Functional characterization of the store-operated calcium entry pathway in naked mole-rat cells
Polina Drugachenok, Paulina Urriola-Muñoz, Lanhui Qiu, Zhuang Zhuang Han, Ewan St John Smith, Taufiq Rahman

TL;DR
This study is the first to investigate how a key calcium entry pathway works in naked mole-rat cells, which may explain their resistance to age-related diseases.
Contribution
The first functional and pharmacological characterization of store-operated calcium entry in naked mole-rats.
Findings
SOCE was functionally active in both excitable and non-excitable NMR cells.
Pharmacological tools confirmed the presence of a Stim-Orai-like mechanism in NMR cells.
The findings suggest SOCE may contribute to the NMR's disease resistance.
Abstract
Naked mole-rats (NMRs, Heterocephalus glaber) are highly unusual rodents exhibiting remarkable adaptations to their subterranean habitat and resistance to developing various age-related diseases such as those related to abnormal cell proliferation or cancer, neurodegeneration and inflammation. In other rodents, as well as humans, a ubiquitous Ca2+ influx pathway, namely the store-operated Ca2+ entry (SOCE), has been implicated in all these diseases. SOCE is triggered by intracellular Ca2+ store depletion resulting in interaction of Stim proteins with Orai proteins, the putative homologues of which appear to be present in the NMR genome, but