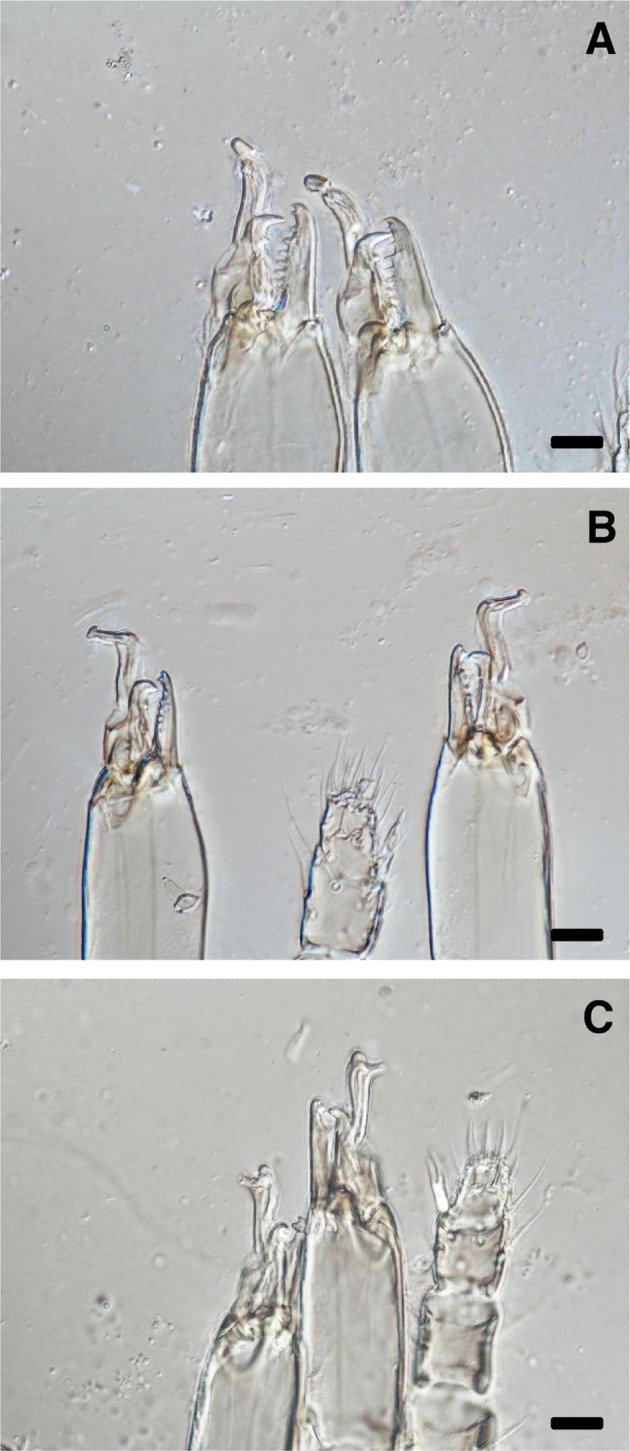 Figure 8
Figure 8- —Landcare Research New Zealand Limited
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect-Plant Interactions and Control · Plant and animal studies · Insect symbiosis and bacterial influences
Introduction
The family Phytoseiidae (Acari: Mesostigmata) contains primarily free-living, plant-inhabiting predatory mites, many of which are essential biological control agents against agricultural pests (McMurtry et al. 2015; Zhang 2003). Species such as Amblyseius swirskii, Neoseiulus cucumeris (Oudemans), and Phytoseiulus persimilis have been extensively utilised in pest management (Amano and Chant 1977; McMurtry et al. 2013; Zhang 2003). Maintaining a stable population of phytoseiid predators is crucial for effective pest management (Amano and Chant 1978; Xie et al. 2018; Xu et al. 2024). Consequently, their reproductive biology has been a key focus of research.
