ClpB enhances thermotolerance in Campylobacter jejuni through protein disaggregation independent of DnaK
Jeong In Hur, Jinshil Kim, Sangryeol Ryu, Byeonghwa Jeon

TL;DR
This study shows that ClpB helps Campylobacter jejuni survive heat stress by breaking up protein clumps, which is important for food safety.
Contribution
ClpB enhances thermotolerance in C. jejuni through protein disaggregation, independent of DnaK.
Findings
ClpB-deficient mutants show significantly lower survival during heat treatment.
ClpB from heat-tolerant strains has unique amino acid substitutions in the nucleotide-binding domain.
ClpB does not bind to DnaK in C. jejuni, indicating a distinct protein disaggregation mechanism.
Abstract
Campylobacter jejuni is a leading cause of foodborne infections worldwide and primarily transmitted to humans through the consumption of contaminated poultry meat. To enhance Campylobacter-associated food safety, it is critical to understand how C. jejuni survives during the thermal processing of poultry products. In this study, we monitored the survival of 86 C. jejuni strains during heat treatment and observed that some strains exhibited elevated heat tolerance. Notably, multilocus sequence typing clonal complex (CC)-443 and CC-607 were dominant among heat-tolerant strains, while the CC-21 strains were mostly heat-sensitive, indicating phylogenetic association with thermotolerance. We also investigated the function of heat shock chaperones in the thermotolerance of C. jejuni. Among several knockout mutants of heat shock chaperones, a mutant lacking clpB exhibited significantly lower…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
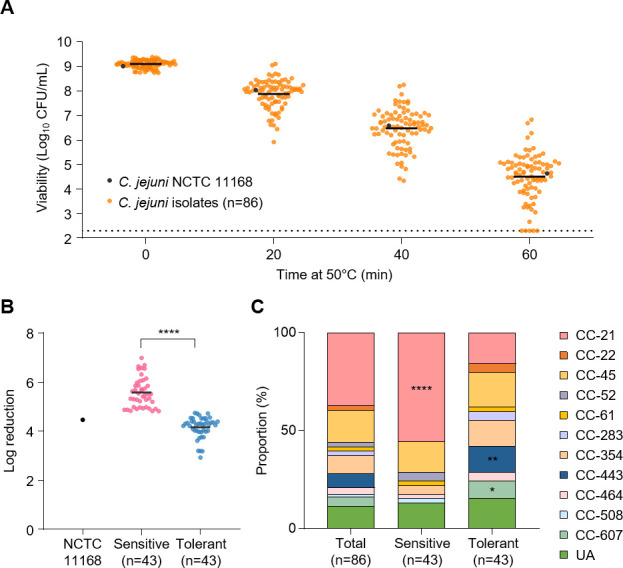 Fig 1
Fig 1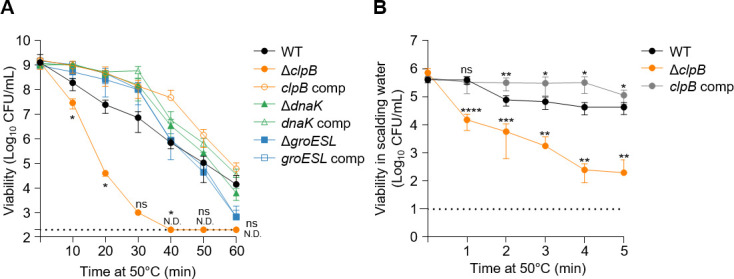 Fig 2
Fig 2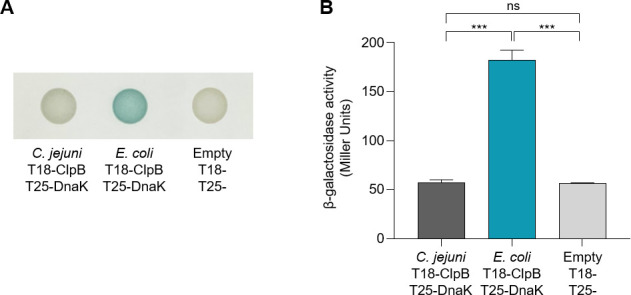 Fig 3
Fig 3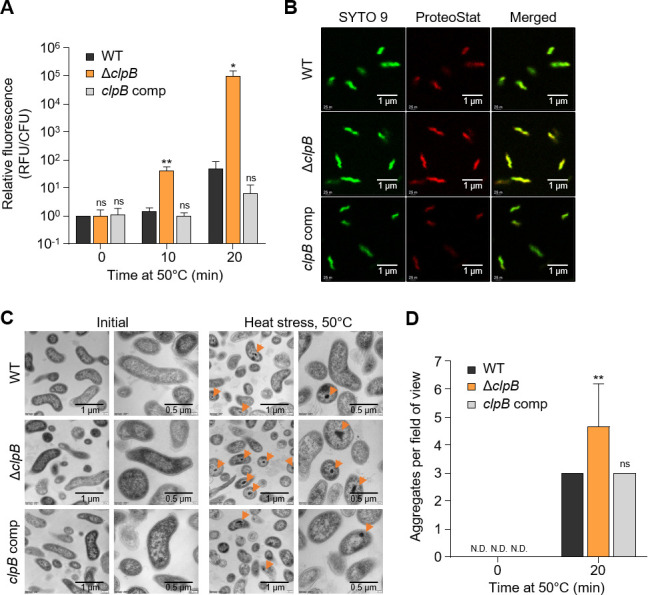 Fig 4
Fig 4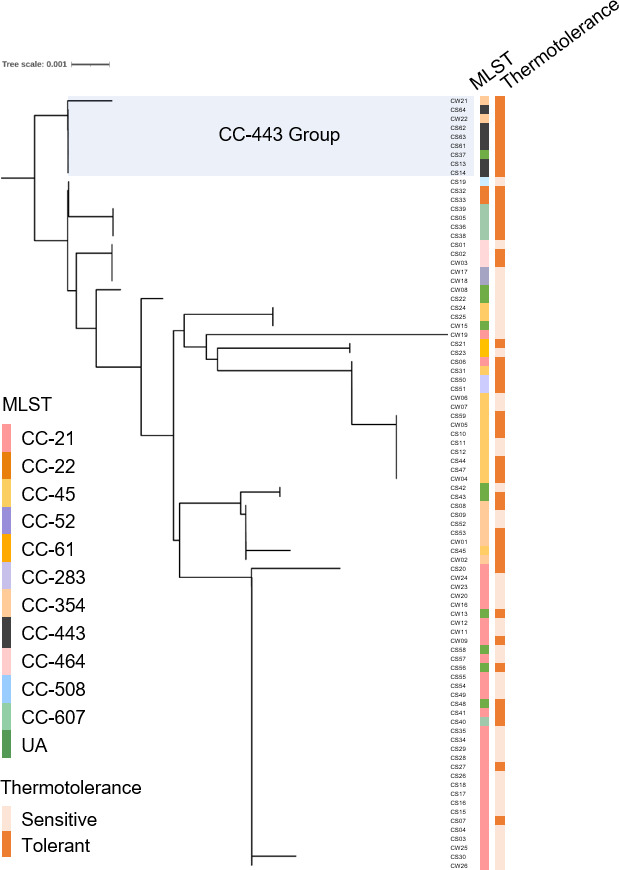 Fig 5
Fig 5| Initial population | WT | Δ | ||||
|---|---|---|---|---|---|---|
| (CFU/mL) | R2 | R2 |
| |||
| 1 × 109 | 12.45 | 0.96 | 4.79 ± 0.01 | 0.9877 | 13.46 ± 0.01 | 0.8576 |
- —National Research Foundation of Koreahttp://dx.doi.org/10.13039/501100003725
- —National Research Foundation of Koreahttp://dx.doi.org/10.13039/501100003725
- —Minnesota's Discovery, Research, and Innovation Economy
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSalmonella and Campylobacter epidemiology · Viral gastroenteritis research and epidemiology · Listeria monocytogenes in Food Safety
INTRODUCTION
Campylobacter jejuni is a predominant bacterial cause of foodborne illnesses globally (1, 2) and primarily transmitted through contaminated poultry products (3, 4). Symptoms of infection in humans include diarrhea, abdominal pains, and fever, with potential severe complications, such as Guillain-Barré syndrome and Miller-Fisher syndrome (5–7). C. jejuni thrives at an optimum temperature of 42°C, which suggests its ability to adapt to the higher body temperature of poultry in comparison to mammals (2). However, C. jejuni is more sensitive to environmental stress than other foodborne pathogens and has fastidious growth requirements (8), such as a narrow temperature range (from 30°C to 45°C) and a microaerobic atmosphere containing carbon dioxide (9). The sensitivity of C. jejuni to environmental stress can be ascribed to the absence of certain stress-adaptive responses that are conserved in other major foodborne pathogens, such as Salmonella and Escherichia coli. For instance, C. jejuni lacks rpoS‐encoded sigma factor stationary phase responsive genes, the major cold‐shock protein CspA, and the alternative sigma factor RpoH that regulates the heat‐shock response in E. coli (10, 11).
