MALDI-TOF lipidomics rapidly detects modification of 2-hydroxymyristate lipid A, a potential virulence trait in Enterobacter bugandensis
Rémy A. Bonnin, Aymeric Jacquemin, Jade Pizzato, Delphine Girlich, Cecile Emeraud, Markus Kostrzewa, Thierry Naas, Gerald Larrouy-Maumus, Laurent Dortet

TL;DR
This study shows how MALDI-TOF lipidomics can quickly detect a virulence trait in Enterobacter bugandensis, which is linked to neonatal septic shock.
Contribution
A rapid MALDI-TOF-based test is introduced for detecting 2-hydroxymyristate lipid A modification in Enterobacter species.
Findings
Only 4 out of 11 E. bugandensis isolates modified their lipid A, correlating with the presence of the lpxO gene.
LpxO-negative isolates had a 1,094 bp deletion encompassing the lpxO gene.
Hydroxymyristate modification was also found in other Enterobacter species, not just E. bugandensis.
Abstract
Enterobacter cloacae complex (ECC) comprises numerous species that can be difficult to identify. Among these species, Enterobacter bugandensis is recognized as a threat in neonatal intensive care units due to its involvement in septic shock. A potential virulence factor responsible for its virulence is the modification of the lipid A of the lipopolysaccharide (LPS). This modification corresponds to a hydroxylation of the myristate component of lipid A. A collection of 168 ECC, including 10 E. bugandensis and one isolate involved in fatal septic shock in the neonatal intensive care unit, were tested for the presence of 2-hydroxymyristate. All isolates were sequenced to determine the genetic diversity of this collection and detect the presence of the lpxO gene responsible for this hydroxylation. matrix assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF)…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
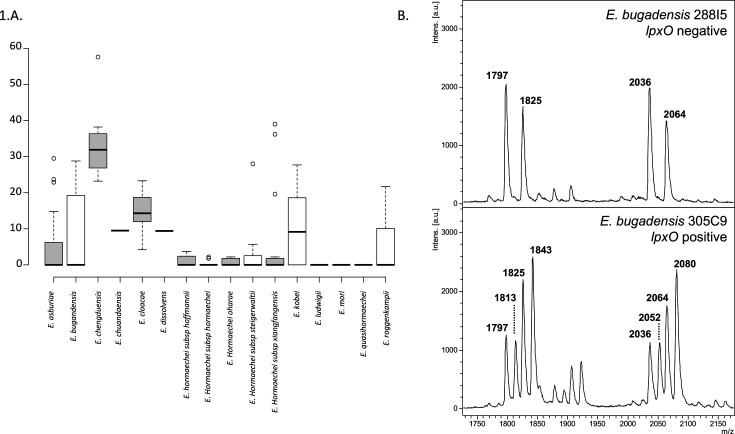 Fig 1
Fig 1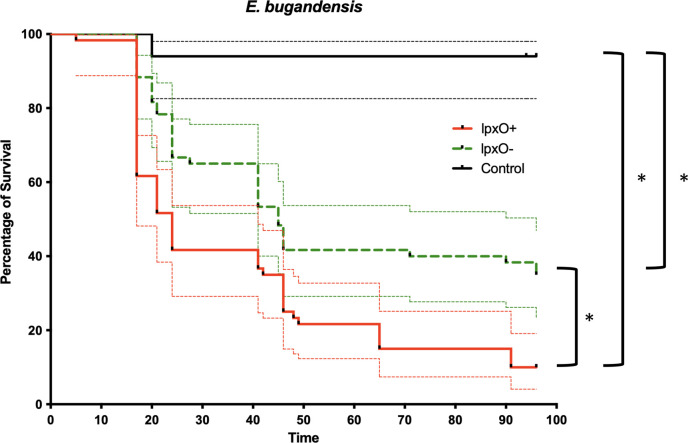 Fig 2
Fig 2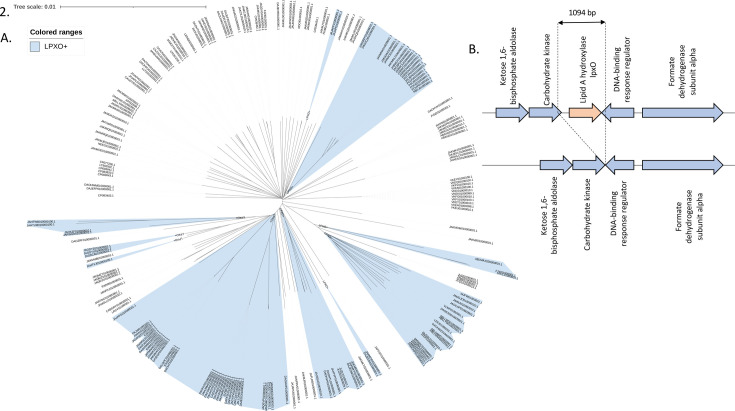 Fig 3
Fig 3| Strain | Species | ST-Oxford | Peak | % of hydroxymyristate | GenBank nucleotide accession number | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1,797 m/z | 1,813 m/z | 1,825 m/z | 1,843 m/z | 2,036 m/z | 2,052 m/z | 2,064 m/z | 2,080 m/z | |||||
| 214 E1 |
| 358 | 4,617 | 0 | 6,191 | 0 | 9,991 | 0 | 11,768 | 0 | 0 |
|
| 268 E7 |
| 250 | 9,396 | 0 | 4,840 | 0 | 8,827 | 0 | 4,174 | 0 | 0 |
|
| 276 E10 |
| 657 | 1,072 | 0 | 485 | 0 | 1,266 | 0 | 460 | 0 | 0 |
|
| 151 C2 |
| 250 | 262 | 0 | 368 | 0 | 860 | 0 | 1,189 | 0 | 0 |
|
| 166D1 |
| 657 | 7,026 | 0 | 2,521 | 0 | 7,934 | 0 | 2,700 | 0 | 0 |
|
| 170 C2 |
| 250 | 4,838 | 0 | 3,523 | 0 | 5,593 | 0 | 3,946 | 0 | 0 |
|
| 183D3 |
| 250 | 704 | 0 | 885 | 0 | 1,850 | 0 | 1,726 | 0 | 0 |
|
| 186 H9 |
| 250 | 909 | 0 | 511 | 0 | 1390 | 0 | 676 | 0 | 0 |
|
| 220D7 |
| 250 | 5,375 | 0 | 6,133 | 0 | 10,092 | 0 | 12,448 | 0 | 0 |
|
| 247 E2 |
| 807 | 1,121 | 0 | 769 | 0 | 1,227 | 0 | 380 | 0 | 0 |
|
| 269 G8 |
| 484 | 4,848 | 0 | 3,751 | 0 | 7,382 | 0 | 2,369 | 0 | 0 |
|
| 172D2 |
| 25 | 7,620 | 489 | 4,037 | 945 | 6,972 | 378 | 2,580 | 0 | 6.2 |
|
| 276I5 |
| 53 | 1,285 | 3,225 | 969 | 2,062 | 8,769 | 3,789 | 1,107 | 1,917 | 22.9 |
|
| 282 G7 |
| 250 | 2,577 | 0 | 2,061 | 0 | 3,173 | 0 | 2,113 | 279 | 0 |
|
| 282I7 |
| 53 | 3,005 | 754 | 2,418 | 533 | 8,883 | 1,477 | 2,756 | 975 | 6.2 |
|
| 287I5 |
| 252 | 2,203 | 3,689 | 2,298 | 3,337 | 6,124 | 2,855 | 1,538 | 1,810 | 29.5 |
|
| 288 F8 |
| 24 | 5,983 | 2,410 | 7,114 | 4,828 | 4,671 | 1,131 | 3,054 | 1,335 | 23.7 |
|
| 288 G10 |
| 252 | 711 | 1,036 | 911 | 998 | 5,487 | 1,633 | 1,328 | 1,618 | 14.8 |
|
| P2B |
| 1084 | 1,528 | 982 | 1,874 | 1,566 | 896 | 620 | 1,050 | 1,037 | 26.7 |
|
| 253 J3 |
| 901 | 3,515 | 0 | 5,129 | 0 | 6,400 | 0 | 7,373 | 0 | 0 |
|
| 118 C5 |
| new | 55,57 | 0 | 6,205 | 0 | 8,230 | 0 | 10,347 | 0 | 0 |
|
| 221 G5 |
| 1677 | 3,247 | 0 | 7,744 | 0 | 2,958 | 0 | 6,609 | 0 | 0 |
|
| 280D5 |
| 1095 | 1,697 | 0 | 1,824 | 0 | 6,702 | 0 | 3,609 | 0 | 0 |
|
| 288I5 |
| 499 | 11,887 | 0 | 9,388 | 0 | 12,907 | 0 | 9,133 | 0 | 0 |
