Lung surfactant reduces Staphylococcus aureus cytotoxicity and protects host immune cells from membrane damage
Maria Predtechenskaya, Corbin J. Arbizzani, Sofia R. Shomento, Timothy R. Borgogna, Jovanka M. Voyich

TL;DR
Lung surfactant protects immune cells from Staphylococcus aureus toxins by reducing their membrane-damaging effects and suppressing virulence gene expression.
Contribution
This study reveals a novel protective role of lung surfactant in modulating S. aureus virulence during lung infections.
Findings
Natural surfactants from mice and rats reduce S. aureus toxin-induced membrane damage in human immune cells.
Mouse and rat surfactants downregulate virulence gene expression in S. aureus, including saeR, lukF-PV, hla, and hlgA.
Commercial surfactant Infasurf does not suppress S. aureus virulence gene transcription like natural surfactants.
Abstract
In this study, we identify that lung surfactant significantly reduces the cytotoxicity of Staphylococcus aureus (S. aureus) membrane-damaging toxins. Data demonstrate that natural surfactants from mice and rats and commercially available surfactant, Infasurf, protect human primary cells (neutrophils and peripheral blood mononuclear cells) from cytolytic activity caused by S. aureus supernatants. Supernatants from S. aureus grown in surfactant showed a significant reduction in plasma membrane damage against primary human cells as compared to supernatants grown without surfactant. This reduction was not due to a direct bactericidal effect of the surfactants on S. aureus growth. Rat and mouse surfactants downregulated the gene expression of saeR, the response regulator of the S. aureus two-component system SaeR/S that is responsible for the production of virulence factors which are…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
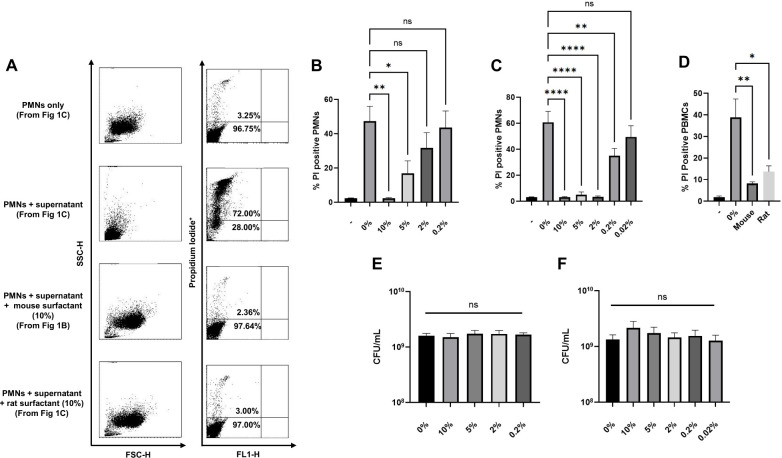 Fig 1
Fig 1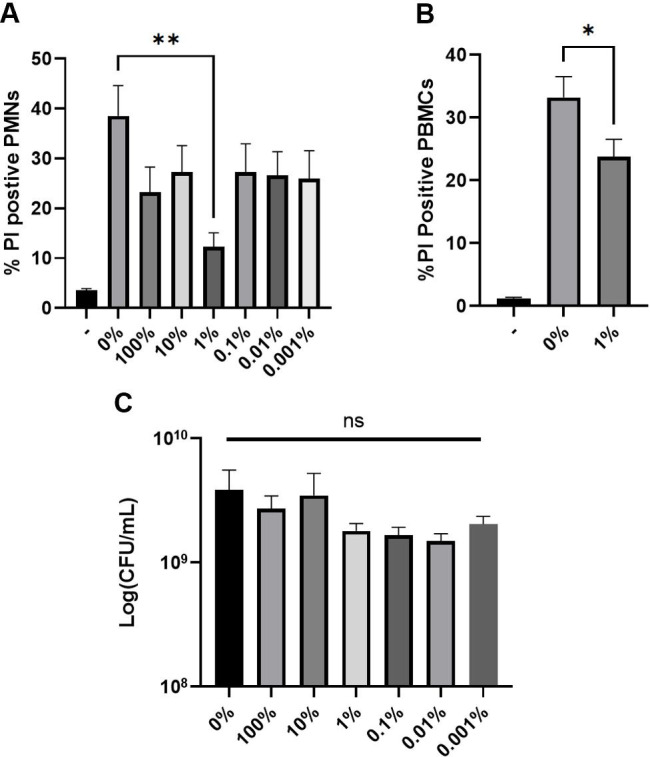 Fig 2
Fig 2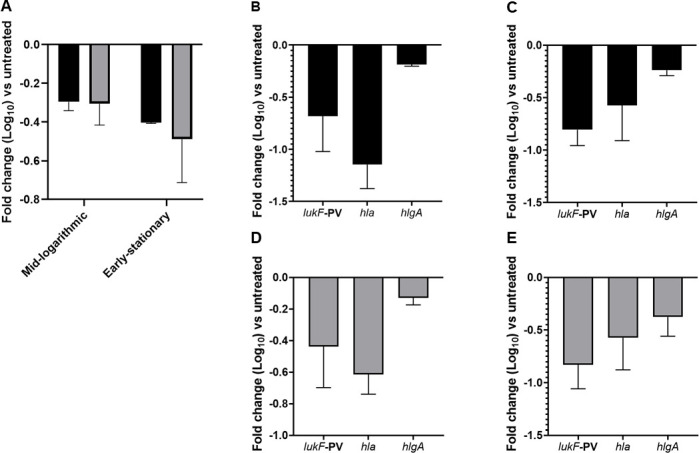 Fig 3
Fig 3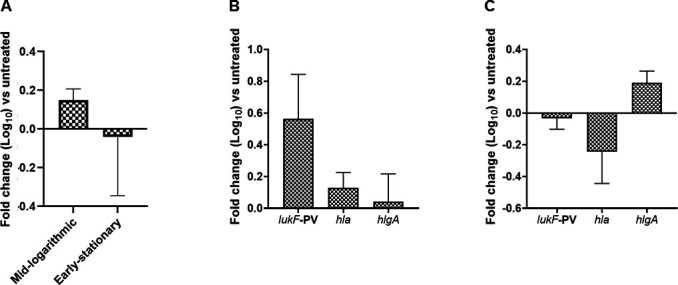 Fig 4
Fig 4- —HHS | National Institutes of Health (NIH)