Limited information is available regarding the role of heat shock proteins in the thermotolerance of C. jejuni. Previous studies have demonstrated that C. jejuni increases expression levels of heat shock proteins (e.g., ClpB, DnaK, and GroESL) in response to heat stress (12, 13). Recent transcriptomic and proteomic studies have focused on the expression of chaperones and the binding of regulators under heat stress (14, 15). Additionally, changes in the binding affinity of heat shock regulators HrcA and HspR in the promoter region of heat shock proteins after heat stress in C. jejuni have been observed (15). HspR and HrcA negatively regulate the expression of three heat shock proteins: DnaK, ClpB, and GroESL (16–18). While many bacteria survive heat stress by utilizing heat shock proteins, ClpB is particularly well known for its contribution to bacterial survival in cooperation with DnaK by disaggregating protein aggregates formed under stress conditions (19, 20). Despite this knowledge, the specific role of heat shock proteins in C. jejuni remains unclear. Elucidating the heat shock defense mechanisms of C. jejuni in coping with heat stress is crucial for understanding foodborne infections resulting from the consumption of contaminated poultry products. This understanding could potentially lead to improved strategies for controlling C. jejuni contamination during food processing and ultimately enhance food safety.
The molecular mechanisms underlying the adaptive response of C. jejuni to heat stress, particularly during poultry processing steps, remain poorly understood. Specifically, during poultry processing, chickens may be subjected to scalding treatments at temperatures around 50–60°C to facilitate feather removal. This scalding process is critical, as it can significantly impact the microbial load of pathogens, such as C. jejuni, on chicken skin. Despite these treatments, C. jejuni is often resilient and can survive, leading to continued food safety concerns (21). Meanwhile, understanding the genetic diversity among different lineages of C. jejuni, particularly clonal complexes (CCs), is crucial for assessing their ecological and epidemiological implications. Variations in heat tolerance among C. jejuni lineages, especially those identified through multilocus sequence typing (MLST), could significantly influence their survival and success during food processing and under environmental stress. However, to date, there has been no established correlation between specific CCs and thermotolerance in the literature. This absence of information indicates that the relationship between strain lineage and stress resistance remains complex and not yet fully understood. A previous study has highlighted the variability among C. jejuni lineages without correlating these strains with heat tolerance (22). Similarly, findings from Oh et al*.* (23) characterized the stress tolerance of C. jejuni strains isolated from retail raw chicken, reinforcing the notion that comprehensive analyses linking clonal complexes to thermotolerance are lacking. Furthermore, our latest studies explored the phylogenetic associations and genetic factors related to cold stress tolerance but did not establish connections to thermotolerance (24, 25). Therefore, our investigation focuses on elucidating the mechanisms that underlie the divergent heat tolerance observed among C. jejuni lineages, contributing to food safety and public health.
In this study, we reveal a significant correlation between bacterial phylogeny and thermotolerance: MLST CC-443 was predominant among heat-tolerant strains, whereas CC-21 was prevalent in heat-sensitive strains. This phylogenetic association suggests the existence of specific genetic lineages of C. jejuni with enhanced thermotolerance. Furthermore, we demonstrate that the heat shock protein ClpB plays a crucial role in C. jejuni’s heat tolerance by disaggregating heat-denatured proteins under thermal stress conditions.
MATERIALS AND METHODS
Bacterial strains and culture conditions
Eighty-six C. jejuni strains, which were isolated from retail raw chicken in our previous study (26), were used to monitor thermotolerance. C. jejuni NCTC 11168, the first genome-sequenced strain in Campylobacter (27), was used as a wild type in this study. C. jejuni strains were routinely grown on Mueller-Hinton media (Oxoid, Hampshire, UK) at 42°C under microaerobic conditions (5% O_2_, 10% CO_2_, and 85% N_2_) generated by Anoxomat (Mart Microbiology BV, Lichtenvoorde, Netherlands). E. coli was grown at 37°C aerobically in Luria-Bertani media (BD Difco, MD, USA) supplemented with carbenicillin (Cb, 100 µg/mL) or kanamycin (Kan, 50 µg/mL) when required.