|
| 291 F6 |
| 1677 | 3,035 | 0 | 4,122 | 0 | 7,412 | 0 | 5,690 | 0 | 0 |
|
| 296B2 |
| 2704 | 3,044 | 0 | 6,738 | 0 | 4,661 | 0 | 8,364 | 0 | 0 |
|
| 243 E5 |
| 1843 | 6,464 | 1,748 | 17,604 | 5,172 | 6,738 | 1,389 | 14,929 | 4,801 | 11.8 |
|
| 257D9 |
| 2069 | 3,520 | 2,644 | 4,699 | 5,396 | 4,171 | 1,988 | 3,402 | 3,635 | 27.3 |
|
| 305 C9 |
| 1092 | 6,549 | 6,449 | 12,611 | 18,105 | 7,190 | 6,626 | 11,416 | 16,339 | 28,8 |
|
| 204 E1 |
| 1065 | 0 | 2,185 | 1,524 | 12,260 | 1,035 | 2,648 | 1,997 | 16,152 | 38.2 |
|
| 215D6 |
| 1065 | 170 | 1,081 | 936 | 3,704 | 6,087 | 2,330 | 1,505 | 4,846 | 23.2 |
|
| 223 A3 |
| 1065 | 351 | 1,529 | 908 | 4,124 | 423 | 695 | 337 | 1,450 | 57.6 |
|
| 250D1 |
| 1065 | 3,856 | 4,400 | 5,953 | 9,108 | 3,398 | 2,881 | 4,031 | 5,511 | 34.5 |
|
| 272I4 |
| 598 | 99 | 617 | 618 | 2,426 | 2,800 | 858 | 668 | 2,846 | 27.8 |
|
| 283 F7 |
| 598 | 894 | 899 | 2,745 | 3,922 | 1,542 | 1,258 | 3,240 | 4,086 | 25.9 |
|
| 291 H5 |
| 598 | 554 | 1,310 | 2,300 | 5,106 | 3,706 | 1,226 | 1,716 | 4,226 | 31.9 |
|
| 228 H2 |
| 1189 | 18,404 | 1,679 | 2,085 | 1,189 | 6,718 | 0 | 0 | 0 | 9.5 |
|
| 198 E8 |
| 1540 | 1,531 | 0 | 7,065 | 3,681 | 960 | 0 | 4,453 | 1,943 | 18.7 |
|
| 199 E8 |
| 84 | 3,999 | 783 | 10,122 | 1,885 | 3,570 | 682 | 8,443 | 1,975 | 8.5 |
|
| 166B3 |
| 456 | 12,941 | 2,447 | 12,561 | 4,906 | 6,019 | 159 | 6,320 | 1,480 | 15.7 |
|
| 178 C6 |
| 820 | 2,618 | 782 | 7,189 | 1,598 | 1,512 | 382 | 3,973 | 882 | 12.6 |
|
| 203 G5 |
| 1540 | 4,362 | 1,741 | 11,618 | 7,229 | 2,378 | 790 | 6,857 | 3,490 | 23.3 |
|
| 208 C9 |
| 412 | 2,947 | 1,093 | 9,055 | 3,265 | 2,451 | 779 | 7,358 | 2,910 | 14.6 |
|
| 229 G3 |
| 1516 | 6,022 | 1,098 | 6,859 | 2,114 | 3,904 | 741 | 4,298 | 1,626 | 12 |
|
| 252 G4 |
| 456 | 14,195 | 5,324 | 13,261 | 8,339 | 8,329 | 2,708 | 7,334 | 4,163 | 21.5 |
|
| 257 F5 |
| 1718 | 6,626 | 1,757 | 9,467 | 5,229 | 4,457 | 795 | 5,529 | 2,400 | 19.3 |
|
| 267I4 |
| 1737 | 85,36 | 2,579 | 4,769 | 1,558 | 6,183 | 1,743 | 2,852 | 867 | 14.2 |
|
| 268D8 |
| 1718 | 6,686 | 1,622 | 7,361 | 2,422 | 3,725 | 775 | 4,196 | 1,515 | 14,3 |
|
| 290 C4 |
| 524 | 8,345 | 534 | 7,078 | 660 | 6,255 | 274 | 4,850 | 406 | 4.2 |
|
| 300 A7 |
| 1517 | 7,165 | 1,308 | 11,001 | 2,492 | 6,769 | 1,185 | 9,178 | 2,371 | 9.2 |
|
| 284 A3 |
| 1715 | 6,968 | 1,488 | 4,620 | 853 | 5,442 | 1,267 | 3,686 | 678 | 9.4 |
|
| 281 H8 |
| 764 | 12,269 | 0 | 7,061 | 0 | 8,160 | 0 | 2,368 | 0 | 0 |
|
| 296I6 |
| 102 | 6,119 | 0 | 2,559 | 0 | 3,435 | 0 | 1,055 | 0 | 0 |
|
| 300 J4 |
| 419 | 3,659 | 0 | 2,180 | 0 | 3,220 | 0 | 1,313 | 0 | 0 |
|
| 302 C10 |
| 419 | 4,700 | 0 | 4,398 | 0 | 2,525 | 0 | 1,931 | 0 | 0 |
|
| 307 G4 |
| 168 | 12,400 | 0 | 8,160 | 0 | 12,611 | 0 | 5,988 | 0 | 0 |
|
| 305 J5 |
| 310 | 9,933 | 0 | 7,238 | 0 | 3,539 | 0 | 2,016 | 0 | 0 |
|
| 128 F4 |
| 78 | 161,15 | 0 | 10,139 | 0 | 8,703 | 0 | 3,927 | 0 | 0 |
|
| 146 F7 |
| 729 | 14,313 | 0 | 14,285 | 0 | 7,302 | 0 | 5,963 | 0 | 0 |
|
| 146 F8 |
| 104 | 8,785 | 0 | 7,844 | 0 | 3,901 | 0 | 3,177 | 0 | 0 |
|
| 146 J2 |
| 419 | 10,950 | 0 | 6,413 | 0 | 5,338 | 0 | 2,249 | 0 | 0 |
|
| 155 G4 |
| 419 | 10,626 | 0 | 6,733 | 0 | 4,213 | 0 | 2,040 | 0 | 0 |
|
| 157B10 |
| 118 | 8,484 | 0 | 6,218 | 0 | 2,484 | 0 | 1,323 | 0 | 0 |
|
| 185I6 |
| 933 | 7,653 | 0 | 8,317 | 0 | 4,851 | 0 | 4,727 | 0 | 0 |
|
| 189 C3 |
| 1123 | 8,071 | 0 | 7,508 | 0 | 4,555 | 0 | 3,210 | 0 | 0 |
|
| 280 C5 |
| 1283 | 11,909 | 0 | 6,180 | 0 | 7,119 | 0 | 2,310 | 0 | 0 |
|
| 279 E3 |
| 233 | 13,009 | 1,005 | 10,635 | 0 | 3,822 | 0 | 1,628 | 0 | 3.3 |
|
| 146 A2 |
| 145 | 7,699 | 719 | 9,785 | 0 | 1,252 | 0 | 1,268 | 0 | 3.5 |
|
| 281 H4 |
| 616 | 13,936 | 1,078 | 10,331 | 0 | 14,170 | 0 | 5,005 | 0 | 2.4 |
|
| 282 C8 |
| 233 | 9,327 | 1,123 | 15,609 | 0 | 10,034 | 0 | 4,579 | 0 | 2.8 |
|
| 298 J7 |
| 1740 | 5,900 | 732 | 6,441 | 0 | 4,914 | 0 | 2,062 | 0 | 3.7 |
|
| 301 C7 |
| 683 | 8,586 | 869 | 15,762 | 0 | 2,674 | 0 | 2,827 | 0 | 2.8 |
|
| 302 G1 |
| 168 | 14,575 | 1,146 | 10,236 | 0 | 6,255 | 0 | 3,444 | 0 | 3.2 |
|
| 304I5 |