- —HHS | National Institutes of Health (NIH)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntimicrobial Resistance in Staphylococcus · Immune Response and Inflammation · Antimicrobial Peptides and Activities
INTRODUCTION
Pulmonary surfactant is a lipid-rich complex within the alveoli that prevents atelectasis during respiration by maintaining surface tension at the air-liquid interface (1, 2). Lung surfactant is predominantly produced by alveolar type II epithelial cells and is composed of 90% lipids, of which dipalmitoylphosphatidylcholine (DPPC) is the most abundant (3). The remaining 10% are proteins (1–4). There are four surfactant proteins, SP-A, SP-B, SP-C, and SP-D, which have been demonstrated to be essential for the structure and functionality of surfactants (2, 5, 6). SP-A and SP-D lung surfactant proteins are also opsonins that assist phagocytosis of bacterial pathogens by alveolar macrophages and neutrophils (7). SP-A and SP-D have also been shown to increase membrane permeability in Gram-negative bacteria leading to lysis (8).
Lung surfactant components have additional immunomodulatory properties, including anti-angiogenesis activity (9) and inhibition of the generation of reactive oxygen intermediates in neutrophils and monocytes (10). Vesicles containing lung surfactant components, like DPPC, have been shown to decrease macrophage inflammation by inducing expression of innate immune receptors, including Fc receptors, CD11b, scavenger and mannose receptors, and complement receptor CR1 or interfering with toll-like receptor-mediated inflammatory responses (3). It follows that aberrant levels of lung surfactant have been associated with many respiratory problems, among which are pulmonary fibrosis, cystic fibrosis, and chronic obstructive pulmonary disease (COPD) (3), as well as during infection, for example, influenza A virus (11). During lower respiratory infections with influenza A virus, alveolar type II cells are preferentially targeted (12). Infection of these cells leads to measurable disruptions in surfactant production (13).