Although phytoseiid species are small (~ 300–450 μm in body length) and difficult to observe without proper tools such as a microscope, their reproductive behaviours have been well documented in various species (e.g., Amano and Chant 1979; Elbadry and Elbenhawy 1968; Hoy and Cave 1985; Pappas et al. 2005; Tsunoda 1994). Most phytoseiid species reproduce via pseudo-arrhenotoky, in which mating is required for oviposition (Hoy 1985; Norton et al. 1993). In this reproductive system females are diploid whereas haploid males arise through paternal genome elimination. Phytoseiid females possess a distinctive ‘phytoseioid-type’ sperm-access system that is unique within the order Mesostigmata (Alberti 2002; Moraza and Linquist 2011).
Unlike species with elaborate courtship displays, such as the peacock spider (Maratus volans) (Girard et al. 2011) or mites of the family Trombidiidae (Zhang 1999), phytoseiids exhibit relatively simple, yet diverse, mating behaviours (Elbadry and Elbenhawy 1968). Phytoseiid males are generally more active in mate searching than females (Amano and Chant 1979; Tsunoda 1994). During mating the male engages in a series of behaviours around and on the female before insemination, while the female remains relatively stationary. Phytoseiid mating behaviour has been classified into two types: ‘Amblyseius-Typhlodromus’ and ‘Phytoseiulus’ (Amano and Chant 1979). In the former, the male climbs onto the female’s dorsum after initial contact before moving underneath to assume a ‘venter-to-venter’ mating posture. In the latter, males directly invert themselves into this mating posture. In this posture males start to transfer sac-like spermatophores into the female’s sperm-access system via their spermatodactyl, a specialised structure on the moveable digits of phytoseiid males (Schulten 1985; Ullah et al. 2017a). Sperm is stored in two spermathecae, each receiving sperm through insemination pores located between the bases of the third and fourth legs. While a single mating event is sufficient for maximum lifetime reproductive success in some phytoseiid species, others require multiple inseminations (Amano and Chant 1978; Momen 1997; Schulten et al. 1978; Tsunoda and Amano 2001).
Most phytoseiid mites reproduce sexually, but at least 10 species reproduce asexually via thelytokous parthenogenesis (thelytoky) (Norton et al. 1993; Zhang 2022). However, males have been reported in some populations of thelytokous species such as Amblyseius elongatus (Oliver 1971) and Neoseiulus tunus (Cavalcante et al. 2017). Unlike pseudo-arrhenotoky, thelytoky enables female offspring to develop from unfertilised eggs laid by virgin females (Norton et al. 1993). The phytoseiid predator Amblyseius herbicolus (Chant) is one such thelytokous species, which is widely distributed and considered a promising biological control agent (Cavalcante et al. 2015; Kalile et al. 2021; Lam et al. 2021; Reis et al. 2007; Xin and Zhang 2021). Field collections and laboratory rearing have not identified males of A. herbicolus in New Zealand (Fan et al. 2024; Lam et al. 2021; Liu et al. 2023, 2024c; Ma et al. 2024; Zhang and Zhang 2021, 2022a, b, c, 2023; Zhang et al. 2024). Moreover, taxonomic studies of A. herbicolus have not reported the presence of males worldwide, except for a single study that documented their collection but did not provide a morphological description (Fang et al. 2022; Zhang 2022). Although some biological studies have suggested the presence of A. herbicolus males (Hou et al. 2022; Notghi Moghadam et al. 2010), these reports either misidentified the specimens or did not provide clear identification criteria.
Interspecific mating, particularly between morphologically similar species, can occur among mites (Reyer 2008; Ullah et al. 2017b). Some phytoseiid females accept interspecific matings, as observed in Neoseiulus womersleyi and Neoseiulus longispinosus, as well as in the spider mites Tetranychus evansi and Tetranychus urticae (Sato et al. 2016; Ullah et al. 2017b). However, these matings generally result in no or sterile offspring. Although A. herbicolus females possess spermathecae, it remains unclear whether they can behaviourally accept mating and be inseminated by males of closely related species. In an attempt to clarify this, we investigated interspecific matings between A. herbicolus females and males of three phytoseiid species: Amblydromalus limonicus (Garman & McGregor), Amblyseius lentiginosus Denmark & Schicha, and N. cucumeris. These four species vary in phylogenetic relatedness yet share a type-III generalist predator lifestyle by preying on various insect and mite species and utilising plant products such as pollen and nectar (McMurtry et al. 2013; Wang et al. 2024).