| 816 | 9,187 | 774 | 13,346 | 0 | 8,385 | 0 | 8,367 | 0 | 1.9 |
|
| 305I10 |
| 683 | 7,953 | 594 | 12,437 | 0 | 2,276 | 0 | 2,757 | 0 | 2.3 |
|
| 307 F9 |
| 97 | 6,416 | 411 | 6,365 | 0 | 4,588 | 0 | 3,272 | 0 | 2 |
|
| 251 J8 |
| 528 | 7,632 | 0 | 9,793 | 0 | 4,339 | 0 | 4,039 | 0 | 0 |
|
| 259I3 |
| 528 | 9,011 | 0 | 9,534 | 0 | 6,999 | 0 | 4,778 | 0 | 0 |
|
| 265 G1 |
| 269 | 15,477 | 0 | 10,450 | 0 | 6,686 | 0 | 3,604 | 0 | 0 |
|
| 267 A4 |
| 6,427 | 0 | 6,304 | 0 | 3,870 | 0 | 2,530 | 0 | 0 |
| |
| 288 E6 |
| 269 | 13,469 | 0 | 12,414 | 0 | 4,687 | 0 | 3,419 | 0 | 0 |
|
| 292B9 |
| 10,328 | 0 | 11,430 | 0 | 13,745 | 0 | 11,754 | 0 | 0 |
| |
| 304I10 |
| 528 | 3,293 | 0 | 8,161 | 0 | 1,757 | 0 | 4,075 | 0 | 0 |
|
| 172I10 |
| 528 | 7,763 | 553 | 12,245 | 0 | 4,593 | 0 | 6,038 | 0 | 1.8 |
|
| 309 A6 |
| 269 | 11,525 | 1,319 | 26,212 | 0 | 6,621 | 0 | 12,891 | 1,566 | 2.2 |
|
| 188D3 |
| 108 | 6,200 | 863 | 32,224 | 0 | 3,135 | 0 | 15,841 | 0 | 1.5 |
|
| 220 H7 |
| 108 | 2,540 | 434 | 9,098 | 0 | 1,962 | 0 | 5,371 | 0 | 2.2 |
|
| 152 E1 |
| 1068 | 6,215 | 706 | 17,811 | 0 | 2,099 | 0 | 6,218 | 0 | 2.1 |
|
| 170I2 |
| 108 | 2,227 | 0 | 8,463 | 0 | 685 | 0 | 2,672 | 0 | 0 |
|
| 269 A7 |
| 108 | 6,011 | 0 | 11,631 | 0 | 2,338 | 0 | 4,500 | 0 | 0 |
|
| 295I10 |
| 1086 | 4,654 | 0 | 20,443 | 0 | 10,786 | 0 | 17,174 | 0 | 0 |
|
| 297 A1 |
| 108 | 1,838 | 0 | 9,459 | 0 | 743 | 0 | 3,848 | 0 | 0 |
|
| 310 C1 |
| 108 | 1,814 | 0 | 7,474 | 0 | 607 | 0 | 2,874 | 0 | 0 |
|
| 306D9 |
| 1116 | 20,442 | 0 | 40,992 | 0 | 9,384 | 0 | 18,378 | 0 | 0 |
|
| 248 E7 |
| 113 | 4,755 | 0 | 2,276 | 0 | 2,279 | 0 | 1,038 | 0 | 0 |
|
| 222 J2 |
| 831 | 4,302 | 0 | 8,144 | 0 | 3,292 | 0 | 4,809 | 0 | 0 |
|
| 223 C7 |
| 51 | 3,120 | 0 | 6,260 | 0 | 903 | 0 | 1,580 | 0 | 0 |
|
| 224 C9 |
| 90 | 6,354 | 0 | 10,724 | 0 | 4,255 | 0 | 5,566 | 0 | 0 |
|
| 237 C9 |
| 45 | 7,913 | 0 | 6,354 | 0 | 3,017 | 0 | 1,738 | 0 | 0 |
|
| 285B3 |
| 111 | 27,827 | 0 | 32,949 | 0 | 18,371 | 0 | 15,431 | 0 | 0 |
|
| 285 F6 |
| 346 | 2,089 | 0 | 1,240 | 0 | 2,388 | 0 | 790 | 0 | 0 |
|
| 285 H5 |
| 177 | 19,409 | 0 | 12,105 | 0 | 9,023 | 0 | 4,066 | 0 | 0 |
|
| 289 F10 |
| 742 | 40,859 | 0 | 32,530 | 0 | 18,721 | 0 | 10,313 | 0 | 0 |
|
| 290B6 |
| 106 | 17,183 | 0 | 659 | 0 | 9,972 | 0 | 0 | 0 | 0 |
|
| 290I4 |
| 175 | 11,090 | 1,212 | 19,746 | 0 | 3,506 | 0 | 3,725 | 0 | 3.1 |
|
| 295 J1 |
| 1296 | 7,930 | 742 | 17,455 | 0 | 3,796 | 0 | 6,713 | 0 | 2 |
|
| 283 J10 |
| 110 | 14,524 | 1,852 | 23,342 | 0 | 4,504 | 0 | 6,501 | 0 | 3.7 |
|
| 283 J8 |
| 50 | 3,400 | 724 | 5,976 | 0 | 1,351 | 0 | 1,339 | 0 | 5.7 |
|
| 296B7 |
| 106 | 5,851 | 4,493 | 7,006 | 6,872 | 5,233 | 3,190 | 4,067 | 3,864 | 28 |
|
| 137I4 |
| 114 | 3,359 | 0 | 9,591 | 0 | 1,185 | 0 | 3,984 | 0 | 0 |
|
| 139I8 |
| 66 | 499 | 0 | 1,693 | 0 | 0 | 0 | 743 | 0 | 0 |
|
| 149I6 |
| 182 | 7,792 | 0 | 19,685 | 0 | 1,802 | 0 | 8,479 | 0 | 0 |
|
| 176B5 |
| 109 | 1,037 | 0 | 7,284 | 0 | 335 | 0 | 1,919 | 0 | 0 |
|
| 184D7 |
| 1261 | 1,101 | 0 | 7,214 | 0 | 2,188 | 0 | 3,616 | 0 | 0 |
|
| 192 F1 |
| 109 | 10,296 | 0 | 31,172 | 0 | 3,141 | 0 | 9,997 | 0 | 0 |
|
| 192 H5 |
| 143 | 1,527 | 0 | 5,768 | 0 | 1,458 | 0 | 3,358 | 0 | 0 |
|
| 201 F6 |
| 171 | 772 | 0 | 5,693 | 0 | 1,540 | 0 | 2,378 | 0 | 0 |
|
| 208 H2 |
| 459 | 1,669 | 0 | 8,410 | 0 | 988 | 0 | 5,095 | 0 | 0 |
|
| 220 A10 |
| 148 | 2,571 | 0 | 20,576 | 0 | 4,506 | 0 | 9,246 | 0 | 0 |
|
| 222B6 |
| 1077 | 8,935 | 0 | 21,219 | 0 | 4,521 | 0 | 9,299 | 0 | 0 |
|
| 157 E2 |
| 98 | 119,502 | 0 | 9,245 | 0 | 33,495 | 0 | 0 | 0 | 0 |
|
| 205I8 |
| 303 | 10,844 | 1,351 | 44,091 | 0 | 3,152 | 0 | 9,693 | 0 | 2 |
|
| 205 J10 |
| 356 | 5,062 | 781 | 24,251 | 0 | 936 | 0 | 4,671 | 0 | 2.2 |
|
| 211 E4 |
| 121 | 8,621 | 1,124 | 35,253 | 0 | 5,563 | 0 | 20,263 | 0 | 1.6 |
|
| 213 E10 |
| 136 | 0 | 0 | 197 | 0 | 0 | 0 | 0 | 440 | 0 |
|
| 175 A9 |
| 171 | 8,540 | 8,922 | 19,553 | 23,818 | 4,900 | 3,943 | 9,529 | 11,128 | 36.2 |
|
| 187 C4 |
| 527 | 2,932 | 0 | 12,391 | 8,412 | 2,664 | 0 | 9,457 | 7,055 | 19.6 |
|
| 199I1 |