In a previous study, we identified that antecedent influenza A virus infection in mice increased virulence gene expression in S. aureus compared to virulence gene expression during S. aureus lung infection only (14). The same study demonstrated that secondary S. aureus pneumonia following influenza A viral infection was SaeR/S-dependent (14). These observations combined with previous observations of the natural properties of surfactant provided the basis for the current study to investigate whether surfactant may directly impact virulence gene expression and cytotoxicity of S. aureus. Our results demonstrate that the presence of natural as well as commercially available lung surfactants prevent membrane damage in neutrophils and peripheral blood mononuclear cells by S. aureus toxins. Murine and rat lung surfactants repressed transcription of S. aureus secreted toxins, whereas commercially available surfactants did not decrease transcripts of virulence genes tested. This work adds to our knowledge of the role of surfactants in healthy lungs and implies that lung surfactant contributes to reducing the pathogenicity of bacterial infections.
MATERIALS AND METHODS
Bacterial strains and culture conditions
Staphylococcus aureus (S. aureus) PFGE-type USA300 strain LAC (15) was used in all experiments. Unless noted otherwise, overnight and sub-cultured bacteria (1:100 dilution of overnight) were grown in 20 mL of tryptic soy broth (TSB; EMD Millipore) supplemented with 0.5% glucose with shaking (250 rpm) at 37°C. Optical density at 600 nm (OD_600_) was measured using a NanoDrop 2000c Spectrophotometer (ThermoFisher Scientific), and colony-forming units (CFUs) were determined by plating diluted samples on tryptic soy agar (TSA; EMD Millipore) and enumerated the following day.
Lung surfactant extraction and growth assays
Pulmonary surfactant isolation was performed using an adaptation of the method described in Inselman et al. (16). Briefly, murine (C57BL/6) or rat (F344BN) lung tissue (0.2 g) was homogenized in 3 mL of ice-cold Dulbecco’s phosphate-buffered saline (DPBS) using a tissue grinder. Homogenate slurry was passed through a 70 µm cell strainer followed by centrifugation at 300 × g for 10 minutes at 4°C. The supernatant was collected and placed in microcentrifuge tubes for centrifugation at 18,000 × g for 30 minutes at 4°C. The resulting supernatant was aspirated and discarded. The remaining surfactant pellet was resuspended in 1 mL of DPBS at 60°C. Research-grade Infasurf was purchased from Onybiotech. For experiments investigating growth in surfactant, 1 mL of surfactant (at varied concentrations) was added to 4 mL of TSB pre-warmed to 37°C and inoculated with 50 µL of overnight culture. For experiments, adding 1 mL of the original surfactant pellet to 4 mL of TSB was considered 100% surfactant. S. aureus cultures were incubated with surfactant for 5 hours at 37°C with shaking (250 rpm). Heat inactivation of proteins was performed by incubating surfactants for 30 minutes at 56°C (17).
Relative quantitative real-time RT-PCR
Transcription of S. aureus genes was assessed using TaqMan real-time reverse transcriptase-PCR (RT-PCR) as previously described (18–20). Briefly, sub-cultured strains were harvested at mid-exponential (ME; OD_600_ = 1.5) or early stationary (ES; OD_600_ = 3.0) phase of growth, mechanically disrupted using a FastPrep FP120 cell disrupter (ThermoFisher Scientific), and RNA purified using RNeasy Kit (Qiagen) as described in reference 18. TaqMan real-time RT-PCR was performed using primer and probe sets as published previously (14) and analyzed with the delta-delta Ct method.