Construction of C. jejuni mutants and complemented strains
Construction of C. jejuni NCTC11168 clpB, dnaK, and groESL deletion mutants, as well as their respective complemented strains, has been described previously (28). For the construction of hrcA and hspR mutants and complemented strains*,* we selected the C. jejuni NCTC 11168 as a reference strain as it is commonly used in many laboratories (27). Suicide plasmids carrying hrcA and hspR were constructed as described previously (29). Briefly, each gene and its flanking region were amplified from C. jejuni NCTC 11168 by PCR with GXL polymerase (TaKaRa, Tokyo, Japan) using the primers listed in Table S2. After digestion with SalI and BamHI, the PCR products were each ligated to pUC19 that had been treated with the same enzymes. The pUC19 plasmid containing each gene was amplified by PCR from inside the gene with inverse primers using the same polymerase and ligated with a kanamycin resistance cassette amplified from pMW10 using the Kan-F and Kan-R primers (Table S2). The suicide vectors were commercially sequenced by Bionics (Seoul, Republic of Korea). These plasmids were used as suicide vectors, and each vector was introduced into C. jejuni NCTC 11168 by electroporation. Mutants were screened by growing on MH agar containing kanamycin (50 µg/mL), and the mutations were confirmed by PCR and sequencing.
Complemented strains were constructed by chromosomal integration as previously described (30). Briefly, DNA fragments containing an intact copy of hrcA and hspR were amplified with primer pairs (Table S2) and ligated to the NotI site of a pUC19 derivative carrying an rRNA gene cluster, pFMBcomCM. pFMBcomCM facilitates the chromosomal integration of the complemented genes. The expression of these genes inserted into pFMBcomCM is driven by the native rRNA promoter, allowing for the proper regulation of gene expression in the context of the C. jejuni cellular environment. Plasmids carrying the genes were sequenced by Bionics (Seoul, Republic of Korea) and used as complementation vectors. The complementation vectors were introduced into ΔhrcA and ΔhspR mutants by electroporation. The complemented strains were selected by growing on MH agar plates supplemented with kanamycin (50 µg/mL) or chloramphenicol (12.5 µg/mL). The complementation of each gene was confirmed by PCR and sequencing.
Thermotolerance test
An overnight culture grown on MH agar was resuspended in MH broth to an optical density of 600 nm (OD_600_) of 0.1 (ca, 10^9^ CFU/mL). The bacterial suspension was transferred to 0.2 mL 8-Strip PCR SnapStrip Tubes, Flat Cap (SSIbio, CA, USA) in 100 µL aliquots. The PCR tubes were sealed and incubated at 50°C for 60 min, and samples were taken at 10 or 20 min intervals for serial dilution and bacterial counting. The detection limit of the assay was 200 CFU/mL.
Determination of delta (D)-value
D-values, the time required to kill 90% of the organism during heat treatment, were determined by the linear regression technique (31, 32). Multiple linear regression models with R^2^ values were higher than 0.85 for all cases. Thermal inactivation curves were determined by plotting populations of C. jejuni on a logarithmic scale as a function of heating time at 50°C. D-values were calculated from the slope of a simple regression line for each thermal inactivation curve at 50°C.
Thermotolerance tests under chicken scalding conditions
C. jejuni-spiked chicken skin samples were prepared as previously described (22, 24). Pieces of raw chicken skin (≈1 cm^2^/piece) were prepared from retail raw chicken skin by cutting with a sterile scissor. Raw chicken skin pieces were dipped in 70% ethanol overnight at 4°C to eliminate bacterial contaminants. The remaining contaminants were sterilized by exposing the chicken skin pieces to ultraviolet light on each side for 30 min. The sterilized-dried chicken skin pieces were placed into 24-well plates. C. jejuni suspensions were prepared by diluting an overnight culture to an OD_600_ of 0.1 with Dulbecco’s phosphate-buffered saline without calcium and magnesium (DPBS) (GenDEPOT, TX, USA). An aliquot of 10 µL of culture (ca, 10^7^ CFU) was inoculated to the surface of chicken skin to obtain an initial population of approximately 10^6^ CFU/mL after dipping in 0.1% buffered peptone water (BPW) (KisanBio, Seoul, Republic of Korea). Negative controls were prepared by inoculating 10 µL of DPBS on chicken skin. Then, all inoculated skin samples were left at room temperature for 30 min to allow bacterial cells to adhere to the skin surfaces.
To determine the impact of thermotolerance on poultry processing interventions, we mimicked the conditions of chicken scalding treatment as described previously with slight modifications (21, 33). A 50 mL falcon tube containing 30 mL of 0.1% BPW was heated at 50°C in the water bath for 30 min before the test. Then, chicken skin pieces spiked with C. jejuni were individually dipped into each pre-warmed falcon tube containing 0.1% BPW and incubated at 50°C for 5 min. Every 1 min for 5 min, the chicken skin was taken from the falcon tube with a sterile tweezer, transferred to 15 mL tubes containing 1 mL of 0.1% BPW, and vigorously vortexed for 2 min. Supernatants were collected for serial dilution and bacterial counting. The detection limit of the assay was 100 CFU/mL.
Bacterial adenylate cyclase two-hybrid system assay
The bacterial adenylate cyclase two-hybrid (BACTH) system was used to examine the protein interaction between ClpB and DnaK. Protein interaction assays were performed following the previously described protocol (34). Briefly, for the BACTH constructs, ClpB and DnaK were amplified from C. jejuni NCTC 11168 by PCR with GXL polymerase (TaKaRa, Tokyo, Japan) using the primers in Table S1. After digestion with XbaI and KpnI, the PCR products of ClpB and DnaK were each ligated to pUT18C and pKT25, which had been treated with the same enzymes. Each pUT18C and pKT25 plasmid was co-transformed into chemically competent E. coli BTH101 (cya mutant) and plated in LB-X-Gal plates. BACTH screening LB-X-Gal plates include LB agar supplemented with carbenicillin (Cb, 100 µg/mL) and kanamycin (Kan, 50 µg/mL) with 20 µg/mL 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside (X-Gal) (Duchefa Biochemie, Haarlem, The Netherlands). For positive control, ClpB and DnaK of E. coli MG1655 were used in both pUT18C and pKT25 backgrounds, as they have previously been shown to interact (19, 35) (Table S1). Negative controls were constructed using empty vectors of pUT18C and pKT25. Colonies were individually picked and grown overnight at 37°C in LB supplemented with Cb or Kan. This cell suspension was used as inoculum for all subsequent screening and growth experiments.
