Subclinical and long-term effects of severe acute respiratory syndrome coronavirus 2 infection in Danish farmed mink: implications for disease surveillance
Michelle Lauge Quaade, Mia Mylin Jensen, Thomas Bruun Rasmussen, Tim Kåre Jensen, Anne Sofie Hammer

TL;DR
This study shows that mink infected with SARS-CoV-2 can have serious lung damage even without obvious symptoms, highlighting gaps in disease surveillance.
Contribution
The study reveals subclinical and long-term pulmonary effects of SARS-CoV-2 in mink, suggesting limitations in current surveillance methods.
Findings
SARS-CoV-2-positive mink showed lung lesions even without clinical signs.
Lesions included respiratory epithelial damage, alveolar damage, and vascular issues like thrombi.
Subclinical infections may lead to undetected outbreaks and hidden health risks in mink.
Abstract
The COVID-19 pandemic has caused over 776 million confirmed cases and more than 7 million deaths worldwide. In addition to humans, various animal species have exhibited natural infections, with mink being the only farmed animals consistently linked to severe illness and zoonotic transmission to humans. This study investigates histological pulmonary lesions in Danish farm mink infected with severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), focusing on groups with different clinical signs and outcomes. Histopathological evaluations revealed lesions in SARS-CoV-2-positive mink with and without clinical signs of disease. The main findings in lungs from SARS-CoV-2-positive mink in all study groups were extensive respiratory epithelial damage, acute diffuse alveolar damage, and vascular lesions, including the formation of thrombi. Additionally, immunohistochemical staining…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