Human PMN or PBMC plasma membrane integrity assays
Heparinized venous blood from healthy donors was collected in accordance with a protocol approved by the Institutional Review Board for Human Subjects at Montana State University. All donors provided written consent to participate in the study. Human polymorphonuclear leukocytes (neutrophils or PMNs) and/or peripheral blood mononuclear cells (PBMCs) were isolated under endotoxin-free conditions (<25.0 pg/mL) and cell viability and purity of preparations were assessed using a FACSCalibur Flow cytometer (BD Biosciences) as described in references 18 and 21. Assays of intoxicating PMNs with extracellular S. aureus proteins were performed as previously described (22, 23). Briefly, supernatants from S. aureus sub-cultured for 5 h in TSB with glucose were sterile-filtered (0.22 µm, Avantor) and diluted in DPBS. PMNs or PBMCs (1 × 10^6^) were exposed to varied dilutions of S. aureus supernatant. Samples were incubated at 37°C for 60 min then stained with propidium iodide (PI; ThermoFisher Scientific) following the manufacturer’s protocol and then analyzed by FACS as in reference 23.
RESULTS
Surfactants from mouse and rat lungs protect immune cells from S. aureus toxin-mediated membrane damage
To test the hypothesis that surfactant influences S. aureus virulence, we investigated the role of surfactant on S. aureus toxin cytotoxicity. In these assays, we compared plasma membrane damage in human neutrophils (PMNs) and peripheral blood mononuclear cells (PBMCs) following exposure to S. aureus supernatants grown in the presence or absence of varied concentrations of mouse or rat lung surfactant (Fig. 1A through D). PMNs exposed to supernatants grown in 10% mouse surfactant demonstrated a significant reduction in plasma membrane damage and had an average of 2.39% ± 0.39% propidium iodide (PI)-positive cells compared to 47.39% ± 8.63% PI-positive cells from supernatants grown in TSB only (Fig. 1B). S. aureus grown in rat surfactant also demonstrated significantly reduced plasma membrane damage, for example, 3.17% ± 0.33% PI positive in 10% rat surfactant compared to PMNs exposed to supernatants from S. aureus grown in TSB only at 60.67% ± 8.44% PI-positive (Fig. 1C). However, unlike mouse surfactant, rat surfactant maintained its ability to significantly reduce plasma membrane damage at a concentration as low as 0.2%. Similarly, growth in mouse or rat surfactants significantly reduced PBMC plasma membrane damage from S. aureus supernatants. PBMCs exposed to supernatants harvested from S. aureus grown in 10% mouse surfactant or 2% rat surfactant were 8.26% ± 0.72% and 13.70% ± 2.68% PI-positive, respectively, compared to control at 38.90% ± 8.50% PI-positive (Fig. 1D). The reduction of membrane damage in cells was not due to decreased bacterial growth in the presence of surfactants, and no significant differences in bacterial growth were seen in cultures grown with or without surfactant (Fig. 1E and F).
*Surfactant from mouse and rat lungs protects human primary cells from S. aureus-mediated cytotoxicity. (A) Representative flow cytometry plots of data displayed in (B) and (C). S. aureus was grown to the early stationary phase in TSB with varied dilutions of (B) mouse or (C) rat lung surfactant. S. aureus supernatants were harvested, diluted to 1:50 final concentration, and incubated with human PMNs for 1 hour. Plasma membrane damage was assessed using propidium iodide (PI) uptake and analyzed by flow cytometry. (D) Human PBMCs exposed to S. aureus supernatants diluted to 1:5 final concentration following growth in 10% mouse or 2% rat surfactant (concentration determined in B and C), and plasma membrane damage assessed with PI. S. aureus CFUs collected after growth in mouse (E) or rat surfactants (F). Dash (-) represents cells without exposure to supernatants or surfactants, while 0% represents cells with exposure to only supernatants. Data are from three biological replicates for (B), (D), and (E), and six biological replicates for (C) and (F). *P < 0.05, **P < 0.005, and ***P < 0.0001 one-way ANOVA followed by Dunnett’s multiple comparison test. NS = not significant.