We hypothesised that A. herbicolus females would mate with and be inseminated by A. lentiginosus males, given their relatively close phylogenetic relationship (i.e., of the same genus) and morphological similarities, but not with the more distantly related Ad. limonicus or N. cucumeris. This study aimed to provide insights into the origin of the mode of reproduction in A. herbicolus and assess whether females of this species retain the ability to mate.
To confirm male viability of Ad. limonicus, A. lentiginosus, and N. cucumeris, we paired them with conspecific females and observed their mating behaviour. To our knowledge the mating behaviours of Ad. limonicus, A. lentiginosus, and N. cucumeris have not been previously reported. We hypothesised that their mating behaviours would be consistent due to their similar lifestyles. This study also aimed to enhance our understanding of mating behaviour and reproductive strategies in phytoseiid mites.
Materials and methods
Mite cultures
All mites used in this study were obtained within New Zealand and maintained under controlled laboratory conditions at Manaaki Whenua– Landcare Research (St Johns, Auckland, New Zealand) (see Liu et al. 2024b for details of the mites). The dried fruit mite, Carpoglyphus lactis (L.) (Acari: Carpoglyphidae), was used as prey to feed all examined phytoseiid predators. C. lactis was cultured on a Petri dish containing a mixture of wheat bran (~ 90%) (Goodman Fielder Limited, New Zealand), sugar (~ 5%) (Chelsea, New Zealand), and dry yeast (~ 5%) (Goodman Fielder Limited, New Zealand), all sourced locally.
To maintain the cultures, a rearing set-up was established using a plastic container filled with water. A sponge was placed inside the container, supporting a black plastic sheet, on which the Petri dish containing C. lactis was positioned (see Wang et al. 2024 for details of the rearing set-up). The water served two functions: restricting mite movement to the plastic sheet and providing a water source. Mixed-stage C. lactis, along with the bran, sugar, and yeast mixture, and water, were replenished regularly. Each colony was provided with an ad libitum supply of prey.
All cultures and experimental units were maintained in plexiglass cabinets under constant environmental conditions: 24 °C ± 1 °C, 80% ± 5% relative humidity, and a 16:8 h (light: dark) photoperiod.
Rearing cells
Both individual rearing and mating experiments of phytoseiid predators were conducted using modified Munger cells (see Zhang and Zhang 2021 for details). Each Munger cell consisted of two transparent plexiglass slides (38 mm × 25 mm × 2 mm). The upper plexiglass slide had a central conical hole (1 mm diameter at the top, narrowing to 0.6 mm at the base) to house the mites. The top of the hole was covered by a layer of food wrap, which was pierced with five small holes (created using a size 0 insect pin) to allow ventilation. A black plastic sheet disc (1.5 mm in diameter) was placed underneath to cover the base of the hole and provide contrast for better mite observation. A stack of filter papers below the disc served as a water reservoir, with the plastic disc pierced with five holes to allow access. The lower plexiglass slide, without apertures, was placed underneath the filter papers to maintain moisture. The two slides were secured with a pair of metal clips.
To minimise prey–predator interactions during the experiments and reduce potential mating interference, frozen feed (C. lactis) was prepared according to Liu et al. (2024a). The collected C. lactis was frozen at − 18 °C for at least 1 week and thawed at room temperature (~ 25 °C) for 30 min before being fed to the phytoseiid species during experimentation.
Experiment 1: interspecific mating
This experiment investigated whether A. herbicolus females would accept mating with heterospecific males. The procedures were as follows.
- Rearing: Similar-aged predator species were established by collecting nymphs (protonymphs or deutonymphs) from the main culture and rearing them individually in the rearing cells with an ad libitum supply of the frozen feed until maturity. Males of Ad. limonicus, A. lentiginosus, and N. cucumeris were used in this experiment, whereas females were used in Experiment 2.