no functional characterization of SOCE in NMRs has yet been conducted. In this study, we provide the first functional and pharmacological characterization of SOCE in NMR using both excitable and non-excitable cells.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
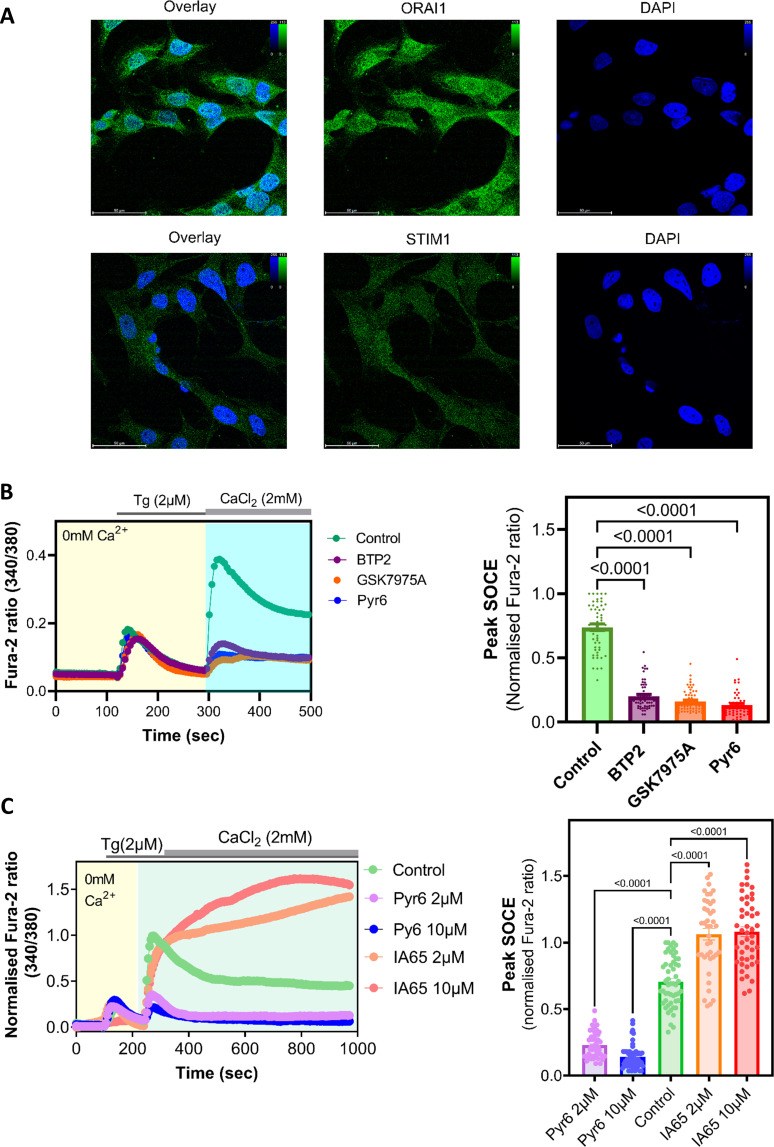 Figure 1
Figure 1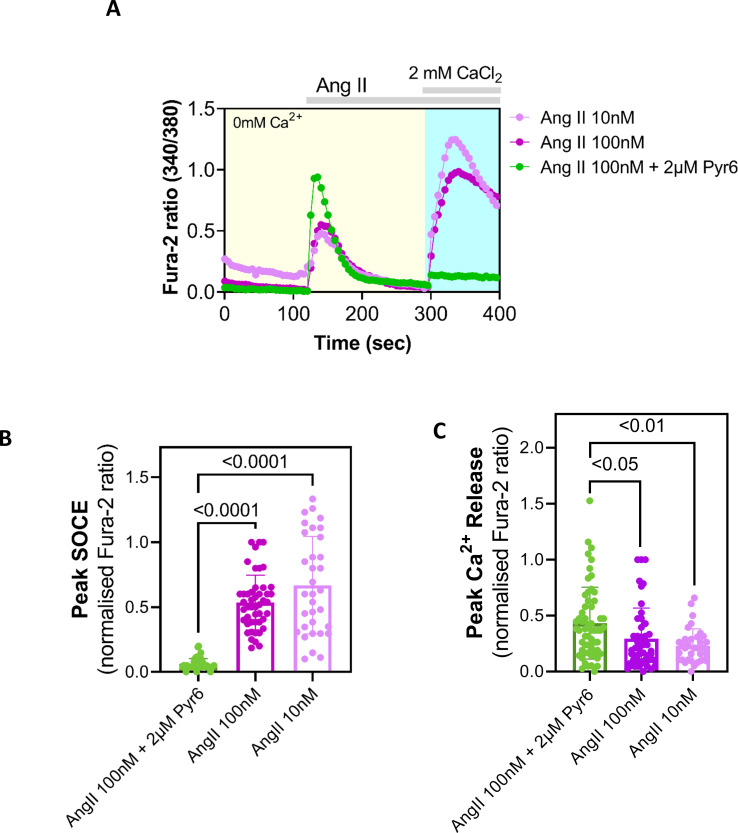 Figure 2
Figure 2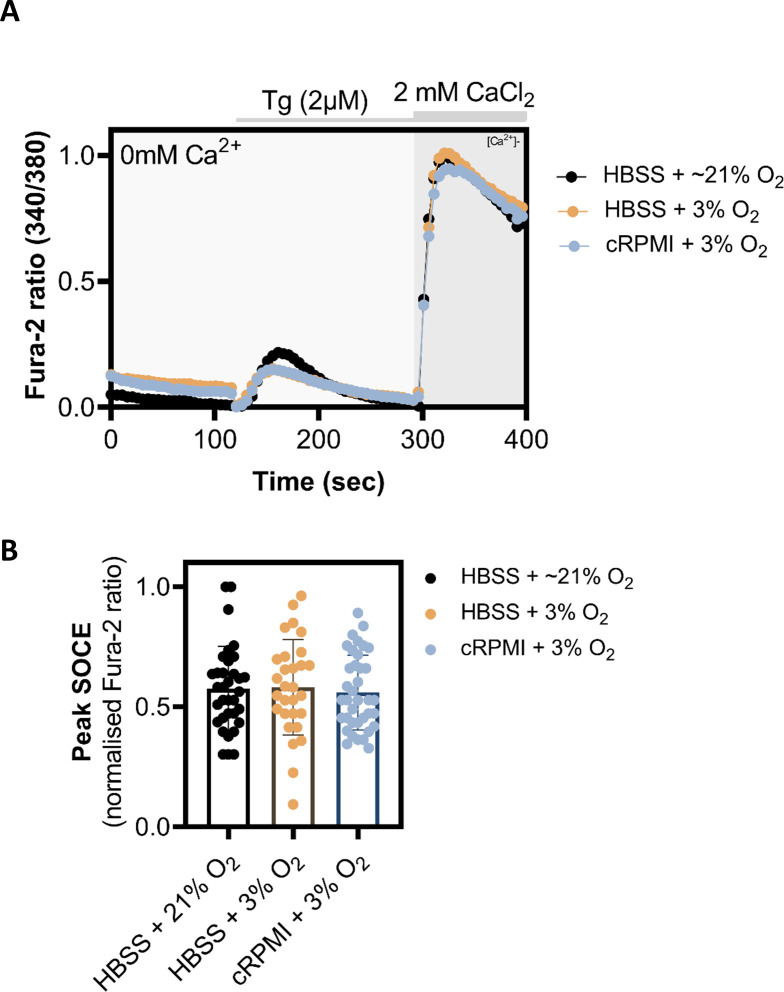 Figure 3
Figure 3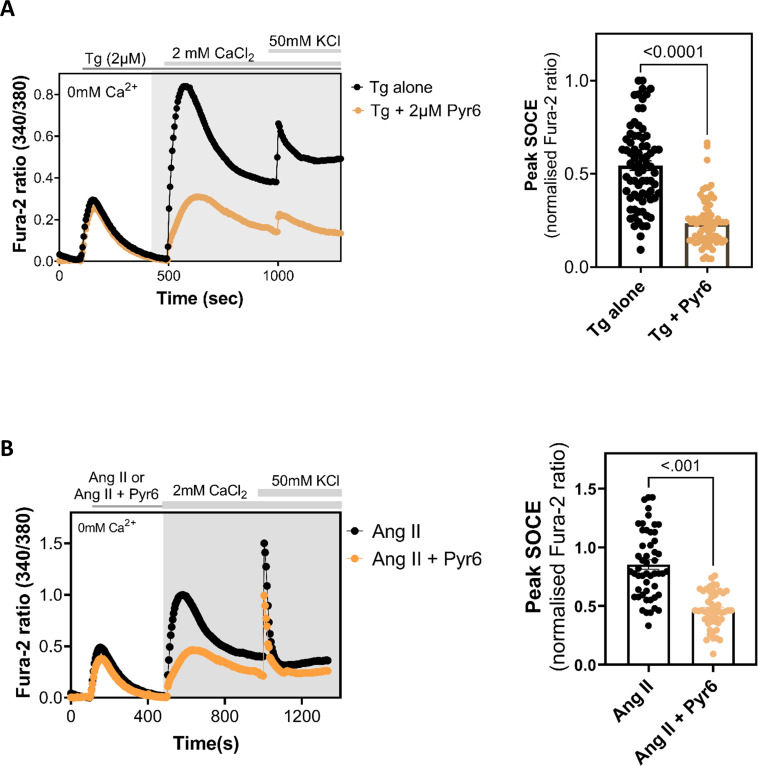 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeuroscience and Neuropharmacology Research · Ion channel regulation and function · Neurobiology and Insect Physiology Research
Introduction
The store-operated Ca^2+^ entry (SOCE) represents a route through which Ca^2+^ from the extracellular fluid enters into the cytosol in response to depletion of intracellular Ca^2+^ stores, such as the endoplasmic reticulum (ER) [1]. Store depletion occurs physiologically upon stimulation of cell surface receptors that produce the second messenger, inositol 1, 4, 5-trisphosphate (IP_3_), in a phospholipase C (PLC)-dependent manner. Pharmacologically, SOCE can also be triggered either by inhibiting (e.g. with thapsigargin or cyclopiazonic acid) the sarcoplasmic reticulum/ER Ca^2+^-ATPase pump, which allows uncompensated passive Ca^2+^ leak from the ER that eventually empties the store or by using a low concentration (≤10 μM) of ionomycin that mobilizes Ca^2+^ preferentially from the intracellular stores, thereby depleting them [1,2].
Initially, SOCE was thought to be an exclusive feature of non-excitable cells, but more recent studies have confirmed that this particular Ca^2+^ influx pathway is present almost in all cells including the excitable ones, although its prominence and indispensability can vary depending on the nature of the cell [1–3]. In most eukaryotic cell types, SOCE is mediated through Ca^2+^ release-activated Ca^2+^ (CRAC) channels. The latter are highly Ca^2+^-selective and, rather unusually for ion channels, formed in situ through protein–protein interaction. CRAC channel activation occurs following store depletion that induces interaction between clusters (formed after store depletion) of the ER-membrane localized Stim proteins and the plasma membrane (PM)-resident Orai proteins at the ER–PM junctions. Evidence continues to amass that CRAC-derived Ca^2+^ signals, besides their well-known ER-refilling housekeeping role, are often utilized for regulating specific cellular functions including gene transcription, secretion, modulation of enzyme activity and motility [1,2]. Abnormal SOCE due to aberrant CRAC channel activity seem to underlie various diseases, including various autoimmune and inflammatory diseases, neurodegenerative diseases, chronic pain and cancer [4,5].