Commercially available surfactant Infasurf protects immune cells from S. aureus toxin-mediated membrane damage
We next assessed whether this same protective effect could be observed with commercially available surfactants. For these experiments, S. aureus was grown in varied concentrations of Infasurf. As above, supernatants were harvested and plasma membrane damage in PMNs and PBMCs was assessed. Only growth in 1% Infasurf provided significantly reduced PI uptake by PMNs. Growth in 1% Infasurf yielded an average of 12.31% ± 2.79% propidium iodide (PI)-positive cells, statistically decreased compared to 38.53% ± 6.05% PI-positive cells from supernatants grown in TSB only (Fig. 2A). This concentration of Infasurf also reduced plasma membrane damage of PBMCs. Without surfactant, PBMCs were 33.21% ± 3.31% PI-positive when exposed to supernatants versus 23.80% ± 2.75% PI-positive with 1% Infasurf (Fig. 2B). As with mouse and rat surfactants, growth in Infasurf did not impact bacterial viability (Fig. 2C). We originally expected that Infasurf would mirror natural surfactants in that higher concentrations would be able to protect immune cells in a dose-dependent manner. Surprisingly, higher Infasurf concentrations no longer protected PMNs from membrane damage from S. aureus supernatants (Fig. 2A). Since a previous report of another commercially available surfactant, Surfactant TA observed changes in PMNs consistent with apoptosis (24), we investigated if higher concentrations of Infasurf impacted PMN plasma membrane damage. Incubating PMNs with higher concentrations of Infasurf (without any S. aureus supernatants) did not increase PI uptake in PMNs over controls (Fig. S1).
*Commercially available surfactant Infasurf protects human primary cells from S. aureus-mediated cytotoxicity. (A) S. aureus was grown to early stationary phase in TSB with varied dilutions of Infasurf and S. aureus supernatants were harvested, diluted to 1:50 final dilution, and incubated with human PMNs for 1 hour. Plasma membrane damage was assessed by PI staining and flow cytometry. (B) Human peripheral blood mononuclear cells were exposed to S. aureus supernatants harvested as in (A) following growth in 1% Infasurf (determined in A), and plasma membrane damage was assessed. (C) S. aureus CFUs collected after growth in Infasurf. Dash (-) represents cells without exposure to supernatants or surfactants while 0% represents cells with exposure to only supernatants. Data are from five biological replicates for (A) and (C), and eight biological replicates for (B). *P < 0.05 and *P < 0.005 one-way ANOVA followed by Dunnett’s multiple comparison test for (A), (B), and (C). NS = not significant.
Mouse and rat lung surfactants modulate S. aureus gene expression
The SaeR/S system is a two-component system of S. aureus responsible for controlling virulence gene expression (23, 25–27). In addition, we and others have previously identified a role for SaeR/S in S. aureus lung infections (14, 26). In the current study, S. aureus was grown to mid-logarithmic and early stationary phases of growth with or without surfactant. The relative fold decrease in saeR compared to control was: −0.29 ± 0.04 for mouse surfactant and −0.30 ± 0.11 for rat surfactant at mid-logarithmic phase (Fig. 3A shown in log scale). The transcript was also reduced at the early stationary phase (−0.40 ± 0.01 for mouse surfactant and −0.48 ± 0.22 for rat surfactant (Fig. 3A).
Lung surfactants from mice and rats impact transcription of S. aureus saeR and decrease transcription of S. aureus virulence genes. (A) S. aureus was grown in mouse (black) or rat (gray) surfactant to mid-logarithmic or early stationary phase. RNA was harvested and subjected to TaqMan RT-PCR. Gene transcripts were normalized to gyrB. Data shown are the mean fold-change of S. aureus saeR relative to treatment with S. aureus only. For B-E, S. aureus was grown in mouse (B and C, black bars) or rat surfactant (D and E, gray bars) to the mid-logarithmic phase (B and D) or the early stationary phase (C and E) of growth. RNA was harvested as in A. Data shown are the mean fold-change of the indicated gene relative to treatment with S. aureus only. Error bars indicate the mean ± SEM of three biological replicates for each surfactant tested.