- Pairing: One newly emerged adult female (< 24 h old) of A. herbicolus was paired with one newly emerged male (< 24 h old) of either Ad. limonicus, A. lentiginosus or N. cucumeris, in a new cell with an ad libitum amount of the frozen feed.
- Observation protocol: Mating pairs were observed every 15 min over an 8 h period under a dissection microscope to observe their interactions. Subsequently, pairs were left to mate overnight without observation, with final observations made at the 24 h mark.
- Endospermatophore verification: At the end of the 24 h period, females of A. herbicolus were slide-mounted (Walter and Krantz 2009) to check for the presence of endospermatophores, indicating successful insemination.
Experiment 2: intraspecific mating
This experiment evaluated whether males that had previously interacted with A. herbicolus females could successfully mate with conspecific females. The procedures were as follows.
- Pairing: With the same set-up as in Experiment 1, adult virgin females (< 72 h old) of Ad. limonicus, A. lentiginosus, and N. cucumeris (obtained from the rearing procedure used in Experiment 1) were paired with males of the same species that had previously interacted with A. herbicolus females for 24 h during Experiment 1.
- Observation protocol: Each mating pair was continuously observed under a dissection microscope from the start of the experiment until the onset of copulation (i.e., the female and male engage in the venter-to-venter position with minimal movement immediately after male oscillation). Subsequently, observations were made every 15 min until separation. Pairs where no mating was observed during the 8 h period were left to mate overnight without observation, with final checks made at the 24 h mark.
- Endospermatophore verification: Post-mating, females were slide-mounted to verify endospermatophore presence as evidence of successful insemination, as in Experiment 1.
To mitigate the high propensity for intra- and inter-specific predation within the Phytoseiidae family (Gu et al. 2024; Zhang and Zhang 2022c, 2023), sufficient food (frozen mixed-stage C. lactis) was provided to each mating pair throughout both Experiments 1 & 2.
The mating latency (time from pairing to the onset of copulation), pre-mating behaviours, and mating durations were recorded. All examined individuals were slide-mounted, and their dorsal plate length measured using NIS-Elements (version 5.10) under a phase-contrast microscope (Eclipse 90i, Nikon Corporation, Japan). The volumes of endospermatophores were estimated using the standard equation of a sphere (Ullah et al. 2017a).
The method for recording the degree of insemination of spermathecae was adopted from Amano and Chant (1979); the first number indicates the number of inflated vesicles (without endospermatophores), while the second number indicates the number of vesicles containing endospermatophores.
Statistical analysis
All statistical analyses were performed using R (R Core Team 2022) in RStudio (version 2023.12.1). Data visualisation was carried out with the ggplot2 package (version 3.4.3) (Wickham 2016). Data were summarised as means with standard errors of the mean (SEMs). Dorsal plate lengths were presented in box plots, showing the interquartile range (IQR; middle 50%), median, and data spread (1.5 times the IQR). Outliers, where present, were retained unless specified otherwise. Given the non-normal distributions of all data from the Shapiro–Wilk test, the Kruskal–Wallis test and Dunn’s test were applied to determine significance. Chi-squared tests were used to compare proportions. Spearman’s rank correlations were done to determine the relationship between the volume of endospermatophores, male size, and copulation duration. Statistical significance was set at p < 0.05.
Results
Experiment 1: interspecific mating
No mating events were observed between A. herbicolus females and males of the three predator species during the first 8 h of observation. Also, slide-mounted females examined after 24 h revealed neither endospermatophores nor inflated vesicles (Table 1; Fig. 1). Occasional brief contacts were noted between A. herbicolus females and males of the other species, but these encounters ended without prolonged interaction. A small proportion of males were preyed upon by A. herbicolus females, with no significant difference in predation rates among the three species (Chi-squared test: χ² = 0.375, df = 2, p = 0.829) (Table 1).