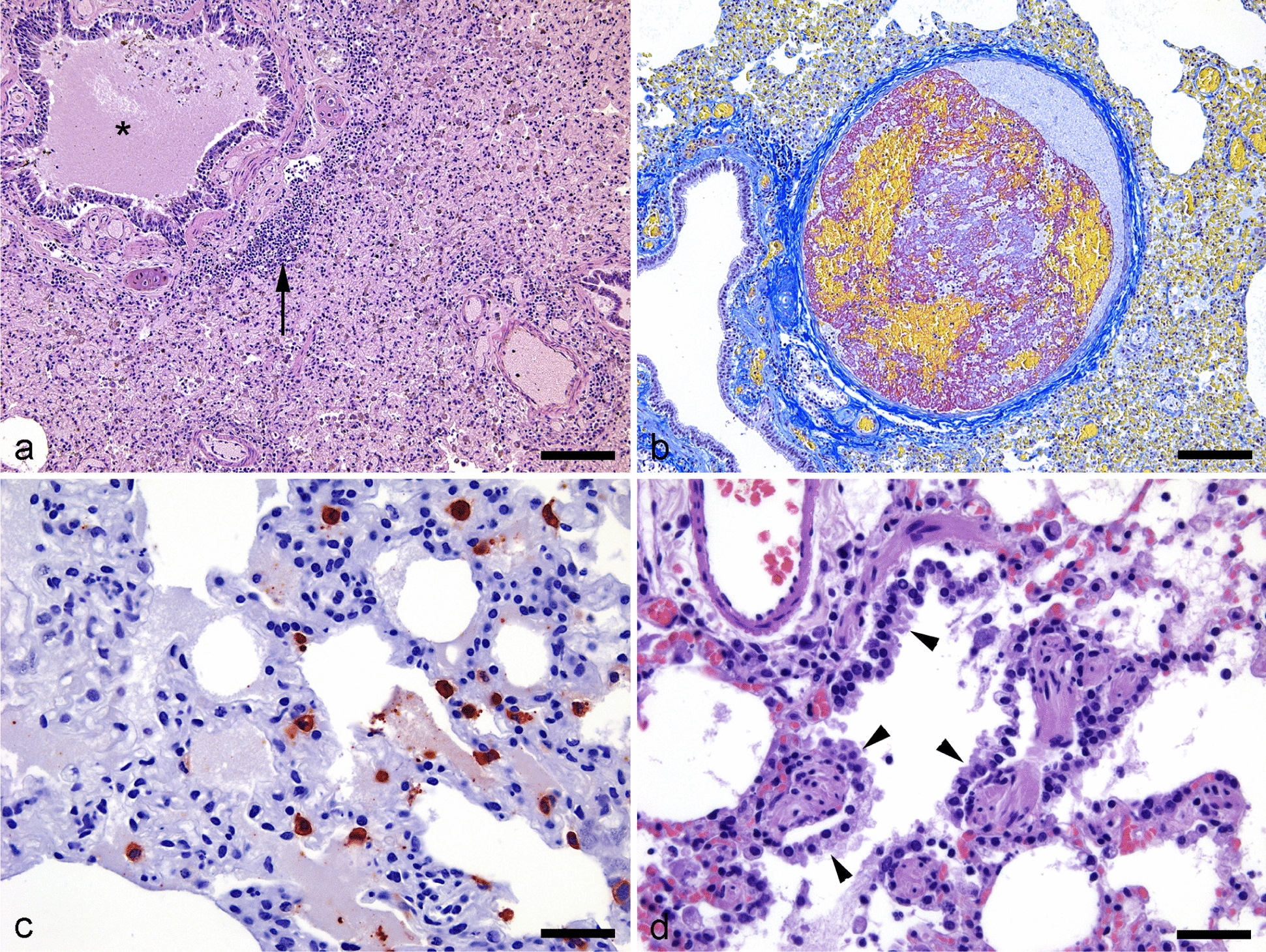 Figure 1
Figure 1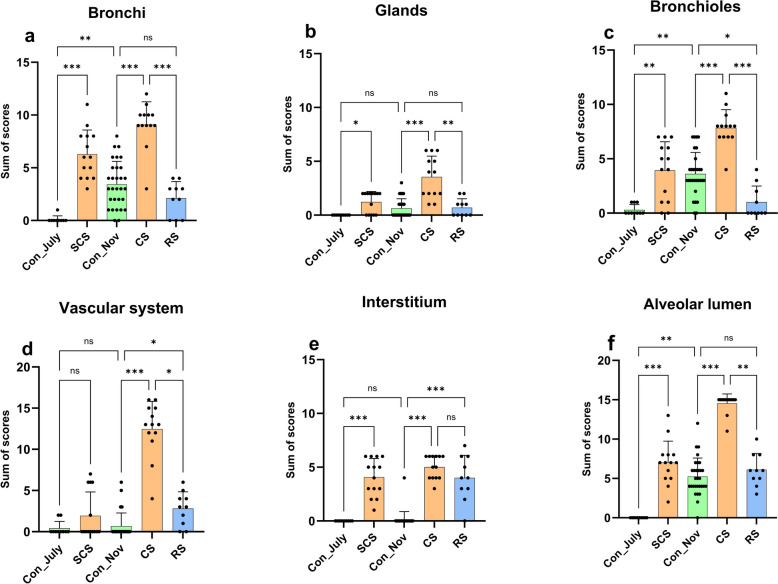 Figure 2
Figure 2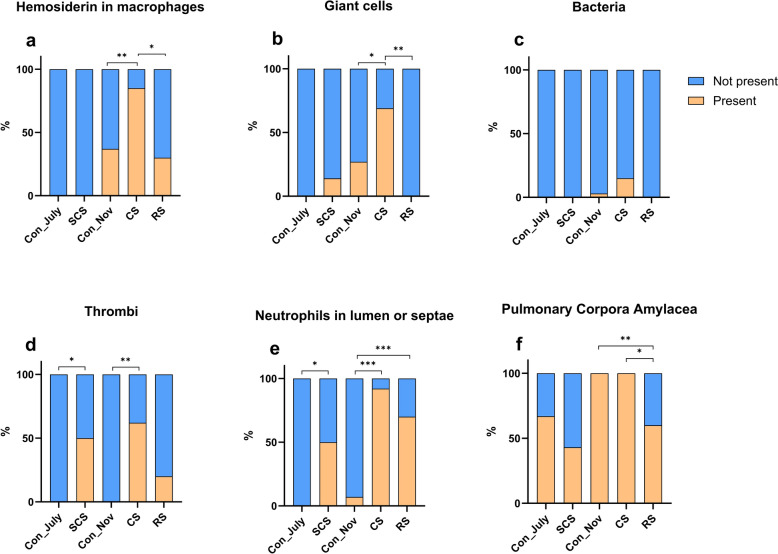 Figure 3
Figure 3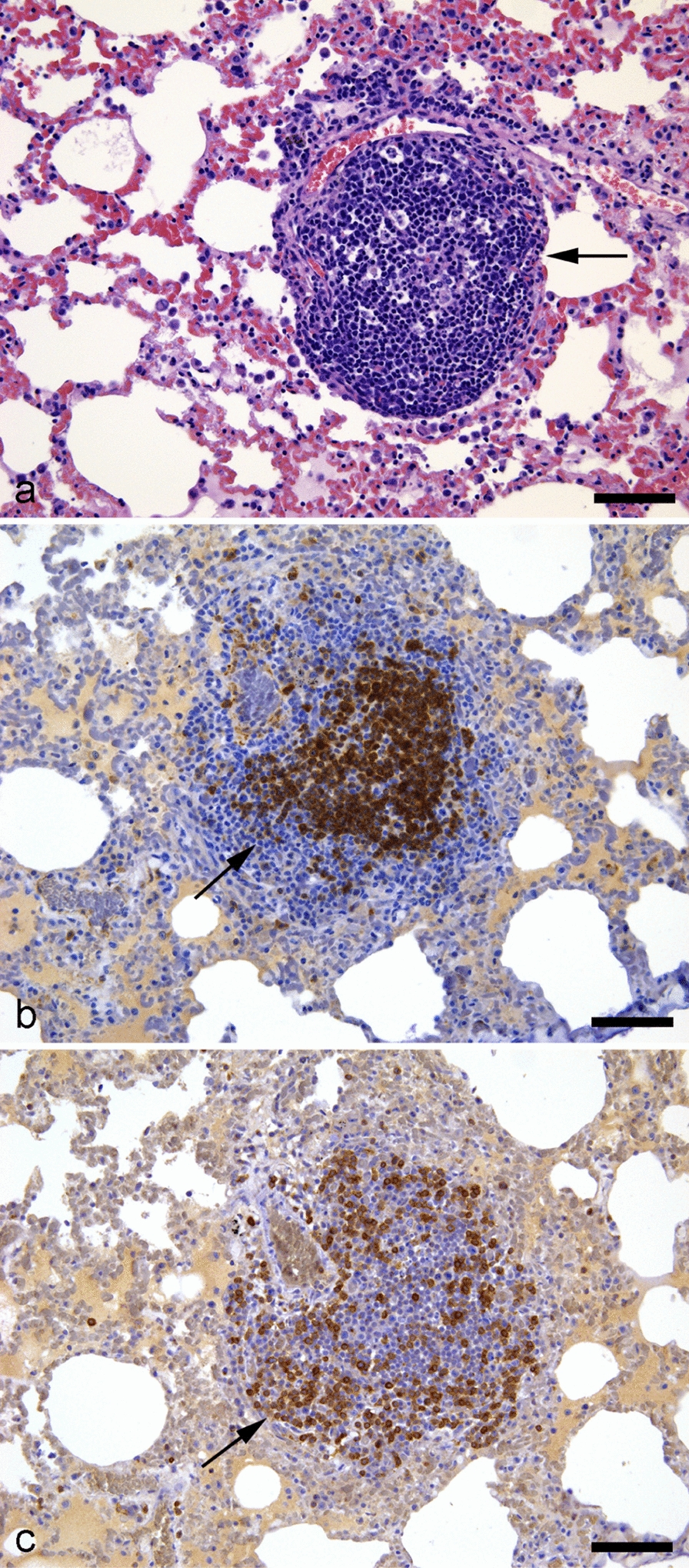 Figure 4
Figure 4- —Copenhagen University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSARS-CoV-2 and COVID-19 Research · Animal Virus Infections Studies · COVID-19 Clinical Research Studies
Background
As of November 2024, the coronavirus disease 2019 (COVID-19) pandemic has produced more than 776 million confirmed human cases and more than 7 million deaths worldwide according to the World Health Organization [1]. Additionally, cases of natural infections have been reported in a wide range of animals including dogs, cats, otters, zoo felids, zoo gorillas, ferrets, and mink [2–7].