The β-galactosidase levels were determined by two methods. To culture cells for β-galactosidase activity measurement, cells grown overnight were sub-cultured in LB media supplemented with Cb or Kan, and 0.5 mM isopropyl β-D-1-thiogalactopyranoside (IPTG, Sigma-Aldrich, MO, USA) was added to induce protein expression. For plate assays, 10 µL of sub-cultured cells was spotted on LB-X-Gal plates, and plates were incubated at 30°C for 48 h under aerobic conditions. For quantitative measurement, β-galactosidase activity was determined using a liquid assay (36). Subcultures were centrifuged, and pellets were resuspended with Z-buffer (60 mM Na_2_HPO_4_·7H_2_O, 60 mM NaH_2_PO_4_·H_2_O, 10 mM KCl, 1 mM MgSO_4_, 50 mM β-mercaptoethanol, adjusted to pH 7.0) to an OD_600_ of 0.7 (blank against Z-buffer). To permeabilize the bacterial cells, sodium dodecyl sulfate and chloroform were added, followed by a light vortex for 10 s, and incubated at room temperature for 10 min. Then, o-nitrophenol-β-galactoside (ONPG; 4 mg/mL) was mixed with gentle inverting. Finally, the reaction was stopped by raising the pH of the solution with 1 M Na_2_CO_3_ precisely after 30 min of incubation time at 37°C. The OD_420_ and OD_550_ were determined and normalized to the cell density in the washed cell suspension (OD_600_), the volume of culture used in milliliters, and the time in minutes of incubation prior to halting the reaction. The β-galactosidase activity was recorded in Miller units.
Quantification of protein aggregates
C. jejuni cultures were taken after 0, 10, and 20 min exposure to 50°C, washed twice with ice-cold DPBS (GenDEPOT, TX, USA), and centrifuged at 10,000 ×g for 5 min. Following the manufacturer’s instructions, samples were stained with PROTEOSTAT aggresome detection reagent (Enzo Life Sciences, Inc., NY, USA). The PROTEOSTAT dye precisely intercalates into the cross-beta spine of aggregated and misfolded proteins, which hinders the dye’s rotation and leads to strong fluorescence. Briefly, pellets were stained in the dark with PROTEOSTAT by incubating at room temperature for 15 min. Samples were placed onto a 96-well plate (black opaque; Corning, NY, USA), and fluorescence was measured with the SpectraMax i3 platform (Molecular Devices, CA, USA) at 500 nm excitation and 600 nm emission wavelengths. The relative fluorescence units (RFU) were normalized to CFU/mL of the sample and reported in RFU/CFU.
Confocal fluorescence microscopy
Overnight cultures of C. jejuni grown on MH agar were suspended in MH broth to an OD_600_ of 0.1. The bacterial suspension was transferred to a disposable culture tube (Kimble, NJ, USA) and incubated at 42°C with shaking (200 rpm) for 6 h. Bacterial cultures were suspended in MH broth to an OD_600_ of 0.1 and transferred to a PCR tube in 100 µL aliquots. The PCR tubes were heat treated at 50°C for 20 min.
For staining and fixing steps, samples incubated at 50°C were taken after 0, 10, and 20 min, washed twice with ice-cold DPBS, and centrifuged at 10,000 ×g for 5 min. Pellets were stained with SYTO 9 Green Fluorescent Nucleic Acid Stain (Invitrogen, MA, USA) and PROTEOSTAT dye, incubating for 20 and 15 min, respectively, at room temperature without light. Then, pellets were washed with ice-cold DPBS and fixed in 4% paraformaldehyde in DPBS at room temperature for 20 min. After fixation, bacterial cells were washed twice with ice-cold DPBS to observe fluorescence.
Slide glass (Paul Marienfeld GmbH & Co. KG, Laud-Königshofen, Germany) with a thickness of 1 mm was coated with 0.01% (w/v) poly-L-lysine solution (Sigma-Aldrich, MO, USA) to improve cell attachment. Then, each sample (5 µL) was dropped onto a poly-L-lysine-coated slide glass. Samples covered with a cover glass were examined on a confocal laser scanning microscope SP8 X (Leica, Wetzlar, Germany) using appropriate filters with fixed excitation/emission wavelengths at 488 nm/500–600 nm for SYTO 9 and excitation/emission wavelengths at 488 nm/600–700 nm for the PROTEOSTAT dye.
Cross-sectional transmission electron microscopy
Bactericidal protein aggregation can be observed by transmission electron microscopy (TEM) of cross-sections of resin-embedded bacteria (37). Samples incubated at 50°C were taken after 0, 10, and 20 min, washed twice with ice-cold DPBS, and centrifuged at 10,000 ×g for 5 min. Pellets were fixed at 4°C overnight with Karnovsky’s fixative consisting of 3% glutaraldehyde and 2% paraformaldehyde in 0.05 M sodium cacodylate buffer (pH 7.2). After primary fixation, each sample was washed thrice with 0.05 M sodium cacodylate buffer at 4°C for 5 min. Each sample was post-fixed for 1 h at room temperature with 0.05 M sodium cacodylate buffer containing 1% osmium tetroxide. The fixed sample was rinsed three times with distilled water at 4°C for 5 min and stained overnight with 0.5% uranyl acetate in distilled water at 4°C. The stained samples were dehydrated at room temperature in an ethanol gradient of 30, 50, 70, and 90% for 20 min and finally repeated three times at 100%, each step with slow rotation. Finally, cells were infiltrated with slow rotation using a 1:1 mix of 100% ethanol and Spurr’s resin for 2 h, following 2:1 of 100% ethanol and Spurr’s resin, and subsequently left in 100% Spurr’s resin overnight. For final infiltration, samples were re-immersed in 100% fresh Spurr’s resin for 2 h under slow rotation and polymerized for 24 h at 70°C. The polymerized specimen block was sectioned with a 5 nm thickness using an Ultramicrotome EM UC7 (Leica, Wetzlar, Germany). The cross-sectional images were observed using a transmission electron microscope JEM1010 (JEOL, Tokyo, Japan) at an accelerating voltage of 80 kV.