We next assessed the transcript abundance of lukF-PV, hla, and hlgA, three S. aureus virulence genes directly regulated by SaeR (21) and demonstrated to be differentially regulated during murine lung infections (14). Compared to control treatment with DPBS, we observed decreases in the abundance of transcripts encoding various toxins when S. aureus was grown to either mid-logarithmic or early stationary phase with mouse and rat surfactants (Fig. 3B through E shown in log scale). At mid-logarithmic phase, the relative fold decrease in lukF-PV, hla, and hlgA compared to control for mouse surfactant was: −0.68 ± 0.33,–1.14 ± 0.23,–0.18 ± 0.01, respectively (Fig. 3B). Similarly, the relative fold decrease in lukF-PV, hla, and hlgA compared to control for rat surfactant was: −0.43 ± 0.26, –0.61 ± 0.12, and –0.12 ± 0.04 (Fig. 3D). At early stationary phase, the relative fold decrease in lukF-PV, hla, and hlgA compared to control for mouse surfactant was: −0.80 ± 0.15, –0.57 ± 0.33, –0.23 ± 0.05 (Fig. 3C). For rat surfactant, the relative fold decrease in lukF-PV, hla, and hlgA compared to control was: −0.83 ± 0.22, –0.57 ± 0.30, –0.37 ± 0.18 (Fig. 3E).
Infasurf modulates gene expression in S. aureus but patterns are distinct from natural surfactants
Next, we investigated whether modulation of gene transcription occurred in S. aureus following growth in Infasurf (Fig. 4A through C shown in log scale). Compared to natural surfactants, which had a stable repression of saeR in both mid-logarithmic and early stationary phases, commercially available surfactants had a moderate upregulation of saeR during the mid-logarithmic phase and a weak downregulation by the early stationary phase. Following growth in 1% Infasurf, the relative fold change of saeR compared to control was as follows: +0.14 ± 0.05 in the mid-logarithmic phase and −0.04 ± 0.30 in the early stationary phase (Fig. 4A). In contrast to the results seen with natural surfactant, the presence of Infasurf upregulated S. aureus virulence gene transcripts by mid-logarithmic phase. When grown in Infasurf to mid-logarithmic phase, lukF-PV, hla, and hlgA expression were upregulated compared to control by +0.56 ± 0.28, +0.12 ± 0.09, +0.04 ± 0.17, respectively (Fig. 4B). This upregulation of toxin genes during mid-logarithmic phase was consistent with the upregulation of saeR observed following growth in Infasurf to mid-logarithmic phase. When grown in Infasurf to early stationary phase, there was a slight reduction in lukF-PV and hla compared to control: −0.03 ± 0.06 and –0.24 ± 0.20 (Fig. 4C). However, hlgA was increased in the presence of surfactant (+0.19 ± 0.07) (Fig. 4C).
Commercially available surfactant Infasurf impacts transcription of S. aureus saeR and S. aureus virulence genes. (A) S. aureus was grown in Infasurf to mid-logarithmic or early stationary phase. RNA was harvested and subjected to TaqMan RT-PCR. Gene transcripts were normalized to gyrB. (A) Data shown are the mean fold-change of S. aureus saeR relative to treatment with S. aureus only. S. aureus was grown to mid-logarithmic (B) or early stationary phase (C) of growth in TSB with 1% Infasurf. RNA was harvested as in A. Data shown are the mean fold-change of the indicated gene relative to treatment with S. aureus only. Error bars indicate the mean ± SEM of three biological replicates for each surfactant tested.
DISCUSSION
This study reports an interesting observation that natural lung surfactants protect host immune cells from damage from S. aureus-secreted toxins. We demonstrated that when S. aureus is grown in the presence of mouse and rat surfactants, there was a significant decrease in the membrane-damaging ability of secreted S. aureus toxins against human PMNs and PBMCs (Fig. 1). To investigate whether the decrease in cytotoxicity in PMNs and PBMCs was regulated at the level of transcription, we assessed gene expression of saeR, the response regulator of the two-component gene regulatory system SaeR/S. SaeR/S regulates numerous adhesins, toxins, and immunomodulatory proteins important in neutrophil evasion (28) and is important in mouse models of S. aureus lung infections (14, 26). We have previously demonstrated a major role for SaeR/S in the secretion of factors that are cytolytic toward human PMNs (23). In addition, we showed that secondary S. aureus pneumonia following influenza A viral infection is SaeR/S-dependent (14). In the current study, we demonstrated that growth of S. aureus to mid-logarithmic and early stationary phases in the presence of mouse and rat surfactants decreased saeR transcript abundance (Fig. 3). Genes regulated by SaeR/S that encode proteins associated with plasma membrane damage (29) and associated with S. aureus lung infections (14, 26) were also downregulated, suggesting that surfactants regulate toxin production at the level of gene transcription. SaeR/S is instrumental in regulating virulence transcripts during the growth phases we tested (19, 20). However, SaeR/S may not be the only two-component system modulated by surfactant. Future studies will investigate other regulatory systems known to regulate toxin production, such as Agr. Agr is well known to impact toxin production in S. aureus (30) and understanding its role in response to exposure to surfactant will advance our understanding of how surfactant is modulating S. aureus virulence.