Shortly after the COVID-19 pandemic in humans began in early 2020, the first outbreaks of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) on domestic mink farms were identified in the Netherlands in April 2020 [8]. These were followed by outbreaks on Danish mink farms from June 2020 [9, 10]. Thereafter, SARS-CoV-2 infections were reported on mink farms in many countries including the United States, Spain, Sweden, Italy, France, Greece, Lithuania, Canada, Poland, and Latvia [5, 11–16]. Danish mink production faced a significant challenge with outbreaks recorded on 290 mink farms by November 2020. The Danish Government subsequently authorized culling on all mink farms on 4 November 2020 [9]. Prior to the culling, Denmark was a leading contributor to the global mink fur industry with more than 1,000 privately owned farms and approximately 2.5 million breeding mink producing around 12.5 million pelts annually [17]. As of November 2024, there are five mink farms with approximately 35,000 mink in total registered in Denmark [18] and a SARS-CoV-2 surveillance program has been initiated. This is based on collection of throat swabs to test for SARS-CoV-2 at various intervals during the year, as well as monitoring for clinical signs [19].
Reports of observations on SARS-CoV-2 infected mink farms range from mostly asymptomatic in the Danish and Spanish outbreaks [16, 20] to various clinical signs, including watery nasal discharge, respiratory distress, anorexia, mild depression, and coughing in the Dutch, Greek, and American outbreaks [8, 11, 21, 22]. Mortality rates range from minimal (0.45%) in Denmark [20], 2.4% in the Netherlands [8, 22], 8–10% in Greece [21], and up to 55% in the United States [11].
While few cases of zoonotic transmission from white-tailed deer [23] and hamsters [24] have been reported, farmed mink are the species with the most widespread confirmed cases of animal-to-human transmission [10, 25]. They are the only nonhuman species consistently associated with severe illness and death [8, 11, 21, 22] and are the second most commonly documented species exhibiting SARS-CoV-2 infections [26]. However, data regarding the pathology of SARS-CoV-2 infection in mink remain limited. To date, descriptions of SARS-CoV-2 outbreaks in domestic mink in the Netherlands, Greece, Spain, and the USA have reported histopathological similarities to the lung tissue of human COVID-19 patients [11, 16, 21, 22, 27].
This study investigates pulmonary lesions in Danish farm mink (Neogale vison) naturally infected with SARS-CoV-2. We studied a group of asymptomatic mink kits, a group of mink kits that had severe respiratory disease with fatal outcome, and a group of mink kits that were in recovery from infection. Two groups of healthy control mink, matched by age to the three case groups, were also included. All the mink were from farms that were free from Aleutian disease (AD).
Methods
Study design and animals
We investigated samples of lung tissue collected from mink infected with SARS-CoV-2 and uninfected control mink. The investigation was designed as an observational case–control study. The study included three case groups and two control groups, as summarized in Table 1. Case groups were matched with control groups of a similar age from farms with no history of SARS-CoV-2 outbreaks. Additionally, control animals were confirmed negative for active and previous SARS-CoV-2 infection by reverse transcription-quantitative polymerase chain reaction (RT-qPCR) on throat swabs and antibody enzyme-linked immunosorbent assay (ELISA) on blood samples or by SARS-CoV nucleocapsid immunohistochemistry and viral metagenomics. These results are summarized in Table 2. Animals from the subclinical SARS-CoV-2 (SCS), controls July (Con_July), controls November (Con_Nov), and recovering SARS-CoV-2 (RS) groups were randomly selected by the farmer. Animals in the SCS were a subset of the animals sampled on 22 June 2020, in a previous study [10]. Animals in the RS group were a subset of animals from a previously described study, collected on 5 October 2020 [28]. Animals in the SCS, RS, and control groups were euthanized after sedation, in accordance with protocols previously described [29]. All animals in the clinical SARS-CoV-2 (CS) group were found dead on the farm and collected within a few hours postmortem. All animals were stored overnight at 5 °C and necropsies were performed the following day.Table 1. Summary of study groupsGroupGroup nameDate, samplingSamplingDescriptionSubclinical SARS-CoV-2 (n = 14)SCSJune 22, 2020Random, euthanizedAsymptomatic SARS-CoV-2 infected mink kits from a farm with infected personnel [10]. 2 months oldControls, July (n = 10)Con_JulyJuly 2017Random, euthanizedHealthy mink kits from a farm with no history of SARS-CoV-2 infection. 2 months oldClinical SARS-CoV-2 (n = 13)CSOct 5, 2020All dead animals on sampling dateSARS-CoV-2 infected mink kits from an outbreak, exhibiting severe respiratory distress and fatalities. 5 months oldControls, November (n = 30)Con_NovNov 4, 2020Random, euthanizedHealthy mink kits from a farm with no history of SARS-CoV-2 infection. 6 months oldRecovering SARS-CoV-2 (n = 10)RSOct 5, 2020Random, euthanizedMink kits recovering from SARS-CoV-2 infection, collected one month after clinical signs ceased; included reduced feed intake and a few cases of diarrhea and respiratory signs [28]. 5 months oldSARS-CoV-2 severe acute respiratory syndrome coronavirus 2Table 2Results of three SARS-CoV-2 detection methods in all study groupsGroupnSARS-CoV-2 RT-qPCRSARS-CoV-2 antibody ELISASARS-CoV nucleocapsid IHC(positive/tested)(positive/tested)(positive/tested)SCS1411/141/149/14Con_July9––0/9CS1313/13–5/5Con_Nov300/300/300/5RS100/1010/100/10Results from different viral detection methods including SARS-CoV-2 antigen in throat swabs using RT-qPCR, anti-SARS-CoV-2 antibodies in blood samples using ELISA, and SARS-CoV nucleocapsid IHC in lung tissue for the five study groups of SARS-CoV-2 infected and uninfected mink kitsSARS‑CoV‑2* severe acute respiratory syndrome coronavirus 2, ELISA enzyme-linked immunosorbent assay, IHC immunohistochemistry, SCS subclinical SARS-CoV-2, Con_July controls from July, CS clinical SARS-CoV-2, Con_Nov controls from November, RS recovering SARS-CoV-2, RT-qPCR reverse transcription-quantitative polymerase chain reaction^*^Samples from Con_July animals were analyzed using viral metagenomics and no coronavirus was detected
Cardiac blood samples were collected from all animals in the SCS, Con_Nov, and RS groups at the time of euthanasia. Throat swabs were also obtained from all animals in the SCS, CS, Con_Nov, and RS groups. No blood samples were collected from animals in the CS group.
No ethical or laboratory animal permits were required according to Danish legislation. All included animals were destined for euthanasia, due to annual pelting or in accordance with national legislation for disease control.
Necropsy
Standard necropsies were conducted as previously described [30]. The body condition of each carcass was scored on a scale of 1–5, in accordance with relevant guidelines [31]. For each animal, sex, age (juvenile/adult), and gross pathological findings were also recorded.
Histopathology
Samples from various organs were collected during necropsies, including cranial and caudal lung lobes from each mink. Tissue samples were fixed in 10% neutral buffered formalin for 2 d. After trimming, tissues from the cranial and caudal lung lobes were processed routinely for histology: Dehydrated using a graded series of ethanol washes (75%, 80%, 96%, and 99%), then treated with xylene and embedded in paraffin. Next, tissue sections (2–4 µm) were stained with hematoxylin and eosin. Fibrin was visualized using Martius scarlet blue stain, following a modified protocol previously described [32]. Collagen was visualized with Masson’s trichrome stain using an adapted technique detailed earlier [33].