The naked mole-rat (NMR, Heterocephalus glaber) is a long-lived, subterranean and eusocial mammal, which exhibits numerous signs of healthy aging alongside a remarkable tolerance to hypoxia and unusual pain phenotypes, as well as notable resistance towards developing certain age-associated diseases, such as cancer, neurodegenerative conditions and cardiovascular ailments [6]. Across pain, cancer and neurodegeneration, the immune system plays an important role. Intriguingly, when compared with standard laboratory rodents, the NMR immune system manifests some unusual features, such as absence of natural killer cells, higher pro-inflammatory cytokine production by macrophages and a novel neutrophil subset [7]. It is well-established that CRAC channels mediating SOCE are critical for proper functioning of various immune cells [8], such that aberrant SOCE due to abnormal CRAC channel activity is involved in various chronic inflammatory diseases [4] to which NMRs are comparatively more resistant. In addition to the role of CRAC channels in regulating immune cell function, research also indicates that Stim and Orai proteins are expressed in dorsal root ganglion (DRG) sensory neurons and spinal cord dorsal horn neurons, thus suggesting that they play a role in nociceptive processing and pain [6,9]. We, therefore, sought to functionally characterize SOCE and its underlying CRAC channel components in NMR cells to determine whether this pathway exhibits any unusual features that might potentially contribute to its unusual biology.
Material and methods
Cell culture
2.1.
Immortalized fibroblasts from NMR skin (‘NMR SV40 fibroblasts’) were established in the Smith laboratory previously [10] and cultured as per a published protocol. Briefly, cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM) high glucose (Gibco™) supplemented with 15% (v/v) fetal bovine serum (Sigma-Aldrich™), non-essential amino acids, 1 mM sodium pyruvate, 100 units per ml penicillin and 100 mg ml^−1^ streptomycin (Gibco™). Cells were incubated in a humidified 32°C incubator with 5% CO_2_ and 3% O_2_.
NMR DRG neurons (‘NMR-DRG neurons’) were isolated and cultured as per a published protocol [11]. NMR were sacrificed using a rising CO_2_ concentration followed by decapitation in accordance with the UK Animal (Scientific Procedures) Act 1986 Amendment Regulations 2012 under a Project License (PP5814995) granted to E.S.J.S. by the Home Office with additional approval by the University of Cambridge Animal Welfare Ethical Review Body. Before collecting the NMR-DRG neurons, sterile 35 mm glass-bottomed Petri dishes (MatTek, USA) were coated with laminin (20 μg ml^−1^) and incubated at 37℃ for 1 h. Once plated, NMR-DRG neurons were incubated in a humidified 37°C, 5% CO_2_ incubator overnight before Ca^2+^ imaging the following day. Jurkat T cells were cultured as previously described [12].
Ca2+ imaging
2.2.
All Ca^2+^ imaging experiments involving the NMR-SV40 fibroblasts and NMR-DRG neurons were performed at room temperature (21–23°C) using Fura-2-based Ca^2+^ imaging as per a previously published protocol [12]. Paired fluorescence images (340 and 380 nm excitation, 510 nm emission) were captured every 5 s using a QIClick™ digital CCD camera (QImaging, British Columbia, Canada) mounted on a Nikon Eclipse Ti-S Microscope with MetaFluor^®^ (Molecular Devices, USA) Fluorescence Ratio Imaging Software.
Immunocytochemistry and confocal microscopy
2.3.
Jurkat T cells or NMR SV40 fibroblasts plated on Poly-D-Lysine (PDL)-coated glass coverslips were fixed using formaldehyde or methanol, respectively, and permeabilized using Triton X-100 following published protocols [13,14]. Fixed cells were treated overnight at 4°C with polyclonal rabbit Fluorescein Isothiocyanate (FITC)-conjugated antibodies (Cusabio^®^, TX, USA) raised against human Orai, hOrai1 (product code: CSB-PA846601LC01HU) and human Stim1, hStim1 (product code: CSB-PA022829LC01HU) at a final concentration of 25 μg ml^−1^ in phosphate-buffered saline (PBS). After overnight incubation, coverslips were washed with PBS thrice and treated with 1 μg ml^−1^ nuclear stain 4′,6-diamidino-2-phenylindole (DAPI) (Sigma-Aldrich) for 5 min at room temperature. The coverslips were then washed thrice with PBS and mounted onto a microscope slide using ProLong™ Gold Antifade Mountant (ThermoFisher Scientific, UK). Confocal images were acquired using a Leica SP5 confocal microscope equipped with a 63× oil immersion lens (Leica Microsystems, Wetzlar, Germany). The excitation wavelengths used were 405 nm for DAPI and 488 nm for FITC. Acquired images were processed using Leica Application Suite X (LAS X).
Statistical analysis
2.4.