Table 1. Number of mating events (first 8 h) and endospermatophores detected in Amblyseius herbicolus females after 24 h of exposure to males of three predator species: Amblydromalus limonicus, Amblyseius lentiginosus, and Neoseiulus CucumerisMale N Mating eventsSpermatophoresMales eaten Ad. limonicus 13007.7% A. lentiginosus 140014.3% N. cucumeris 110018.2%
Fig. 1. Spermathecae of Amblyseius herbicolus under a phase-contrast microscope at 400× magnification. Arrows point to the vesicles. Scale bar = 25 μm
Significant differences were found in body size (i.e., dorsal plate length) among the predator species for both females (Kruskal–Wallis test: χ² = 34.989, df = 3, p < 0.001) and males (χ² = 10.541, df = 2, p = 0.005). Specifically, both males and females of N. cucumeris were the largest, while A. herbicolus females were similar in size to A. lentiginosus but approximately 10% smaller than Ad. limonicus and N. cucumeris (Fig. 2).
Fig. 2. Dorsal plate length of Amblyseius herbicolus females, and Amblydromalus limonicus, Amblyseius lentiginosus, and Neoseiulus cucumeris females and males. Box plots represent interquartile range (IQR), median, and data spread (1.5× IQR). Different letters denote significant differences (Dunn’s test: p < 0.05). Sample sizes (N) are given under each box
Experiment 2: intraspecific mating
The mating process in Ad. limonicus, A. lentiginosus, and N. cucumeris was categorised into five sequential stages (Fig. 3), as follows.
- Contact: Males initiated contact with females using their first pair of legs (Leg I). Some females also touched males with Leg I. Contact direction depended on male approach (front, behind, or side). Some males moved onto the female’s dorsum after the first contact. Only three A. lentiginosus males circled stationary females while touching them with Leg I before mounting.
- Dorsal mounting: Males mounted the female’s dorsum, mostly from the posterior end. Some disengaged after this stage. In one A. lentiginosus mating pair, the female drove the male away twice after the initial contact.
- Ventral positioning: Males turned around and moved to the female’s ventral side from the posterior. Females often lifted their bodies to assist males by creating more space.
- Male oscillation: Males performed oscillatory (back-and-forth or sideways) movements (or jerking) before becoming stabilised for insemination.
- Mating or insemination phase: Both sexes remained relatively stationary. Occasionally, females moved around with males attached to their ventrum.
Fig. 3. Stages of mating behaviour in Amblydromalus limonicus, Amblyseius lentiginosus, and Neoseiulus cucumeris
Endospermatophores were detected in all examined females following a single mating event, except in three A. lentiginosus females (Table 2; Figs. 4, 5 and 6). Two pairs of A. lentiginosus failed to copulate within 8 h, though one female from these pairs had endospermatophores after 24 h (Fig. 5C), suggesting delayed insemination. Also, two pairs of A. lentiginosus had early termination during mating (< 30 min), with no endospermatophores detected and no inflated spermathecae post-mounting. These data were excluded from further analysis of mating duration. The occurrence of mating was statistically similar among species (Chi-squared test: χ² = 4.547, df = 2, p = 0.103) (Table 2). However, successful insemination was significantly lower in A. lentiginosus than in the other species (χ² = 7.093, df = 2, p = 0.029).