Our observations with Infasurf were inconclusive. While we initially expected to find that commercially available surfactant Infasurf would be equally as protective of immune cells as natural surfactants, this was not the case. Protection of cells from membrane damage by cytolytic toxins was only observed when supernatants were harvested from S. aureus after growth in 1% Infasurf (Fig. 2). Supernatants harvested following growth in Infasurf in concentrations over 1% increased membrane damage in cells, and protection was rapidly lost at dilutions below 1%. However, this was not due to a direct effect of Infasurf on membrane integrity (Fig. S1).
Results analyzing virulence transcripts following growth in Infasurf were similarly inconclusive and demonstrated no clear pattern of suppression or activation of the genes investigated. Unlike mouse and rat surfactants, Infasurf did not decrease the transcription of saeR or associated virulence factors. Similar to our results, a report published by Ishii et al. tested another commercially available surfactant, Surfacten, which is derived from bovine lungs like Infasurf (31). Although there were differences in experimental conditions, including strain used, growth conditions, and assay used for transcript analysis, the study demonstrated that lukF-PV, hla, and hlgA were not significantly influenced by the surfactant at the late logarithmic phase (31). We conclude that Infasurf may be protecting cells through another mechanism or perhaps the timing of when it impacts virulence gene expression is different than that observed with the mouse and rat surfactants. Of note, at the time of this study, we were unable to obtain other commercially available surfactants for research purposes. However, future studies will investigate additional commercially available surfactants to see whether they may behave more similarly to natural surfactants.
One explanation behind the observed variability in the influence of natural and commercially available surfactants on S. aureus virulence gene expression and toxin production may be due to differences in surfactant composition, whether this is from different species or varying concentrations. For example, it is known that rat surfactant has a higher phosphatidylcholine concentration than mouse surfactant, both of which are different in composition from bovine surfactant (32). Specific surfactant lipids that have been shown to attenuate inflammation and alter the host response (33) may be at different concentrations in these surfactants. Besides lipid components, there are reports that lung surfactant proteins SP-A and SP-D are involved in pathogen opsonization (3, 5), puncturing microbial membranes (3, 8), suppressing microbial growth, aiding in detoxifying bacterial LPS, and modulating phagocytosis and inflammatory responses by alveolar macrophages (3, 4). SP-D has also been shown to be able to bind to immune cell receptors, modulate complement activation, and enhance bacterial phagocytosis (5). We performed a preliminary screen to identify if proteins contributed to the protective effect we observed in this study. Heat-inactivated mouse and lung surfactants were as protective as untreated surfactants (Fig. S2). Results from our data using heat-inactivated surfactant suggest a lipid component is responsible for our observations. Ishii et al. also suggested that lipid components in surfactants like palmitate can affect virulence expression of S. aureus (31). The authors of that study suggest surfactant components may cause membrane stress, triggering S. aureus virulence gene expression through the stress response regulator SigB (31).
Others have shown that surfactant components, including SP-A, SP-D (8), and free fatty acids (34), can be bactericidal. However, our findings were not due to surfactant directly impacting S. aureus growth, highlighting that another mechanism is influencing the observed decrease in virulence. Our data suggest that modulation of gene expression by natural surfactants reduces S. aureus cytotoxicity. Published observations demonstrate that an exposure of S. aureus to an overabundance of fatty acids (e.g., oleic, sapienic, myristic, palmitic, myrsitelaidic, lauric acids, and more) shuts down the SaeR/S system (35, 36) support our findings. S. aureus has evolved strategies to incorporate and protect itself from host fatty acids through lipases and the incorporation of exogenous fatty acids that alter the S. aureus lipid membrane (37–39). The presence of exogenous fatty acids has also been reported to impact SaeR/S activity (40–43). Future studies will investigate the differences in the composition of surfactants to determine the effectors responsible for our observations of reduced virulence gene expression and cytotoxicity following growth or exposure to surfactants.