Results were expressed as means ± s.e.m. for independent experiments undertaken over three or more different days. Statistical comparisons of the mean values were performed in GraphPad Prism 9 (GraphPad software Inc., CA, USA) using one-way analysis of variance (ANOVA) followed by Tukey’s multiple comparisons test or Student’s t‐test (unpaired, two-tailed); p < 0.05 was considered to be significant.
Results and discussion
Preliminary characterization of store-operated Ca2+ entry from naked mole-rats SV40 cells
3.1.
We first wanted to determine the likelihood that the protein machineries underpinning SOCE or CRAC channels, namely Orai and Stim, are expressed by NMR cells. Therefore, we looked up the NMR genome database (http://www.naked-mole-rat.org/) as well as UniProt (https://www.uniprot.org/) and identified amino acid sequences representing the putative NMR Orai and Stim isoforms, with significant (>85%) similarities with cognate sequences from mouse, rat and human (electronic supplementary material, S1–S5). Since Orai1 and Stim1 are the major isoforms that underlie native SOCE in most mammalian cells [2], we conducted immunostaining of immortalized NMR skin fibroblasts (NMR SV40 fibroblasts, developed previously in the Smith lab [10]), using FITC-conjugated antibodies raised against human Stim1 (hStim1) and human Orai1 (hOrai1) protein, being able to detect both Stim1 and Orai1 (figure 1A). The fidelities of these antibodies were assessed in Jurkat T cells (electronic supplementary material, figure S6) that are well known to express hStim1 and hOrai1 isoforms and manifest robust SOCE [1,2,12].
Preliminary characterization of SOCE and its underlying protein players in NMR SV40 fibroblasts. (A) Exemplar confocal images showing expression of endogenous Orai and Stim proteins in NMR SV40 fibroblasts. The latter were fixed and stained with FITC-conjugated antibodies raised against hOrai1 and Stim1; nuclei were stained with DAPI and shown in blue. Scale bar, 50 μm. (B) Left, exemplar traces showing Ca2+ signals (indicated by Fura-2 fluorescence ratio) triggered by adding 2 μM of thapsigargin (Tg) to NMR SV40 fibroblasts with or without pre-treatment (10 μM for 10 min) with the indicated known SOCE/CRAC channel inhibitors (shown in different non-green colours). Right, summary data obtained from Ca2+ imaging experiments shown in left, showing peak Ca2+ entry (SOCE) observed in NMR SV40 fibroblasts under different conditions. (C) Left, exemplar traces showing Tg-evoked Ca2+ signals to NMR SV40 fibroblasts with or without pre-treatment (2 or 10 μM, for 10 min) with Pyr6 (known SOCE/CRAC channel inhibitor) and IA65 (known SOCE/CRAC channel enhancer). Right, summary data obtained from Ca2+ imaging experiments shown in left, showing peak Ca2+ entry (SOCE) observed in NMR SV40 fibroblasts under different conditions. Data are mean ± s.e.m. from ≥90 cells (per each experimental condition) plated on three Petri dishes, each imaged on different days. Averages from the experimental groups (i.e. different SOCE-inhibitor treated groups) were compared with that of the control group using one-way ANOVA followed by Tukey’s multiple comparisons test; p < 0.05 was considered to be significant. NMR, naked mole-rats; SOCE, store-operated Ca2+ entry; CRAC, Ca2+ release-activated Ca2+.