| 245 | 9,922 | 10,516 | 36,887 | 49,106 | 4,052 | 4,950 | 15,114 | 22,525 | 39 |
|
| 270 C3 |
| 32 | 9,575 | 3,245 | 11,187 | 3,478 | 8,902 | 2,935 | 10,538 | 4,277 | 12.4 |
|
| 162 E3 |
| 125 | 2,710 | 2,028 | 2,186 | 2,443 | 1,838 | 1,378 | 1,653 | 1,904 | 27.7 |
|
| 243 E3 |
| 56 | 1,668 | 0 | 1,906 | 760 | 3,245 | 1,051 | 2,851 | 1,490 | 5.9 |
|
| 161 E4 |
| 87 | 7,757 | 2,006 | 9,949 | 5,266 | 4,307 | 1,021 | 6,104 | 2,636 | 18.6 |
|
| 260 J7 |
| 910 | 1,918 | 0 | 2,445 | 0 | 3,985 | 0 | 2,849 | 0 | 0 |
|
| 261B10 |
| 1034 | 249 | 0 | 157 | 0 | 504 | 0 | 273 | 0 | 0 |
|
| 238 H9 |
| 13 | 1,767 | 0 | 0 | 0 | 1,170 | 0 | 0 | 0 | 0 |
|
| 265 H2 |
| 374 | 684 | 0 | 0 | 0 | 345 | 0 | 0 | 0 | 0 |
|
| 265 H3 |
| 374 | 18,513 | 0 | 2,334 | 0 | 11,167 | 0 | 1,051 | 0 | 0 |
|
| 293 H7 |
| 20 | 7,915 | 0 | 0 | 0 | 6,713 | 0 | 0 | 0 | 0 |
|
| 62 F4 |
| 130 | 10,950 | 0 | 0 | 0 | 6,250 | 0 | 0 | 0 | 0 |
|
| 120 J4 |
| 1006 | 3,882 | 0 | 0 | 0 | 3,491 | 0 | 0 | 0 | 0 |
|
| 118 G6 |
| 873 | 3,001 | 0 | 5,011 | 0 | 4,414 | 0 | 5,298 | 0 | 0 |
|
| 153 C2 |
| 487 | 1,170 | 0 | 2,954 | 0 | 2,605 | 0 | 3,298 | 0 | 0 |
|
| 207 H2 |
| 873 | 5,608 | 0 | 8,067 | 0 | 6,578 | 0 | 8,210 | 0 | 0 |
|
| 245I7 |
| 873 | 4,413 | 0 | 5,709 | 0 | 6,345 | 0 | 5,493 | 0 | 0 |
|
| 289 A7 |
| 873 | 3,000 | 0 | 3,409 | 0 | 3,627 | 0 | 3,013 | 0 | 0 |
|
| 172D4 |
| 165 | 3,059 | 0 | 2,581 | 0 | 0 | 0 | 0 | 0 | 0 |
|
| 186 J1 |
| 595 | 1,779 | 0 | 1,617 | 0 | 1,588 | 0 | 1,419 | 0 | 0 |
|
| 206 C2 |
| 96 | 408 | 0 | 1,108 | 0 | 648 | 0 | 1,548 | 0 | 0 |
|
| 217 F8 |
| 997 | 1,446 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
|
| 258 A9 |
| 561 | 1,085 | 0 | 612 | 0 | 637 | 0 | 475 | 0 | 0 |
|
| 275 C8 |
| 561 | 1,647 | 0 | 1,250 | 0 | 946 | 0 | 831 | 0 | 0 |
|
| 276 A9 |
| 1614 | 3,102 | 0 | 2,482 | 0 | 1,700 | 0 | 1,579 | 0 | 0 |
|
| 308B3 |
| 165 | 1,267 | 0 | 1,622 | 0 | 1,295 | 0 | 1,804 | 0 | 0 |
|
| 219 E6 |
| 166 | 579 | 0 | 692 | 0 | 799 | 0 | 965 | 0 | 0 |
|
| 159 A4 |
| 1576 | 1,150 | 0 | 2,153 | 1,536 | 674 | 0 | 1,501 | 981 | 19.2 |
|
| 168 A8 |
| 595 | 1,124 | 443 | 0 | 0 | 358 | 117 | 0 | 0 | 21.7 |
|
| 187 G7 |
| 515 | 296 | 0 | 924 | 320 | 937 | 330 | 2,176 | 1,207 | 5.2 |
|
| 192 H1 |
| 523 | 3,807 | 0 | 2,938 | 0 | 6,155 | 401 | 4,428 | 0 | 0 |
|
| 234D7 |
| 1594 | 3,476 | 702 | 3,913 | 731 | 2,293 | 386 | 2,669 | 504 | 9.8 |
|
| 249D5 |
| 1259 | 1,605 | 263 | 2,021 | 332 | 1,087 | 0 | 1,306 | 0 | 9 |
|
| 289 F4 |
| 1614 | 1,549 | 212 | 1,549 | 269 | 935 | 119 | 894 | 0 | 8.7 |
|
| 290B7 |
| 486 | 1,833 | 0 | 0 | 0 | 1,954 | 0 | 570 | 203 | 0 |
|
| 293 J9 |
| 826 | 1,777 | 0 | 2,307 | 0 | 2,796 | 0 | 3,699 | 2,539 | 0 |
|
| 298I3 |
| 595 | 556 | 0 | 886 | 0 | 1,301 | 0 | 2,102 | 1,985 | 0 |
|
| 303 C5 |
| 515 | 99 | 0 | 413 | 374 | 287 | 174 | 628 | 690 | 14 |
|
| 304 J9 |
| 595 | 4,311 | 694 | 7,268 | 2,930 | 2,620 | 0 | 5,052 | 1,859 | 14.7 |
|
| 305 G5 |
| 165 | 2,391 | 266 | 3,136 | 602 | 0 | 0 | 0 | 0 | 13.6 |
|
| 307 F3 |
| 1134 | 1,569 | 0 | 2,764 | 1,090 | 1,444 | 0 | 2,819 | 835 | 10.4 |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEnterobacteriaceae and Cronobacter Research · Antibiotic Resistance in Bacteria · Infections and bacterial resistance
INTRODUCTION
The genus Enterobacter, consisting of gram-negative rods, belongs to the order Enterobacterales and the family Enterobacteriaceae. The List of Prokaryotic Names with Standing in Nomenclature (LPSN) website inventoried 49 validly published species, including synonyms, among which 25 possessed a validated name with taxonomic status (https://lpsn.dsmz.de/, 6 December 2024). Enterobacter spp. are responsible for a variety of nosocomial infections including urinary tract or bloodstream infections (1).
Enterobacter spp. is a member of ESKAPE, encompassing seven species or genra involved in nosocomial infections associated with antimicrobial resistance (2). This genus possesses an inducible cephalosporinase (blaACT-like gene) that can be overexpressed, leading to resistance to third-generation cephalosporins (e.g., ceftazidime and cefotaxime). Resistance to carbapenems can be due to (i) the overexpression of this AmpC associated with a decrease in membrane permeability or (ii) the production of a carbapenemase, an enzyme possessing significant hydrolysis activity toward carbapenems (3).