In summary, this study identified a putative role for surfactants in the protection of the lung from S. aureus infection. Our data suggest that the surfactants may provide a first line of defense in healthy lungs against S. aureus infection. This would provide a logical explanation for the observation that despite the high number of individuals who are colonized with S. aureus in the nares (28, 44), providing frequent opportunity for exposure to the lungs due to natural aspiration, primary S. aureus lung infections are not common. We hypothesize that surfactant suppresses S. aureus virulence so the pathogen can be cleared with innate immune defenses. It follows that disruption of surfactant abundance or composition may predispose individuals to S. aureus lung infection as occurs following influenza A virus infections (14). Future studies will be conducted to test these hypotheses and identify additional mechanisms behind the influence of surfactants on S. aureus virulence.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Veldhuizen EJA, Haagsman HP. 2000. Role of pulmonary surfactant components in surface film formation and dynamics. Biochimica et Biophysica Acta (BBA) - Biomembranes 1467:255–270. doi:10.1016/S 0005-2736(00)00256-X 11030586 · doi ↗ · pubmed ↗
- 2Huck B, Hidalgo A, Waldow F, Schwudke D, Gaede KI, Feldmann C, Carius P, Autilio C, Pérez-Gil J, Schwarzkopf K, Murgia X, Loretz B, Lehr C-M. 2021. Systematic analysis of composition, interfacial performance and effects of pulmonary surfactant preparations on cellular uptake and cytotoxicity of aerosolized nanomaterials. Small Science 1:12. doi:10.1002/smsc.202100067 PMC 1193602140212853 · doi ↗ · pubmed ↗
- 3Chroneos ZC, Sever-Chroneos Z, Shepherd VL. 2010. Pulmonary surfactant: an immunological perspective. Cell Physiol Biochem 25:13–26. doi:10.1159/00027204720054141 PMC 3025886 · doi ↗ · pubmed ↗
- 4Kishore U, Greenhough TJ, Waters P, Shrive AK, Ghai R, Kamran MF, Bernal AL, Reid KBM, Madan T, Chakraborty T. 2006. Surfactant proteins SP-A and SP-D: structure, function and receptors. Mol Immunol 43:1293–1315. doi:10.1016/j.molimm.2005.08.00416213021 · doi ↗ · pubmed ↗
- 5Zhang Z, Abdel-Razek O, Hawgood S, Wang G. 2015. Protective role of surfactant protein D in ocular Staphylococcus aureus infection. P Lo S ONE 10:e 0138597. doi:10.1371/journal.pone.013859726398197 PMC 4580580 · doi ↗ · pubmed ↗
- 6Ding J, Takamoto DY, von Nahmen A, Lipp MM, Lee KY, Waring AJ, Zasadzinski JA. 2001. Effects of lung surfactant proteins, SP-B and SP-C, and palmitic acid on monolayer stability. Biophys J 80:2262–2272. doi:10.1016/S 0006-3495(01)76198-X 11325728 PMC 1301417 · doi ↗ · pubmed ↗
- 7Mc Cormack FX, Whitsett JA. 2002. The pulmonary collectins, SP-A and SP-D, orchestrate innate immunity in the lung. J Clin Invest 109:707–712. doi:10.1172/JCI 1529311901176 PMC 150919 · doi ↗ · pubmed ↗
- 8Wu H, Kuzmenko A, Wan S, Schaffer L, Weiss A, Fisher JH, Kim KS, Mc Cormack FX. 2003. Surfactant proteins A and D inhibit the growth of Gram-negative bacteria by increasing membrane permeability. J Clin Invest 111:1589–1602. doi:10.1172/JCI 1688912750409 PMC 155045 · doi ↗ · pubmed ↗