After confirming the presence of SOCE components at both genomic and protein levels, we next sought to functionally characterize SOCE in NMR SV40 fibroblasts. For this, we used Fura-2-based Ca^2+^ imaging as per published protocol [12], initially depleting internal stores with 2 µM thapsigargin (Tg) in the absence of extracellular Ca^2+^, followed by addition of Ca^2+^ (as 2 mM CaCl_2_). This protocol induced a robust Ca^2+^-entry rapidly after the addition of Ca^2+^ to the extracellular solution bathing the NMR SV40 fibroblasts (figure 1B). The Ca^2+^ response was significantly attenuated when NMR SV40 fibroblasts were preincubated for 10 min with 10 μM BTP2, GSK-7975A or Pyr6, each being a widely used blocker of SOCE/CRAC channels [3,12] (figure 1B). In a separate series of Ca^2+^ imaging experiments, we used the same protocol and recorded Tg-evoked Ca^2+^ entry in the NMR SV40 fibroblasts pretreated with either low (2 μM) or high (10 μM) concentration of Pyr6, or the recently discovered enhancer of Orai1, namely IA65 [15]. Compared with the untreated cells, Tg-evoked Ca^2+^ entry in Pyr6-pretreated NMR SV40 fibroblasts was significantly attenuated, whereas pretreatment with IA65 resulted in a significantly greater Ca^2+^ response (figure 1B–C). Thus, these two well-characterized SOCE-selective pharmacological tools modulated NMR SOCE in a way similar to how they have been shown to affect SOCE in other mammalian cell types [15]. We noticed that 2 μM Pyr6 sufficed at causing significant suppression of SOCE in NMR SV40 fibroblasts and that 10 μM did not produce any greater attenuation: the peak SOCE responses (indicated by mean Fura-2 ratio) observed for cells pretreated with 2 and 10 μM Pyr6 were comparable (mean ± s.d. = 0.11 ± 0.06 and 0.07 ± 0.04, respectively) with no statistically significant difference (p = 0.122, one-way ANOVA followed by Tukey’s multiple comparison test). Similar was the finding with IA65 as SOCE enhancer; the peak SOCE responses observed for cells pretreated with 2 and 10 μM of this molecule were comparable (mean ± s.d. = 1.06 ± 0.28 and 1.08 ± 0.25, respectively) with no statistically significant difference (p = 0.47, one-way ANOVA followed by Tukey’s multiple comparison test). Thus, in NMR SV40 fibroblasts we were able to detect clear, pharmacologically evoked (i.e. Tg-triggered) SOCE representing active CRAC channels that were sensitive to well-known pharmacological modulators of this pathway.
Next, we aimed to determine whether SOCE could be physiologically evoked in NMR SV40 fibroblasts, i.e through depleting the ER Ca^2+^ store by stimulating cells to activate PLC isoforms and thereby generate the Ca^2+^-mobilizing second messenger, inositol 1,4,5-trisphosphate (IP_3_). Given no a priori knowledge was available regarding the presence of specific Gq-coupled receptors or receptor tyrosine kinases in NMR SV40 fibroblasts, we arbitrarily tested some agonists for cell surface receptors including ATP, endothelin-1 and histamine that are well known to activate PLC and produce IP_3_ in many eukaryotic cell types [16]. However, none of these agonists produced any discernible Ca^2+^ signal in the NMR SV40 fibroblasts (results not shown). In contrast, angiotensin II (Ang II), an endogenously produced octapeptide and a highly potent vasoconstrictor [17], was observed to mobilize Ca^2+^ in NMR SV40 fibroblasts and deplete Ca^2+^ stores, such that Ca^2+^ addition led to robust Ca^2+^ entry via SOCE, as indicated by inhibition by Pyr6 (figure 2). Thus, these NMR fibroblasts are likely to express AT_1_ receptors that are G_q_-coupled G protein-coupled receptors, stimulation of which is known to deplete ER Ca^2+^ stores and trigger SOCE [17,18]. We, however, noticed that Ang II -induced Ca^2+^ release was significantly enhanced in the NMR fibroblasts that were pretreated with Pyr6 (figure 2A,C). The specific reason(s) for this greater Ca^2+^ mobilization upon Pyr6 pretreatment were unclear. Among other potential possibilities, Pyr6 could somehow sensitize component(s) of the relevant Ca^2+^ signal-generating cascade comprising the AT_1_ receptor, PLCβ isoforms and inositol 1,4,5-trisphosphate receptors (IP_3_Rs) endogenous to those cells but this merits future investigation. Since the enhanced Ca^2+^ mobilization was still complete within timeframe comparable to the cells that were not pretreated with Pyr6 (figure 2A), it is conceivable that the ER Ca^2+^ store was emptied properly for activating SOCE that was sensitive to Pyr6 challenge. Altogether the result suggests that NMRs, like other mammals, are able to trigger SOCE physiologically and use this Ca^2+^ entry pathway to conduct various cellular functions, including the housekeeping store refilling purpose [1,2].
Physiologically evoked SOCE in NMR SV40 fibroblasts. (A) Exemplar traces showing Ca2+ signals (indicated by Fura-2 fluorescence ratio) triggered in NMR fibroblasts by adding indicated concentrations of angiotensin II (Ang-II) alone or together with Pyr6 in Ca2+-free extracellular media followed by subsequent Ca2+ add back. (B) Summary data obtained from Ca2+ imaging experiments are shown , showing peak Ca2+ entry (SOCE) observed in NMR SV40 fibroblasts under different conditions. (C) Summary data obtained from Ca2+ imaging experiments shown, showing peak Ca2+ release from the ER observed in NMR SV40 fibroblasts under different conditions. Data are mean ± s.e.m. from ≥75 cells (per each experimental condition) plated on three Petri dishes, each imaged on different days. Averages from the experimental groups (i.e. different SOCE-inhibitor treated groups) were compared with that of the control group using one-way ANOVA followed by Tukey’s multiple comparisons test; p < 0.05 was considered to be significant. NMR, naked mole-rats; SOCE, store-operated Ca2+ entry, ER, endoplasmic reticulum.