Nowadays, classification of Enterobacter species remains a complex problem. Although the heterogeneity of E. cloacae complex (ECC) was known and recognized, classical methodologies were not able to robustly discriminate the different species included in this complex. Thus, to overcome this problem, molecular-based methods were advocated (4). First, a classification based on partial sequencing of heat shock protein gene hsp60 was developed to discriminate species among ECC (4). From this study, 12 clusters (I–XII) were determined. The advent of high-throughput sequencing led to a more precise classification based on phylogenetic analysis (5). This classification led to the identification of 22 clades within ECC (6).
Among ECC, Hoffmann’s cluster IX and Chavda’s clade R now corresponds to Enterobacter bugandensis. This species was initially identified in Tanzania during an outbreak of septicemia in a neonatal ward (7). Analysis of the genome of E. bugandensis EB247 revealed that this species might possess several potential virulence factors that could explain its increased virulence (8). Among these potential virulence factors, increased resistance to human serum was observed compared with non-bugandensis Enterobacter isolates (8). Recently, septic shocks involving E. bugandensis with preterm newborns were reported during an outbreak in France (9). In this study, six fatal septic shocks due to ECC were observed during a 14-month period. Whole genome analysis revealed that all these fatal septic shocks were caused by E. bugandensis. Interestingly, septic shocks were caused by different sequence types (STs), including ST1084 (n = 3), ST917 (n = 1), ST1085 (n = 1), and ST1090 (n = 1), strongly suggesting a particular virulence of this species rather than a clone with increased virulence (9). Recently, the role of newborn incubators as the reservoir of Enterobacter spp. was confirmed during another outbreak in neonatology ward (10). This study also concluded that incubators can play a role in the spread of Enterobacter spp., notably the species E. bugandensis and Enterobacter xiangfangensis.
A modification of lipid A, a hydroxylation of a myristate residue of lipid A, was argued to be the main weapon responsible for the increased virulence of E. bugandensis (11). This modification is due to the presence of the lpxO gene encoding an enzyme involved in 2-hydroxylation of lipid A residue. LpxO enzymes have been reported in many gram-negative rods such as, but not limited to, Pseudomonas aeruginosa, Burkholderia pseudomallei, or Klebsiella pneumoniae (12–14).
The aim of this study was to provide a simple diagnostic test to assess the presence of 2-hydroxymyristate on lipid A in ECC (particularly E. bugandensis).
MATERIALS AND METHODS
Strain collection
A collection of 168 Enterobacter spp. recovered from the “French National Reference Center for carbapenem resistance in Enterobacterales” were used in this study except for the isolate E. bugandensis P2B. This previously reported isolate was recovered from a fatal sepsis shock in neonatal (9). Overall, this collection of ECC comprised 16 different species including Enterobacter asburiae (n = 18), E. bugandensis (n = 11), E. chengduensis (n = 7), Enterobacter chuanduensis (n = 1), E. cloacae subspecies cloacae (n = 13), E. cloacae subspecies dissolvens (n = 1), E. hormaechei subspecies hoffmannii (n = 25), E. hormaechei subspecies hormaechei (n = 9), E. hormaechei subspecies oharae (n = 8), E. hormaechei subspecies steigerwaltii (n = 16), E. hormaechei subspecies xiangfangensis (n = 19), Enterobacter kobei (n = 6), Enterobacter ludwigii (n = 4), Enterobacter mori (n = 2), Enterobacter quasihormaechei (n = 5), and Enterobacter roggenkampii (n = 23) (Table 1).