Sequence alignment and phylogenetic analysis
The phylogenetic tree was visualized via iTOL v6 based on amino acid sequence alignments of ClpB homologs generated by CLC Main Workbench v7.7.1.
Statistical analysis
Student’s t-test was performed for comparative analysis between the two groups. A χ test was conducted to compare the proportion of the two groups. GraphPad Prism v8.0.1 (GraphPad Software, Inc., CA, USA) was used for statistical analysis.
RESULTS
C. jejuni exhibits diverse thermotolerance profiles with phylogenetic association
C. jejuni grows at temperatures ranging from 37 to 45°C, with an optimum at 42°C, and is considered thermotolerant (2). While most pathogenic Campylobacter species are thermophilic, they are typically sensitive to heat treatments above 50°C, which is a temperature range for pasteurization and cooking. Despite heat treatments during processing, such as the scalding of chicken carcasses, C. jejuni is still frequently found in retail poultry meat (21, 38). To understand the mechanisms underlying the survival of C. jejuni under heat stress, we measured the survival of 86 C. jejuni strains at 50°C, which were isolated from retail raw chicken in our previous study (26). C. jejuni NCTC 11168 was selected as a reference strain, as it is one of the common C. jejuni strains used by many laboratories (27). The 86 strains that were tested exhibited various survival patterns in their viability at 50°C for 60 min (Fig. 1A). For a comparative analysis of their thermotolerance, based on the log reduction values after heat treatment for 60 min, 86 strains were divided into two groups of equal size (n = 43). The group with a higher log reduction value was classified as heat-sensitive, while the other group was classified as heat-tolerant (Fig. 1B). The log reduction value of the dividing point was approximately 4.72 at 60 min. Heat-sensitive strains (n = 43) showed significantly higher log reduction values (P < 0.0001) compared to heat-tolerant strains (n = 43).
*Differential survival of 87 C. jejuni strains under heat stress and variations in the distribution of multilocus sequence typing (MLST) clonal complexes (CCs) between heat-sensitive and heat-tolerant strains. (A) Viable counts of 87 C. jejuni strains were measured at 0, 20, 40, and 60 min of exposure to 50°C in Mueller-Hinton broth. The experiment was repeated three times, and each dot represents the average of three replicates. The dotted line indicates the detection limit (200 CFU/mL). (B) Log reduction values of heat-sensitive and heat-tolerant strains after 60 min of heat treatment. In panels A and B, colored circles represent C. jejuni strains as follows: orange, C. jejuni isolates (n = 86); pink, heat-sensitive strains (n = 43); blue, heat-tolerant strains (n = 43); and black, reference strain (C. jejuni NCTC 11168). The experiment was performed in triplicate with consistent results. Black lines indicate mean values. (C) Comparison of MLST CC proportions between heat-sensitive (n = 43) and heat-tolerant strains (n = 43). Student’s t-test was used to compare viabilities between heat-sensitive and heat-tolerant strains. A χ test was conducted to compare the proportions of CCs. Significance levels are denoted as follows: *, P < 0.05; **, P < 0.01; ***, P < 0.0001. CC, clonal complex; UA, unassigned to any defined CC.
Among the heat-sensitive strains, those belonging to MLST CC-21 accounted for 55.6% of the population, indicating a prevalence of these strains (P < 0.001) (Fig. 1C). Strains belonging to CC-45, CC-61, CC-354, and UA present in the heat-sensitive population were not found to be statistically dominant among the heat-sensitive strains when compared to the heat-tolerant strains (Fig. 1C). In contrast, a significant proportion of heat-tolerant strains belonged to CC-443 and CC-607 (13.3 and 8.9%, respectively) and exhibited a high proportion (P < 0.01and P < 0.05, respectively) (Fig. 1C). These results indicate that C. jejuni exhibits diverse survival patterns under heat stress, and thermotolerance is associated with its phylogeny.
ClpB plays a critical role in C. jejuni survival under scalding conditions
Bacterial cells respond to exposure to temperatures above optimal growth through synthesizing heat shock proteins (11). We sought to elucidate the role of heat shock proteins in the thermotolerance of C. jejuni during poultry processing steps that involve heat treatment. The wild type background used in our experiments was C. jejuni NCTC 11168, which belongs to CC-21. First, we tested knockout mutants defective in heat shock chaperone genes, including clpB, dnaK, and groESL, which are repressed by two heat shock response regulators, HrcA and HspR, under normal conditions (14, 16). We evaluated the survival of heat shock chaperone mutants at 50°C (Fig. 2A). A ΔclpB knockout mutant showed a rapid decrease in its viability at 50°C and failed to recover after 40 min of heat treatment (Fig. 2A). Meanwhile, the viability of other mutants, including a ΔdnaK knockout mutant, remained comparable to that of WT during heat treatment (Fig. 2A). Next, we determined delta (D)-values with survival data from the heat treatment experiment (Fig. 2A) using the log-linear model (Fig. S1). The D-value refers to the time required at a specific temperature to reduce the viable population of an organism by 90%, or one log cycle. This parameter is crucial for understanding the thermal inactivation kinetics of C. jejuni during heat treatment. When D-values were compared, the ΔclpB mutant showed significantly different standard errors of the mean (P < 0.0001) compared to WT (Table 1). The D-value markedly decreased to 4.79 min in the ΔclpB mutant, while WT showed 12.45 min at 50°C, and the clpB-complemented strain restored its D-value up to 13.46 min (Table 1). These results indicate that ClpB plays a critical role in the thermotolerance of C. jejuni.
*Contribution of clpB to thermotolerance in C. jejuni. (A) Survival of C. jejuni strains was measured during 60 min of exposure to 50°C. The dotted line indicates the detection limit (200 CFU/mL). (B) Viability of C. jejuni strains measured after exposure to scalding water at 50°C for 5 min. The dotted line indicates the detection limit (100 CFU/mL). Data shown are representative of three independent experiments with similar results. Error bars represent standard errors of the means. Statistical analysis was performed using Student’s t-test to compare viabilities between WT and indicated strains at each sampling time point. Significance levels are denoted as follows: *, P < 0.05; **, P < 0.01; ***, P < 0.001; ***, P < 0.0001; ns, non-significant; N.D., not detected; WT, C. jejuni NCTC 11168 wild type; ΔclpB, ΔclpB mutant; clpB comp, clpB-complemented strain; ΔdnaK, ΔdnaK mutant; dnaK comp, dnaK-complemented strain; ΔgroESL, ΔgroESL mutant; groESL comp, groESL-complemented strain.