Evaluating the effect of exposure to different oxygen levels on store-operated Ca2+ entry in naked mole-rat SV40 fibroblasts
3.2.
It is well documented that NMR fibroblasts thrive under lower O_2_ culturing conditions (3%) than cells from mouse or human, perhaps an adaptation to their subterranean environment and in agreement with their hypoxia resistance [10,14,19]. However, all Ca^2+^ imaging experiments, with or without pre-incubation with various SOCE modulators, were performed at atmospheric O_2_ levels and in Hanks' balanced salt solution (HBSS; figures 1B–C and 2), which might not be optimal for culturing and maintaining the NMR SV40 fibroblasts. Although, NMR SV40 fibroblasts looked healthy during the time period in atmospheric O_2_ and responded well to our Ca^2+^ imaging protocol, we nevertheless sought to explore whether or not changes in the O_2_ level could affect SOCE in these cells. Accordingly, in subsequent experiments, we performed all the pre-Ca^2+^ imaging preparations such as Fura-2AM loading, washing and de-esterification etc., in the NMR SV40 fibroblast maintained in optimal media (cRPMI) and optimal oxygen level (3% O_2_) and compared this to when procedures were conducted in sub-optimal media (HBSS) and optimal oxygen level (3% O_2_) and in sub-optimal media (HBBS) and sub-optimal/atmospheric oxygen level (21% O_2_). Regardless of the pre-Ca^2+^-imaging conditions, we measured SOCE in our usual recording conditions of room temperature and atmospheric O_2_ and in HBSS. When compared with one another, the peak Ca^2+^ entry via SOCE was comparable across different cohorts of NMR SV40 fibroblasts (figure 3), thus suggesting that short-term changes in atmospheric O_2_ did not overtly compromise the function of NMR SV40 fibroblast SOCE.
Evaluation of exposure to different oxygen level on SOCE in NMR SV40 fibroblasts. (A) Exemplar Ca2+ imaging traces showing Tg-evoked SOCE (indicated by Fura-2 fluorescence ratio) in NMR SV40 fibroblasts for which loading of Fura-2AM dye and its subsequent de-esterification was carried out in suboptimal media and optimal O2 level (HBSS, 21% O2), suboptimal media and optimal O2 level (HBSS, 3% O2) and optimal media and optimal O2 level (cRPMI, 21% O2). (B) Summary data obtained from Ca2+ imaging experiments are presented , showing peak Ca2+ entry (SOCE) observed in NMR SV40 fibroblasts under different conditions. Data are mean ± s.e.m. from ≥75 cells (per each experimental condition) plated on three Petri dishes, each imaged on different days. Averages from the experimental groups (i.e. different SOCE-inhibitor treated ones) were compared with that of the control group using one-way ANOVA followed by Tukey’s multiple comparisons test; p < 0.05 was considered to be significant. NMR, naked mole-rats; SOCE, store-operated Ca2+ entry,
Characterizing store-operated Ca2+ entry in primary naked mole-rat neurons
3.3.
Characterization of NMR SOCE in immortalized NMR fibroblasts does not necessarily serve as true representative of NMR SOCE, and we, therefore, sought to characterize SOCE in NMR primary cells. Given the fact that SOCE has been well documented in mammalian DRG neurons [20] and our experience with these cells [11,21], we decided to characterize SOCE in NMR DRG neurons. As in NMR SV40 fibroblasts, we were able to detect robust SOCE both pharmacologically (i.e. using Tg to passively deplete the ER Ca^2+^ store, figure 4A) as well as physiologically (using Ang II to deplete the ER Ca^2+^ store through generating IP_3_, figure 4B), SOCE being significantly suppressed in both cases when pretreated with the SOCE inhibitor Pyr6 (2 μM). At the end of each experiment, 50 mM KCl was added to depolarize the neurons and trigger Ca^2+^ entry via voltage-gated Ca^2+^ channels, only such cells, thus identified as neurons, were analysed for SOCE activity.