Table 2. Proportion of successful mating events (0–8 h) and endospermatophores detected after a single mating event or at 24 h in Amblydromalus limonicus, Amblyseius lentiginosus, and Neoseiulus CucumerisMale N Mating eventsEndospermatophores Ad. limonicus 10100%100% A. lentiginosus 977.8%66.7% N. cucumeris 9100%100%
Fig. 4. Spermathecae of Amblydromalus limonicus under a phase-contrast microscope. The balloon-shaped structures within the vesicles are endospermatophores. A: A spermatophore found in one vesicle. B: One spermatophore is found in each of the vesicles. C: An enlarged view of a vesicle. Arrows point to the vesicles. The magnifications are 400× (A & B) and 1000× (C). Scale bars = 25 μm (A & B) and 10 μm (C)
Fig. 5. Spermathecae of Amblydromalus lentiginosus under a phase-contrast microscope. The balloon-shaped structures within the vesicles are endospermatophores. A: No spermatophores in the vesicles. B: Spermatophores are found in each of the vesicles. C: One spermatophore is found in one vesicle and two in the other. D: An enlarged view of a vesicle with one spermatophore. E: An enlarged view of a vesicle with two spermatophores. Arrows point to the vesicles. The magnifications are 400× (A–C) and 1000× (D & E). Scale bars = 25 μm (A–C) and 10 μm (D & E)
Fig. 6. Spermathecae of Neoseiulus cucumeris under a phase-contrast microscope. The balloon-shaped structures within the vesicles are endospermatophores. A: Spermatophores are found in one of each vesicle. B: One spermatophore is found in one vesicle and two in the other. C: An enlarged view of a vesicle with one spermatophore. D: An enlarged view of a vesicle with two spermatophores. Arrows point to the vesicles. The magnifications are 400× (A & B) and 1000× (C & D). Scale bars = 25 μm (A & B) and 10 μm (C & D)
Mating latency differed significantly among species (Kruskal–Wallis test: χ^2^ = 6.730, df = 2, p = 0.035), with Ad. limonicus initiating the most rapid insemination (Table 3). However, the number of initial contacts did not differ significantly across species (χ^2^ = 0.502, df = 2, p = 0.778). The duration of pre-mating behaviours was significantly longer in A. lentiginosus than in the other two species (χ^2^ = 10.676, df = 2, p = 0.005). This included extended durations for initial contact (χ^2^ = 8.608, df = 2, p = 0.014), dorsal mounting (χ^2^ = 9.922, df = 2, p = 0.007), and ventral positioning (χ^2^ = 10.161, df = 2, p = 0.006), whereas the duration of male oscillation did not vary significantly among species (χ^2^ = 4.399, df = 2, p = 0.111) (Table 3). Despite its prolonged pre-mating phase, A. lentiginosus exhibited the shortest mating duration among the three species (χ^2^ = 7.930, df = 2, p = 0.019) (Table 3).
Table 3. Pre-mating and mating parameters in mating of Amblydromalus limonicus, Amblyseius lentiginosus, and Neoseiulus cucumeris. All data are summarised as mean ± SEMSpecies N Mating latencyPre-mating durationMating duration (h)Duration (min)Contact (number)Contact (sec)Mounting (sec)Positioning (sec)Oscillation (sec)Total (sec) Ad. limonicus 105.1 ± 1.2^a^3.4 ± 1.17.2 ± 1.7^a^7.1 ± 1.4^a^2.4 ± 0.4^a^56.6 ± 6.373.3 ± 8.4^a^5.5 ± 0.3^b^ A. lentiginosus 722.7 ± 10.0^b^4.3 ± 1.536.3 ± 16.2^b^29.5 ± 8.4^b^9.0 ± 2.4^b^80.8 ± 7.7155.7 ± 29.6^b^4.0 ± 0.3^a^* N. cucumeris 911.7 ± 2.8^b^2.9 ± 0.510.7 ± 1.6^ab^7.4 ± 1.0^a^3.0 ± 0.4^a^68.3 ± 7.189.4 ± 6.4^a^6.0 ± 0.6^b^Note: Different letters denote significant differences (Dunn’s test, p < 0.05). * For A. lentiginosus, two pairs had a mating duration of < 30 min and lacked endospermatophores upon slide-mounting; these were considered early terminations and excluded from further analyses, reducing the sample size (N) for mating duration to five
The number of endospermatophores differed among species (Kruskal–Wallis test: χ^2^ = 6.730, df = 2, p = 0.035), with Ad. limonicus having fewer endospermatophores compared to N. cucumeris and A. lentiginosus (Table 4). Most females of A. lentiginosus and N. cucumeris received two or more spermatophores (typically one in each spermatheca, except for one N. cucumeris female that received two spermatophores in one spermatheca). In contrast, 70% of Ad. limonicus females had only one spermatophore, which was distributed between the left (~ 40%) and right (~ 60%) spermathecae; the side preference of male Ad. limonicus was not statistically significant (Binomial test: p = 1). Only one Ad. limonicus female had an inflated spermathecal vesicle without an endospermatophore (Table 4), whereas all other inflated vesicles contained at least one endospermatophore.