Previous reports have shown that the hrcA·grpE·dnaK operon and the groESL operon are both cooperatively regulated by HspR and HrcA, while clpB is regulated solely by HspR (14, 16) (Fig. S2A). To further investigate the role of these heat shock regulators, we assessed the survival of knockout mutants of the heat shock regulator genes hrcA and hspR. During heat treatment, the ΔhspR mutant exhibited significantly enhanced survival compared to the WT strain (Fig. S2B). The elevated survival of the ΔhspR mutant during heat treatment can suggest a potential derepression of ClpB, but further empirical validation regarding the interaction and regulation of these heat shock proteins seems necessary. Genetic complementation of the ΔhspR mutant with an intact copy of hspR fully restored thermotolerance to WT levels (Fig. S2B). These results corroborate previous studies (14, 16) and suggest that ClpB significantly contributes to the thermotolerance of C. jejuni.
To evaluate the impact of ClpB on C. jejuni survival under conditions mimicking chicken scalding procedures, we assessed the viability of C. jejuni on chicken skin exposed to a scalding temperature of 50°C. The ΔclpB mutant exhibited a significant decrease in viability compared to the WT strain (Fig. 2B). While WT maintained its viability until the end of the experiment (5 min) with less than one log reduction (Fig. 2B), the ΔclpB mutant experienced a rapid decline in viability, showing approximately a 5-log reduction after 5 min (Fig. 2B). Notably, the genetic complementation of the ΔclpB mutant restored its survival under scalding conditions to WT levels (Fig. 2B). These findings strongly suggest that ClpB plays a crucial role in the survival of C. jejuni under heat stress conditions.
C. jejuni ClpB does not bind with DnaK
Previous studies in E. coli have demonstrated that the cooperation of DnaK with ClpB is crucial for protein disaggregation, involving direct binding and interaction between the two proteins (19, 35). Moreover, a rapid decrease in bacterial survival under heat treatment has been observed in an E. coli ΔdnaK mutant (39). In contrast, our results showed no decrease in the survival of the C. jejuni ΔdnaK mutant under heat treatment (Fig. 2A). To further assess protein-protein interactions, we performed BACTH assays using both X-gal indicator plates and liquid assays. These experiments revealed that DnaK and ClpB in C. jejuni do not bind to each other, whereas DnaK and ClpB in E. coli exhibited clear interaction (Fig. 3A and B). These findings suggest that ClpB contributes to the thermotolerance of C. jejuni under heat stress without direct protein binding to DnaK. This distinct mechanism in C. jejuni, differing from the well-characterized DnaK-ClpB interaction in E. coli, demonstrates the unique adaptations of C. jejuni to thermal stress.
*C. jejuni ClpB does not interact with DnaK. (A) Bacterial adenylate cyclase two-hybrid (BACTH) assays demonstrating heterotypic interactions between ClpB and DnaK of C. jejuni. Positive (T18-ClpB of E. coli versus T28-DnaK of E. coli) and negative controls (T18-empty vector versus T28-empty vector) are shown for comparison. (B) Quantification of β-galactosidase activity from the BACTH interactions shown in panel A, expressed in Miller units. The experiment was performed in triplicate. Error bars represent standard errors of the means. Statistical analysis was conducted using Student’s t-test. Significance levels are denoted as follows: **, P < 0.001; ns, non-significant.
ClpB assists C. jejuni in mitigating protein aggregates caused by thermal stress
ClpB is a chaperone that mediates the transformation of aggregated proteins to their native state by disaggregating protein aggregates formed under stress conditions (19). In many bacterial species, ClpB is known to function cooperatively with DnaK as a disaggregase under heat stress (20, 35). However, our findings in C. jejuni reveal that DnaK does not play a significant role in survival during heat treatment, nor does it interact with ClpB ([Fig. 2A and 3](#F2 F3)). To elucidate the role of ClpB in protein disaggregation in C. jejuni, we examined the formation of protein aggregates after heat treatment. We measured protein aggregates using the PROTEOSTAT dye, which binds to the quaternary protein structures found in misfolded and aggregated proteins, causing strong red fluorescence by inhibiting the dye’s rotation (40). Relative fluorescence levels increased in all samples after heat treatment, indicating the formation of protein aggregates due to heat exposure (Fig. 4A). Notably, the ΔclpB mutant showed significantly higher levels of protein aggregates compared to WT after 10 min and 20 min of heat treatment (P < 0.01 and P < 0.05, respectively) (Fig. 4A).
*Protein disaggregation by ClpB of aggregates formed under heat stress in C. jejuni. (A) Comparison of aggregated protein levels during heat treatment at 50°C for 20 min. The fluorescence intensity of the WT strain before heat treatment (0 min) was set as 1. (B) Confocal microscopy images of protein aggregates using SYTO 9 dye (green) and PROTEOSTAT dye (red) in samples heat-treated for 20 min. Merged images of SYTO 9 and PROTEOSTAT dye staining are shown on the right. (C) Cross-sectional transmission electron microscopy (TEM) images of C. jejuni strains before (Initial) and after heat treatment for 20 min at 50°C (heat stress). Protein aggregates are indicated by orange arrows. (D) Aggregates per field of view obtained from cross-sectional TEM images of C. jejuni strains before (0 min) and after heat treatment at 50°C for 20 min. The data shown are representative of three independent experiments with similar results. Error bars represent standard errors of the means. Statistical analysis was performed using Student’s t-test. Significance levels are denoted as follows: *, P < 0.05; *, P < 0.01; ns, non-significant. WT, C. jejuni NCTC 11168 wild type; ΔclpB, ΔclpB mutant; clpB comp, clpB-complemented strain.
To further validate these findings, we employed two widely used techniques for observing protein aggregates: fluorescent dye staining and cross-sectional transmission electron microscopy (TEM) (37, 41). After 20 min exposure to heat treatment, the PROTEOSTAT dye was detected at higher levels in the ΔclpB mutant compared to both the WT and clpB-complemented strains (Fig. 4B), indicating increased protein aggregation in the ΔclpB mutant. Moreover, cross-sectional TEM visualization of the cells revealed more extensive formation of protein aggregates in the ΔclpB mutant compared to the WT and clpB-complemented strains after 20 min of heat treatment (P < 0.01) (Fig. 4C and D). Collectively, these results provide strong evidence that ClpB plays a critical role in the thermotolerance of C. jejuni by mitigating the formation of protein aggregates during thermal stress.