DNA extraction and whole genome sequencing
Genomic DNA of Enterobacter isolates were extracted using the GeneJet genomic DNA extraction kit according manufacturer’s recommendations (Thermo Fischer Scientific, Les Ulis, France). Whole genome sequencing based on Illumina’s technology was performed as previously described (15). After sequencing, raw data were assembled de novo using the CLC genomics v.21 program (Qiagen, Les Ulis, France), and the genomes were analyzed online using software applications available at the Center for Genomic Epidemiology-CGE (https://www.genomicepidemiology.org/).
Bioinformatic analysis of E. bugandensis lpxO genes
In addition to our collection of E. bugandensis, all available sequences of E. bugandensis available in Genbank on the 1st March 2024 were included. A sequence search was performed with the alignment-search tool BLAST+ with all E. bugandensis whole genome sequences as subjects and a 909 bp long sequence of lpxO originated from E. bugandensis. The lpxO gene was searched by BLAST analysis using 80% nucleotide identity over 90% coverage of the gene as criteria. The phylogenetic analysis of the data set consisted of a phylogenetic tree inferred by the maximum likelihood algorithm IQ-tree (best-fit model automatically selected by ModelFinder). It was based on an alignment of the core genome of the data set. The alignment was done using MAFFT, and the core genome was constructed using Panaroo with default parameters.
MALDIxin-based detection of 2-hydroxymiristate modification of lipid A
A volume of 150 µL bacterial suspension was pipetted into a 1.5 mL Eppendorf tube, centrifuged at 15,000 × g for 5 min, and then washed twice with 400 µL double distilled water (ddH_2_O). Lipid A from Enteobacter strains was extracted using the MBT Lipid Xtract Kit (Bruker Daltonic, Bremem, Germany), following the manufacturer’s instructions. Briefly, 100 µL of bacterial suspension was suspended in 50 µL of the MBT Lipid Xtract hydrolysis buffer. Then, 44 µL of the cell suspension was discarded, and the remaining 6 µL was submitted with the lid of the tube closed to a heating process at 90°C for 30 min. The tubes were left for 2 min with the lid open to completely evaporate the buffer. The dried pellets were washed with 50 µL of the MBT Lipid Xtract washing buffer for a few seconds without dissolving the pellet. The total volume of the washing buffer was discarded by pipetting. Finally, 5 µL of the matrix was pipetted up and down for 15–20 s to resuspend the dried pellet, and 2 µL of the suspension was spotted onto either an MSP 96 polished steel target (Bruker Daltonics, Part-No. 8280800) or a MBT Biotarget 96 (Bruker Daltonics, Part-No. 1840375).
To calibrate the MALDI, 0.5 µL peptide calibration standard II (Bruker Daltonik) together with 0.5 µL of its corresponding calibration matrix were loaded onto the target plate along with the samples to be detected. A mixture of 0.4 µL water and 1.2 µL matrix was used as a negative control. Spectra were acquired on MALDI Biotyper Syrius automated using the linear negative-ion mode (laser intensity: 65%, laser frequency: 200.0 Hz, voltage: 15 kV, pulsed ion extraction: 200 ns, detection range: 820–4200 m/z). FlexAnalysis software (Bruker Daltonik) was used for baseline subtraction and peak detection of the spectra.
Galleria mellonella larva infection model
Wax moth larvae (INRA UMR-1319, Université Paris Saclay, Jouy en Josas, France) were used on the day of reception. Larvae with a cream-colored cuticle presenting minimal speckling or discoloration, weighing 200–300 mg and measuring 2–2.5 cm in length were used for each experiment. Four isolates of E. bugandensis were used for worm virulence assays. Two lpxO-positive isolates (P2B and 305C9) and two lpxO-negative isolates (253J3 and 291F6). Overnight bacterial cultures in LB were diluted 1:10 and grown to an OD_600_ of 0.4–0.6. Cultures were centrifuged and bacterial cells were resuspended in physiological sterile water and serially diluted from 10^9^ CFU/mL to 10 CFU/mL. An inoculum of 10^5^ CFU was injected (10 µL of 10^7^ CFU/mL) into the hindmost proleg of healthy larvae (3 × 10 animals per group) using a microinjector (Hamilton syringe model 802 RNW 25 µL, Fisher Scientific, Illkirch, France). Physiological water (10 µL) was injected in the control group. Then, injected larvae were incubated at 37°C for 96 h. Larvae were considered dead when they displayed no movement in response to touch and had turned black. The survival Kaplan-Meier curves were drawn using the GraphPad Prism software (version 6.0 c). Statistical analysis (log-rank test) was performed on three independent experiments (3 × 10 animals per condition) using GraphPad Prism v10 software.
RESULTS
ECC collection analysis
A collection of 168 clinical isolates of representative members of the Enterobacter genus were used in this study. Sequencing followed by genetic analysis revealed a wide genetic diversity. Overall, a total of 118 different STs were identified over the 168 isolates. Of note, 10 different STs were identified among the 11 E. bugandensis isolates. A wide genetic diversity was of great importance for the tested collection to avoid a bias related to a clonal relationship. In our collection, most of the ECC produced a carbapenemase, including VIM, OXA-48-like, NDM, and IMI, reflecting the French epidemiology identified in Enteroabcter spp (16, 17).
MALDIxin-based detection of 2-hydroxymiristate modification of lipid A
The analysis of lipid A of the 168 EC isolates of our collection was performed using MALDIxin protocol on a clinical microbiology routine MALDI-TOF mass spectrometer, MALDI Biotyper Syrius. Peaks analyses are presented in Table 1.
As shown in Fig. 1B, the mass spectrum of E. bugandensis 288I5 (lpxO-negative isolate) is dominated by a set of two peaks assigned to bis-phosphorylated hexa-acyl lipid A and bis-phosphorylated hepta-acyl lipid A. The major peak at m/z 1797 corresponds to hexa-acyl diphosphoryl and lipid A, respectively, containing four C14:0 3-OH, one C14:0, and one C12:0 (18). The peak at m/z 2035 corresponds to hepta-acyl diphosphoryl lipid A four C14:0 3-OH, one C14:0, one C12:0, and one C16:0. Peaks at m/z 1825 and m/z 2064 are also observed in E. bugandensis, which can tentatively be assigned to hexa-acyl diphosphoryl lipid A, containing four C14:0 3-OH and one C14:0, and hepta-acyl diphosphoryl lipid A, containing five C14:0 3-OH, one C14:0, and one C16:0, respectively. In E. bugandensis 305C9 (lpxO-positive strain), additional peaks were identified at m/z 1813, m/z 1843, m/z 2052, and m/z 2080 corresponding to the addition of an hydroxyl group on myristic acid on the native fatty acid chain (11) and annotated as to the 2-hydroxymyristate modified lipid A.