Table 4. Number, degree of insemination, and size (individual volume) of endospermatophore after one mating event of Amblydromalus limonicus, Amblyseius lentiginosus, and Neoseiulus cucumeris. Means ± SEM are given for the number and size of endospermatophoresSpecies N NumberDegree of insemination N Size (mm^3^)0–01–01–12–12–2 Ad. limonicus 101.3 ± 0.2 (1–2)^a^00613134.4 × 10^−5^ ± 2.1 × 10^−6 a^ A. lentiginosus 52.0 ± 0.0 (2)^b^00005134.8 × 10^−5^ ± 3.5 × 10^−6 ab^ N. cucumeris 72.1 ± 0.1 (2–3)^b^00007156.2 × 10^−5^ ± 5.0 × 10^−6 b^Different letters denote significant differences (Dunn’s test, p < 0.05). Degree of insemination: number of inflated vesicles minus number of inseminated vesicles
Spermatophore size also varied significantly among species (χ² = 9.354, df = 2, p = 0.009), with N. cucumeris producing the largest spermatophores, whereas those of A. lentiginosus were comparable to both N. cucumeris and Ad. limonicus (Table 4). However, there were no significant correlations between male body size, mating duration, and total volume of spermatophores for any of the three species (Fig. 7).
Fig. 7. Correlation between individual male size, mating duration, and the total volume of endospermatophores inseminated into females’ spermathecae of Amblydromalus limonicus, Amblyseius lentiginosus, and Neoseiulus cucumeris by each conspecific male during a single mating event. Spearman’s rank correlation coefficients () and p-values are given in each graph
Discussion
In this study mating and insemination between A. herbicolus females and males of Ad. limonicus, A. lentiginosus, and N. cucumeris were not observed (Experiment 1). However, all males, except three A. lentiginosus individuals, successfully inseminated a conspecific female when exposed to one (Experiment 2). These findings indicate that interspecific mating between A. herbicolus females and males of the other three species is improbable. In addition to differences in body size, these species exhibit distinct morphological characteristics (Zhang 2003).
It is possible that A. herbicolus females and the heterospecific males tested in this study might have been able to distinguish one another, as species recognition has been documented in members of Phytoseiidae, including N. cucumeris (Schausberger and Croft 2000). Moreover, some males of all three species examined served as intraguild prey for A. herbicolus, with no apparent predation preference among male species. However, the absence of mating behaviour does not necessarily indicate that A. herbicolus females are incapable of mating, either behaviourally or physiologically. Future studies could investigate their compatibility with Amblyseius largoensis (Muma), a more closely related species with greater morphological and genetic similarities to A. herbicolus (Ma et al. 2019; dos Santos and Tixier 2017).
The courtship sequences of conspecific pairs of Ad. limonicus, A. lentiginosus, and N. cucumeris were similar and conformed to the Amblyseius-Typhlodromus type described by Amano and Chant (1979). Since phytoseiid species lack functional eyes (Liu et al. 2024b), whether the mating sequence functions as a courtship display or a form of pre-mating stimulation requires further investigation. However, significant interspecific differences were observed in the duration of pre-mating and mating stages, which suggest possible species-specific adaptations. Whether these differences arise from variations in mate recognition, mate choice or courtship behaviour remains unclear. In addition to the longest pre-mating duration among examined species, A. lentiginosus exhibited a lower insemination rate than Ad. limonicus and N. cucumeris, with two incidents of early mating termination. Also, one A. lentiginosus female was observed chasing the male away after contact. These results suggest that A. lentiginosus females might be more selective.
The number and size of spermatophores inseminated after the first mating event further highlight interspecific differences in reproductive strategies (e.g., sperm allocation). Males of Ad. limonicus were similar to those of the phytoseiid predators P. persimilis and Kampimodromus aberrans, mostly inseminating one spermatophore to one of the spermathecae (Amano and Chant 1979; Pappas et al. 2005). In contrast, males of A. lentiginosus and N. cucumeris showed a pattern comparable to Amblyseius andersoni, typically inseminating two spermatophores, one into each spermatheca (Amano and Chant 1979).