Distinct polymorphisms in ClpB in the CC-443 group
Our observations of differential thermotolerance among various C. jejuni strains (Fig. 1) and the significant role of ClpB in this process ([Fig. 2 and 4](#F2 F4)) led us to hypothesize that thermotolerance may be associated with polymorphisms in the ClpB protein sequence. To investigate this, we conducted a phylogenetic analysis using ClpB amino acid sequences from 86 C. jejuni isolates (Fig. 5). The amino acid sequence of ClpB in strains CW21 (CC-354), CW22 (CC-354), and CS37 (UA) is identical to the ClpB sequence found in all six CC-443 strains (CS13, CS14, CS61, CS62, CS63, and CS64). Consequently, we have designated a total of nine strains sharing this identical sequence as part of the CC-443 group. The ClpB of the CC-443 group exhibited 99.18% identity (850/857 amino acids) with no gaps compared to C. jejuni NCTC 11168, which shares an identical sequence with the CC-21 group (Fig. 5; Fig. S3). Based on the alignment of the ClpB amino acid sequences, we predicted the domain organization of clpB homologs from the CC-443 group and C. jejuni NCTC 11168 (Fig. S3). As the domains of C. jejuni ClpB have not been previously characterized, we used the domains of E. coli ClpB, which consists of the same length (857 amino acids), as a reference for domain prediction. Seven amino acid substitutions were identified in the CC-443 group compared to C. jejuni NCTC 11168: A20V, S44N, T45A, M150I, A474T, A596T, and N726S (Fig. S3).
Analysis of ClpB amino acid sequences from C. jejuni isolates. (A) Phylogenetic tree based on ClpB amino acid sequences from 86 C. jejuni isolates. CC, clonal complex; UA, unassigned to any defined CC.
Notably, ClpB from the CC-443 group exhibited distinct amino acid substitutions in the N-terminal domain (NTD) and the second nucleotide-binding domain (NBD-2). Specifically, we observed changes from alanine (A) to valine (V) in the NTD and to threonine (T) in NBD-2 compared to ClpB in other CC groups (Fig. S3). The NTD of ClpB plays a crucial role in substrate binding and disaggregation activity, although it is dispensable for oligomerization and ATPase activity (42). NBD-1 and NBD-2 exhibit distinct properties, with NBD-2 showing higher affinity for nucleotides and greater ATPase activity (43, 44). These results suggest that although direct data demonstrating their impact on protein folding compared to variants are lacking, these unique substitutions in the NTD and NBD-2 of ClpB may correlate with the differential heat stress tolerance capabilities observed in C. jejuni strains (Fig. 1A).
DISCUSSION
In this study, we sought to elucidate the survival mechanisms of C. jejuni during heat stress by first investigating the phylogenetic relationship between different survival rates among 86 C. jejuni strains isolated from retail raw chicken. Our results demonstrate that C. jejuni exhibits strain-dependent variation in survival ability under heat treatment (Fig. 1A). Comparative analysis of MLST and thermotolerance revealed a significant association between C. jejuni’s phylogenetic background and its thermotolerance (Fig. 1C). Specifically, strains belonging to CC-443 were represented in significantly higher proportion among heat-tolerant strains, whereas CC-21 strains generally displayed sensitivity to heat stress (Fig. 1C). These findings align with our previous studies, which demonstrated that CC-443 strains are also tolerant to aerobic and cold stresses (24–26), suggesting a broader stress resistance phenotype for this clonal complex. Surprisingly, CC-443 strains of C. jejuni show a significant correlation between human and poultry sources. According to the PubMLST database (www.pubMLST.org), among the 81 CC-443 isolates reported thus far, human stool (58.1%) and chicken (30.9%) represent the primary sources. This distribution strongly suggests that CC-443 strains possess a significant potential for foodborne transmission from poultry to humans potentially due to their enhanced stress tolerance. The ability of these strains to withstand various environmental stressors, including heat, may contribute to their survival during food processing and storage, thereby increasing their likelihood of causing human infections. However, further studies are still needed to investigate the potential threat of CC-443 in human infections.
Previous studies on the heat shock proteins of C. jejuni have revealed their transcriptomic and proteomic changes and binding of heat shock regulators to promoter regions (14–16). However, the specific function of the heat shock chaperones in thermotolerance in C. jejuni has not been thoroughly investigated. Therefore, it is fundamental to understand how chaperones function in C. jejuni. In this study, we evaluated the function of heat shock chaperones in the survival of C. jejuni under heat stress. Interestingly, among the knockout mutants of heat shock chaperones, only the ΔclpB mutant showed a significant difference in viability compared to WT under heat treatment (Fig. 2A). The D-values, which represent the time required to reduce the bacterial population by 90% at a specific temperature, also showed a considerable difference in the ΔclpB mutant, indicating that clpB is a crucial genetic element in the thermotolerance of C. jejuni (Table 1). Additionally, the ΔhspR mutant exhibited significantly elevated levels of survival compared to WT (Fig. S2) possibly due to the elevated expression of clpB. The hspR-complemented strain showed slightly decreased survival compared to WT (Fig. S2), suggesting that the overexpression of HspR may lead to increased repression of clpB (18, 45, 46). Still, while our results indicated a possible regulatory effect of HspR on ClpB, a definitive elucidation of this relationship required additional studies involving double mutant analyses. A comprehensive understanding of the regulatory network governing heat shock responses in C. jejuni is critical.