(A) Percentage of hydroxymyristate detected in the different Enterobacter species. Median and quartile are represented. (B) Peak analysis of two representative E. bugandensis isolates possessing or not lpxO gene. m/z values of the peak are indicated.
The ratio of 2-hydroxymyristate modified lipid A was calculated by dividing the intensities of peaks 2-hydroxymyristate modified lipid A (m/z 1813, m/z 1843, m/z 2052, and m/z 2080) by the sum of intensities of all peaks (unmodified + 2-hydroxymyristate modified peaks) (Table 1).
Ratio analysis of lipid A 2-hydroxymyristate modification revealed that (i) this lipid A modification is not unique to E. bugandensis and (ii) this property is not shared by all members of a species (Fig. 1A; Fig. S1). For instance, in three species E. ludwigii, E. mori, and E. quasihormaechei, no 2-hydroxymyristate branched on lipid A could be identified, whereas all representatives of E. chengduensis possessed this modification on their lipid A (Fig. 1A).
In most cases, only some isolates of the species were able to branch this 2-hydroxy-myristate. As an example, in E. bugandensis, only four of the 11 tested strains produced a 2-hydroxymyristate modified lipid A, suggesting that the locus involved in this modification did not belong to the core genome of E. bugandensis.
Of note, a variable level of 2-hydroxymyristate modified lipid A can also be observed alongside the Enterobacter species. E. chengduensis isolates presented a high rate of 2-hydroxymyristate modified lipid A compared with other species. In this species, all isolates (n = 7) presented a modified version of lipid A. However, it should be noted that the seven tested E. chengduensis isolates corresponded to only two different STs (ST1065 and ST598).
Role of 2-hydroxymyristate-branched lipid A in the virulence of E. bugandensis
It has been advocated that 2-hydroxymyristate-branched lipid A is a major virulence factor in E. bugandensis (11). To confirm this hypothesis, Galleria mellonella survival assay was performed with E. bugandensis exhibiting lipid A with or without 2-hydroxymyristate according to MALDIxin results (Fig. 2). Four isolates were selected and tested in triplicate. Two isolates displaying 2-hydroxymyristate modified lipid A, E. bugandensis P2B isolated from a blood culture sample of a preterm neonate who died from a fatal septic shock (ST1084) (9) and 305C9 (ST1092), and two isolates with native lipid A, strains 253J3 (ST901) and 291F6 (ST1677). Over a time span of 4 days post-inoculation, a significant decrease in survival was observed for the two isolates displaying 2-hydroxymyristate-modified lipid A. A median of survival at 24% was observed for lpxO-positive E. bugandensis versus a median of 45% for lpxO-negative isolates. Log-rank Mantel-Cox test indicated that these curves were different with a P value < 0.001 (Fig. 2). It confirmed the involvement of 2-hydroxymyristate lipid A modification in the virulence of E. bugandensis.
Survival rates of G. mellonella infected by either lpxO-positive or -negative isolates or by physiological water. LpxO-positive isolates used were isolates P2B & 305C9 and lpxO-negative isolates were 291F6 & 253J3. Survival rates were performed over a 4-day period and with 3 × 10 animals per group. A star indicated a P value < 0.001.
Distribution of lpxO in E. bugandensis
The addition of 2-hydroxymyristate is the consequence of several successive lipid A modification involving LpxM, LpxL, and LpxO enzymes. However, the final 2-OH hydroxylation is mediated by the lipid A hydroxylase LpxO. This enzyme is encoded by the lpxO gene. Among the 11 E. bugandensis tested isolates, only four exhibited a 2-hydroxymyristate-modified lipid A according to MALDIxin results (Table 1). It suggested that either lpxO did not belong to the core genome of E. bugandensis or this gene is not expressed in all genetic backgrounds. To answer this question, a collection of 209 E. bugandensis genomes from our collection and from the Genbank database were tested for the presence of lpxO (Fig. 3A). Among these 209 genomes only 97 were positive for the presence of lpxO. Unexpectedly, the presence of lpxO was not distributed by phylogenetic branches with a common ancestor but rather distributed across several branches as usually observed for acquired mobile genetic elements. Accordingly, the close genetic environment of lpxO was analyzed to search for potential mobile genetic elements potentially involved in its acquisition. Intriguingly, this analysis did not reveal the presence of any mobile elements (Fig. 3B). Instead, in lpxO-negative isolates, a deletion of 1,094 bp systematically occurred. This deletion encompassed the whole lpxO gene and the 3’ extremities of the two surrounding genes without a trace of any genetic rearrangement linked to mobile element excision.
(A) Distribution of the lpxO gene in E. bugandensis. All available genomes in Genbank on the 1st of March 2024 were included. The lpxO gene was searched by blast analysis with 80% nucleotide identity over 90% coverage of the gene. GenBank accession numbers are indicated on the tree. Isolates possessing the lpxO gene are indicated in blue. (B) Genetic context analysis of the lpxO gene. The lpxO gene is indicated in orange and other genes in blue.
DISCUSSION
E. bugandensis is now recognized as a potential threat in neonatal wards due to its ability to survive in incubators and its virulence properties (9, 10). This increased virulence was proposed to be linked to a modification of lipid A. This modification corresponded to the addition of 2-hydroxymyristate (11). This study aimed to propose a rapid and simple assay to decipher the presence of 2-hydroxymyristate on lipid A directly from the bacterial colony by using MALDI-TOF technology with MBT lipid Xtract assay (Bruker Daltonik, Germany). We tested the robustness of the test on a collection of 168 Enterobacter spp. clinical isolates belonging to 16 different species. This test demonstrated that (i) different species can modify their lipid A by the addition of 2-hydroxymyristate and (ii) this modification can be present in only some isolates within the same species. Indeed, E. bugandensis is not the sole species able to modify its lipid A by adding a 2-hydroxymyristate, and among the E. bugandensis tested isolates, only four over seven gave a positive result. This observation prompted us to look for the presence of the incriminated gene involved in this modification, lpxO. In our collection of seven E. bugandensis isolates, the four strains modifying their lipid A possessed a chromosome-encoded lpxO gene. Overall, this gene was present only in half of all E. bugandensis genomes present in the NCBI database (Fig. 2). Unfortunately, we could not determine the reason for the lipid A can be the absence/presence of lpxO in these genomes. Indeed, in lpxO-negative strains, we observed a systematic deletion of 1,094 bp in size in the bacterial chromosome. Since no genetic mobile elements were identified bracketing this gene or a larger portion, the gene loss in these genomes could be hypothesized but remained enigmatic.