The different numbers and sizes of endospermatophores between species may reflect a different level of sperm allocation to mating, or adaptations to maximise fertilisation success and male reproductive fitness. For example, in P. persimilis, which exhibits low levels of polyandry and where females often avoiding multiple matings (Amano and Chant 1978; Schausberger et al. 2017), males may not need to transfer excess sperm to ensure reproductive success. Conversely, in N. cucumeris, where polyandry is more common (Ji et al. 2007), males may benefit from inseminating both spermathecae to enhance their reproductive success. However, this hypothesis requires further investigation, particularly to determine the level of polyandry in Ad. limonicus and A. lentiginosus.
There are morphological differences in the structure of the spermatodactyl on the movable digit of the male chelicerae (Fig. 8). Whether these structural differences relate to their mating behaviour and degree of insemination requires further investigation.
Fig. 8. Chelicerae of the examined phytoseiid male species in this study, showing the spermatodactyl structure on the movable digits at 1000× magnification. A: Amblydromalus limonicus; B: Amblyseius lentiginosus; and C: Neoseiulus cucumeris. Scale bar = 10 μm
No correlation was found between male body size, mating duration, and total volume of endospermatophores after the first mating in any species examined. A positive relationship between mating duration and endospermatophore size has been reported in several phytoseiid species, including Neoseiulus womersleyi, Neoseiulus longispinosus, and P. persimilis (Schulten et al. 1978; Ullah et al. 2017a). The absence of such a correlation in this study may be due to all individuals being allowed to complete the full mating sequence or to the limited sample size. Future studies incorporating larger sample sizes may reveal more informative patterns. Also, investigating whether male body size influences lifetime sperm production, rather than sperm allocation per mating, could provide further insight into the selective advantages of larger body size beyond increased female receptivity and competitive ability (Schausberger et al. 2017).
One limitation of this study was that the prolonged confinement of A. herbicolus females with heterospecific males in enclosed arenas may have influenced subsequent mating behaviour with conspecifics. Females of A. herbicolus, as superior intraguild predators, may have induced stress in the males, subsequently affecting their physiological and behavioural responses (Gu et al. 2022, 2024). Furthermore, forced pairings in artificial conditions may not accurately reflect natural mating dynamics. The multiple contacts observed between pairs before insemination suggest a tendency to seek alternative mates. Future research on phytoseiid mating behaviour would benefit from more ecologically relevant experimental designs that incorporate natural choice conditions.
In conclusion, A. herbicolus females did not accept mating from heterospecific males of Ad. limonicus, A. lentiginosus, and N. cucumeris. Further research should investigate the mechanisms underlying and maintaining parthenogenesis in this species, such as the potential role of endosymbionts such as Wolbachia, which induce parthenogenesis in other insects and mites (Ramalho et al. 2021; Weeks and Breeuwer 2001). Also, the observed differences in pre-mating duration, mating behaviours, and insemination strategies among Ad. limonicus, A. lentiginosus, and N. cucumeris may reflect adaptations to distinct ecological pressures, including mate availability, predation risk, and sperm competition.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Liu Z, Zhang K, Zhang Z (2024 b) Phototactic behavior and oviposition of seven species of Phytoseiidae (Acari: Mesostigmata). 10.1002/ps.8575. Pest Manag Sci n/a 10.1002/ps.857539632773 · doi ↗ · pubmed ↗
- 2Xu Y, Zhang K, Han X, Zhang Z (2024) Early life food intake modulates effects of diet restriction on lifespan and fecundity in later life in a predatory mite (Acari: Phytoseiidae). 10.1093/cz/zoae 047. Curr Zool:zoae 04710.1093/cz/zoae 047PMC 1222741540620589 · doi ↗ · pubmed ↗
- 3Zhang K (2022) Kin recognition in a predatory mite Amblyseius herbicolus Chant (Acari: Phytoseiidae). Dissertation, University of Auckland