It is widely recognized that E. coli ClpB cooperatively functions to disaggregate protein aggregates by directly binding to DnaK (20); thus, previous studies have focused on identifying the binding sites of ClpB and DnaK (19, 35). However, our results using the BACTH system showed that ClpB and DnaK of C. jejuni do not bind to each other (Fig. 3). This functional independence between ClpB and DnaK in C. jejuni may explain the different survival patterns observed in these mutants under heat treatment; the ΔclpB mutant showed markedly lower survival than WT at 50°C, whereas the viability of the ΔdnaK mutant remained comparable to that of WT (Fig. 2A). Although previous screenings may not have detected a direct interaction (47), the absence of such evidence in C. jejuni supports our hypothesis that ClpB operates without the cooperative binding typically observed in other organisms like E. coli. Furthermore, our data showing the significant impact of ClpB on survival under heat stress, even in the absence of DnaK, reinforce the notion that C. jejuni has developed unique adaptations for thermotolerance. While the stand-alone disaggregase ClpG has been reported to confer thermotolerance in Pseudomonas aeruginosa (39), further studies on ClpB of C. jejuni are necessary to determine whether ClpB functions independently as a standalone disaggregase or cooperates with a chaperone protein other than DnaK. Our results demonstrate that protein aggregates are formed at significantly higher levels in the absence of clpB compared to the WT strain during heat treatment (Fig. 4), potentially resulting in a marked decrease in the survival of the ΔclpB mutant. Collectively, our results show the disaggregating role of ClpB in C. jejuni under heat treatment without direct association with DnaK. It is worth noting that a better understanding is needed about which step ClpB takes its role, whether it acts as a protectant in the prior step of aggregation formation or disaggregates protein aggregates formed after heat stress.
Our study reveals a phylogenetic association with thermotolerance among C. jejuni strains. Certain clonal complexes, specifically CC-443 and CC-607, exhibit higher heat tolerance, while others, such as CC-21, demonstrate increased heat sensitivity (Fig. 1C). This knowledge can lead to targeted approaches in managing C. jejuni contamination based on the prevalence of tolerant CCs. Notably, we observed two distinct amino acid substitutions in the NTD and NBD of ClpB in the thermotolerant CC-443 group (Fig. 5). We also examined whether any of the other thermotolerant strains carried the same allele or shared point mutations with the CC-443 group (Fig. 5; Fig. S3). Our analysis revealed that while they may possess varying levels of heat tolerance, the specific unique substitutions observed in the CC-443 group (A20V and A596T) are indeed distinctive (Fig. S3). These domains play crucial roles in substrate binding and disaggregation activity (48, 49). The NTD fluctuates rapidly and mediates allosteric interactions with the regulatory middle domain and NBDs, thereby influencing ClpB’s overall activity (50). Previous research has shown that mutations in conserved amino acids in the NTD can enhance substrate binding (48), suggesting that the observed substitutions in CC-443 may contribute to its enhanced thermotolerance. The two ATP-binding sites located in the NBD-1 and NBD-2 of ClpB exhibit allosteric interactions that are essential for its chaperone function, with mutations in these sites affecting cooperativity and oligomerization properties (51). Especially, NBD-2 is essential for ClpB function, supporting an unfolding/threading mechanism for disaggregation (49). Moreover, the sequence preservation of NBD-2 seems to be an important factor of the ClpB function, as the affinity of NBD-1 depends on nucleotide binding to NBD-2 (52). Collectively, our results demonstrate that amino acid substitutions in the NTD and NBD-2 of ClpB in the CC-443 group can potentially impact its disaggregation activity through a complex interplay between structural elements and protein function. These modifications may contribute to the enhanced thermotolerance observed in this clonal complex. Altogether, while the unique substitutions observed in the ClpB of the thermotolerant CC-443 strains—specifically the A20V substitution in the N-terminal domain and the A596T change in the second nucleotide-binding domain (NBD-2)—are of interest, further studies are required to elucidate their precise roles in influencing the thermotolerant phenotype. Therefore, our findings suggest a potential correlation between these substitutions and enhanced thermotolerance capabilities, and we acknowledge the necessity for more rigorous experimental validation.
Here, we have demonstrated the significant role of ClpB in aiding the survival of C. jejuni under heat stress. Moreover, our findings indicate that specific phylogenetic groups of C. jejuni are associated with increased thermotolerance. ClpB enhances the survival of C. jejuni by alleviating protein aggregates accumulated due to exposure to heat stress. Our study suggests that the observed differences in thermotolerance among various C. jejuni lineages, particularly between clonal complexes CC-443 and CC-21, have significant consequences for food processing practices. The thermotolerance of CC-443 strains to heat stress indicates that traditional heat treatment methods, such as scalding at 50°C, may not be sufficient to ensure the elimination of all C. jejuni strains in poultry products. Understanding the molecular mechanisms of the thermotolerance of C. jejuni is essential for the effective control of this common foodborne pathogen.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Burnham PM, Hendrixson DR. 2018. Campylobacter jejuni: collective components promoting a successful enteric lifestyle. Nat Rev Microbiol 16:551–565. doi:10.1038/s 41579-018-0037-929892020 · doi ↗ · pubmed ↗
- 2Kaakoush NO, Castaño-Rodríguez N, Mitchell HM, Man SM. 2015. Global epidemiology of Campylobacter infection. Clin Microbiol Rev 28:687–720. doi:10.1128/CMR.00006-1526062576 PMC 4462680 · doi ↗ · pubmed ↗
- 3European Food Safety Authority. 2010. Analysis of the baseline survey on the prevalence of Campylobacter in broiler batches and of Campylobacter and Salmonella on broiler carcasses, in the EU, 2008. EFS 2 8. doi:10.2903/j.efsa.2010.1503 · doi ↗
- 4Golden CE, Rothrock MJ Jr, Mishra A. 2021. Mapping foodborne pathogen contamination throughout the conventional and alternative poultry supply chains. Poult Sci 100:101157. doi:10.1016/j.psj.2021.10115734089937 PMC 8182426 · doi ↗ · pubmed ↗
- 5Blaser MJ. 1997. Epidemiologic and clinical features of Campylobacter jejuni infections. J Infect Dis 176 Suppl 2:S 103–S 105. doi:10.1086/5137809396691 · doi ↗ · pubmed ↗
- 6Blaser MJ, Engberg J. 2008. Clinical aspects of Campylobacter jejuni and Campylobacter coli infections, p 99–121. In Nachamkin In, Szymanski CM, Blaser MJ (ed), Campylobacter, 3rd ed. ASM Press, Washington, DC.
- 7Acheson D, Allos BM. 2001. Campylobacter jejuni infections: update on emerging issues and trends. Clin Infect Dis 32:1201–1206. doi:10.1086/31976011283810 · doi ↗ · pubmed ↗
- 8Gölz G, Kittler S, Malakauskas M, Alter T. 2018. Survival of Campylobacter in the food chain and the environment. Curr Clin Micro Rpt 5:126–134. doi:10.1007/s 40588-018-0092-z · doi ↗