We previously demonstrated that E. bugandensis exhibited a higher virulence compared with other Enterobacter species in a G. mellonella model (9), and this result was confirmed using a mouse model in which LPS from E. bugandensis demonstrated higher immunogenic properties (11). Here, we tested four clinical isolates of E. bugandensis (two with and two without the lpxO gene) in a G. mellonella model. We observed a clear difference between these two conditions confirming the role of lpxO in the virulence of E. bugandensis. However, an interesting point should be highlighted; despite this lipid A modification being involved in virulence, its role might not be sufficient to explain the global virulence of E. bugandensis in neonates. Indeed, some other ECC species such as E. chengduensis or E. xiangfangensis also include isolates displaying a similar rate of 2-hydroxymyristate modification (Table 1; Fig. 1A) but were never reported in neonatal fatal sepsis shock. Accordingly, deeper investigations are still remaining to fully understand the particular virulence of E. bugandensis in neonates. Nevertheless, the presence of lpxO leading to the final 2-hydroxymyristate modification of lipid A can be considered an additional risk factor if E. bugandensis is involved.
Since the MALDIxin assay can be performed easily on the routine machine using a ready-to-use commercial kit (MBT Lipid Xtract Kit, Bruker Daltonic), this test could be performed directly on E. cloacae complex colonies cultured from a screening sample (such as rectal swabs) or from clinical samples recovered from a neonate or from during an environmental sampling of incubators in the neonatal ward.
In the reported fatal cases of septic shocks in newborns, E. bugandensis isolates were susceptible to β-lactams, and thus, treatment was microbiologically efficient but infection led to early septic shock responsible for the patient death (9). This tragic issue resulted in modifications to patient monitoring. Indeed, in the neonate ward, newborns are now screened for ECC carriage. Accordingly, the MALDIxin test might help clinicians detect the presence of lpxO-positive ECC (particularly if E. bugandensis is suspected) and implement measures to avoid potentially fatal septicemia. We assume that this test possesses some limits since it is not specific to E. bugandensis. However, an ECC positive for the presence of 2-hydroxymyristate on lipid A can be considered a risk factor for the newborn. The procedure following the identification of 2-hydromyristate-lipid A ECC remains to be determined. They can include decolonization procedures based on the result of the antibiogram, increased surveillance of the newborn, or implementation of hygiene measures regarding invasive procedures or catheter manipulations.
This MALDIxin assay can also be useful to test ECC strains cultured from environmental sampling of incubators. Indeed, incubators were identified as environmental sources of ECC-related outbreaks in neonatal intensive care units (9, 10). The identification of ECC isolates with 2-hydroxymyristate-modified lipid A could indicate the presence of a lpxO-positive E. bugandensis that can be considered a risk factor for newborns. Accordingly, supplemental hygiene measures such as incubator disinfection or incubator replacement might be considered to limit the risk of further infection of preterm neonates.
To conclude, we provided a simple and rapid test to decipher the presence of 2-hydroxymyristate on lipid A of ECC isolates. This method can decipher the presence of modified lipid A rather than just the presence of lpxO gene that might not be expressed. Although this modification of lipid A is not only present in E. bugandensis, lpxO-positive E. bugandensis is now recognized as a potential threat of sepsis in the neonatal ward. Thus, the MALDIxin assay could be helpful to identify the presence of this pathogen and implement control measures. In addition, this assay used the MALDI-TOF machine already available in many clinical microbiology laboratories for the routine identification of bacterial colonies. Of note, despite being commercially available, the MBT Lipid Xtract Kit required a routine MALDI-TOF machine able to work in a negative mode, which is not the case for the machines of all manufacturers. Indeed, bacterial identification only required the usage of the positive mode (detection of positively charged proteins), whereas the MBT Lipid Xtract Kit allows the identification of negatively charged lipids including lipid A.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Davin-Regli A, Lavigne J-P, Pagès J-M. 2019. Enterobacter spp.: update on taxonomy, clinical aspects, and emerging antimicrobial resistance. Clin Microbiol Rev 32:e 00002-19. doi:10.1128/CMR.00002-1931315895 PMC 6750132 · doi ↗ · pubmed ↗
- 2De Oliveira DMP, Forde BM, Kidd TJ, Harris PNA, Schembri MA, Beatson SA, Paterson DL, Walker MJ. 2020. Antimicrobial resistance in ESKAPE pathogens. Clin Microbiol Rev 33:e 00181-19. doi:10.1128/CMR.00181-1932404435 PMC 7227449 · doi ↗ · pubmed ↗
- 3Bonnin RA, Jousset AB, Emeraud C, Oueslati S, Dortet L, Naas T. 2020. Genetic diversity, biochemical properties, and detection methods of minor carbapenemases in Enterobacterales. Front Med:616490. doi:10.3389/fmed.2020.616490 PMC 785559233553210 · doi ↗ · pubmed ↗
- 4Hoffmann H, Roggenkamp A. 2003. Population genetics of the nomenspecies Enterobacter cloacae. Appl Environ Microbiol 69:5306–5318. doi:10.1128/AEM.69.9.5306-5318.200312957918 PMC 194928 · doi ↗ · pubmed ↗
- 5Chavda KD, Chen L, Fouts DE, Sutton G, Brinkac L, Jenkins SG, Bonomo RA, Adams MD, Kreiswirth BN. 2016. Comprehensive genome analysis of carbapenemase-producing Enterobacter spp.: new insights into phylogeny, population structure, and resistance mechanisms. M Bio 7:e 02093-16. doi:10.1128/m Bio.02093-1627965456 PMC 5156309 · doi ↗ · pubmed ↗
- 6Sutton GG, Brinkac LM, Clarke TH, Fouts DE. 2018. Enterobacter hormaechei subsp. hoffmannii subsp. nov., Enterobacter hormaechei subsp. xiangfangensis comb. nov., Enterobacter roggenkampii sp. nov., and Enterobacter muelleri is a later heterotypic synonym of Enterobacter asburiae based on computational analysis of sequenced Enterobacter genomes. F 1000 Res 7:521. doi:10.12688/f 1000 research.14566.230430006 PMC 6097438 · doi ↗ · pubmed ↗
- 7Doijad S, Imirzalioglu C, Yao Y, Pati NB, Falgenhauer L, Hain T, Foesel BU, Abt B, Overmann J, Mirambo MM, Mshana SE, Chakraborty T. 2016. Enterobacter bugandensis sp. nov., isolated from neonatal blood. Int J Syst Evol Microbiol 66:968–974. doi:10.1099/ijsem.0.00082126637818 · doi ↗ · pubmed ↗
- 8Pati NB, Doijad SP, Schultze T, Mannala GK, Yao Y, Jaiswal S, Ryan D, Suar M, Gwozdzinski K, Bunk B, Mraheil MA, Marahiel MA, Hegemann JD, Spröer C, Goesmann A, Falgenhauer L, Hain T, Imirzalioglu C, Mshana SE, Overmann J, Chakraborty T. 2018. Enterobacter bugandensis: a novel enterobacterial species associated with severe clinical infection. Sci Rep 8:5392. doi:10.1038/s 41598-018-23069-z 29599516 PMC 5876403 · doi ↗ · pubmed ↗