SOCE in DRG neurons isolated from the NMRs. (A) Left, exemplar traces showing Ca2+ signals (indicated by Fura-2 fluorescence ratio) triggered in DRG neurones from NMR by adding Tg (2 μM) alone or together with Pyr6 (2 μM) in Ca2+-free extracellular media followed by subsequent Ca2+ add back. Right, summary data obtained from Ca2+ imaging experiments shown in left, showing peak Ca2+ entry (SOCE) observed in DRG neurones from NMR under different conditions. (B) Left, exemplar traces showing Ca2+ signals (indicated by Fura-2 fluorescence ratio) triggered in DRG neurones from NMR by angiotensin II (200 nM) II (Ang-II) alone or together with Pyr6 (2 μM) in Ca2+-free extracellular media followed by subsequent Ca2+ add back. Right, summary data obtained from Ca2+ imaging experiments shown in left, showing peak Ca2+ entry (SOCE) observed in DRG neurones from NMR under different conditions. Data are mean ± s.e.m. from ≥60 DRG neurons (per each experimental condition) plated on three Petri dishes, each imaged on different days. Averages were compared using Student’s t-test, two-tailed, unpaired; p < 0.05 was considered to be significant. NMR, naked mole-rats; SOCE, store-operated Ca2+ entry; DRG, dorsal root ganglion.
Conclusion
In this study, we functionally and pharmacologically characterized SOCE in non-excitable (skin fibroblast) and excitable (DRG neurons) NMR cells, finding that these NMR cells exhibit robust SOCE, which was sensitive to some widely used selective modulators of this pathway, similar to what is observed in human and other mammalian cells. Considering the key role that SOCE plays in a multitude of cellular processes, from refilling depleted intracellular Ca^2+^ stores to gene transcription, cytokine secretion and metabolism [2], it is perhaps not surprising that SOCE activity appears normal in NMR cells, but this was still worthy of investigation considering the unusual physiology of NMRs [6]. What remains to be determined is whether the SOCE measured was reliant mainly on Orai1 and Stim1 proteins as is the case in most other mammalian cell types, but the expression of Orai1 and Stim1 in NMR SV40 fibroblasts indicates that they are likely involved. In addition, the potential contributions and physiological roles of Stim2 and Orai 2 and Orai3 isoforms also need to be investigated. Future studies may also include cloning and functional characterization of individual Stim and Orai isoforms, determining their fundamental biophysical properties and sensitivity to pharmacological modulators.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Prakriya M, Lewis RS. 2015 Store-operated calcium channels. Physiol. Rev. 95, 1383–1436. (10.1152/physrev.00020.2014)26400989 PMC 4600950 · doi ↗ · pubmed ↗
- 2Emrich SM, Yoast RE, Trebak M. 2022 Physiological functions of CRAC channels. Annu. Rev. Physiol. 84, 355–379. (10.1146/annurev-physiol-052521-013426)34637326 · doi ↗ · pubmed ↗
- 3Rubaiy HN. 2023 ORAI calcium channels: regulation, function, pharmacology, and therapeutic targets. Pharmaceuticals 16, 162. (10.3390/ph 16020162)37259313 PMC 9967976 · doi ↗ · pubmed ↗
- 4Feske S. 2019 CRAC channels and disease—from human CRAC channelopathies and animal models to novel drugs. Cell Calcium 80, 112–116. (10.1016/j.ceca.2019.03.004)31009822 PMC 6545165 · doi ↗ · pubmed ↗
- 5Stauderman KA. 2018 CRAC channels as targets for drug discovery and development. Cell Calcium 74, 147–159. (10.1016/j.ceca.2018.07.005)30075400 · doi ↗ · pubmed ↗
- 6Buffenstein R et al. 2022 The naked truth: a comprehensive clarification and classification of current ‘myths’ in naked mole-rat biology. Biol. Rev. Camb. Philos. Soc. 97, 115–140. (10.1111/brv.12791)34476892 PMC 9277573 · doi ↗ · pubmed ↗
- 7Lin T, Buffenstein R. 2021 The unusual immune system of the naked mole-rat. Adv. Exp. Med. Biol. 1319, 315–327. (10.1007/978-3-030-65943-1_12)34424522 · doi ↗ · pubmed ↗
- 8Feske S, Wulff H, Skolnik EY. 2015 Ion channels in innate and adaptive immunity. Annu. Rev. Immunol. 33, 291–353. (10.1146/annurev-immunol-032414-112212)25861976 PMC 4822408 · doi ↗ · pubmed ↗