A semiquantitative scoring protocol for pulmonary lesions was developed. In this scoring system, the following six main categories were divided into subcategories: (1) bronchi, (2) glands, (3) bronchioles, (4) vascular system, 95) interstitium, and (6) alveolar lumen. For the bronchi and bronchioles categories, the subcategories included scoring of epithelial damage, luminal secretion and hemorrhage, and submucosal inflammation. For the glands category, the subcategories included scoring of cellular inflammation and degeneration. For the vascular system category, the subcategories included scoring of vasculitis, perivasculitis, perivascular edema, and perivascular accumulation of mononuclear cells. For the interstitium category, the subcategories included scoring of cell infiltration, type II pneumocyte proliferation, and alveolar damage. For the alveolar lumen category, the subcategories included scoring of inflammation, alveolar fibrin, luminal edema, and an overall evaluation of reduction of airspace (for detailed descriptions, see Additional file 1). Each animal was assigned a combined score for each main category. This combined score was the sum of the scores from each of the relevant subcategories. Scores were assigned on an ordinal scale from 0 to 3, with 0 indicating no pathological lesions, and scores of 1 to 3 indicating increasing severity of pathological lesions (a score of 3 indicates the most severe lesions).
Additionally, a dichotomous scale (present/not present) was used to evaluate the presence or absence of the following: (1) hemosiderin in macrophages; (2) giant cells; (3) bacteria; (4) thrombi; (5) neutrophils in lumen or septae; and (6) pulmonary corpora amylacea (PCA).
All tissue sections were single blinded prior to evaluation. The evaluation was performed by the same person in the two case–control groups and the evaluation process was supervised by the same senior pathologists throughout. A Leica DM4 B microscope was used for reading and images were obtained using a Leica DFC7000 T camera and Leica Application Suite X software, version 3.7.6.25997 (Leica Microsystems, Wetzlar, Germany).
Immunohistochemistry
Immunohistochemistry (IHC) markers applied in this study included the following: (1) MAC374G/MAC287 IHC antibody (Bio Rad Laboratories A/S, Copenhagen, Denmark) for identifying macrophages; (2) SARS-CoV nucleocapsid IHC antibody (Nordic Biosite ApS, Copenhagen, Denmark) for identifying SARS-CoV-2 infected cells; and 3) CD3 and CD79 stains (both DAKO Denmark ApS, Glostrup, Denmark) for identifying T and B lymphocytes, respectively. The MAC387, CD3, and CD79 IHC procedures were used on all tissue sections from the CS and Con_Nov groups. The SARS-CoV nucleocapsid IHC procedure was used on tissue samples from the Con_July, SCS, and RS groups, and on five randomly selected tissue sections from the Con_Nov and CS groups. The interpretation of IHC results was based on the presence, localization, and intensity of staining. Positive staining was defined as specific chromogenic signals localized within the expected cellular compartments (e.g., cytoplasmic or nuclear) compared to negative controls. The absence of staining or nonspecific background signal was recorded as negative.
Virology
RT-qPCR, serology evaluation, and sequencing were performed as previously described [10]. Throat swabs were analyzed for SARS-CoV-2 viral RNA by RT-qPCR, in accordance with the procedure described by Corman et al*.* [34]. Blood samples were analyzed for the presence of anti-SARS-CoV-2 antibodies using ELISA (Beijing Wantai Biological Pharmacy Enterprise, Beijing, China), as previously described [10]. Sequencing of individual variants and determination of relationship to the pangolin lineage were performed as previously described [28, 35]. All included farms were confirmed free of antibodies specific for Aleutian mink disease virus (AMDV).
Statistics
For all data, non-parametric tests were employed, as no assumptions of normal distribution were made.
Wilcoxon’s signed-rank test was used to investigate any differences between samples from the cranial and caudal lung lobes in the same individual from the Con_Nov and CS groups; the Mann–Whitney U test was used to investigate differences between scores from the same groups. The Kruskal–Wallis test with Dunn’s correction for multiple comparisons was used to evaluate the sum of scores across all groups included in the study. A P-value < 0.05 was considered statistically significant. All statistical analyses were performed using GraphPad Prism software (ver. 10.0; GraphPad Software, San Diego, CA, USA). When tissue samples were randomly selected for staining, the RAND()-function in Excel software (2016; Microsoft Corporation, Redmond, WA, USA) was used.
Results
Gross pathology
All CS mink kits had enlarged dark-red lungs with increased texture. No other gross lesions were recorded.
In the SCS group, five animals had congested lungs with increased texture and nine animals had no apparent gross lesions.
One animal from the Con_Nov control group had congested lungs. In the RS group, all animals had an enlarged congested spleen and mild steatosis and only one animal did not have lungs with increased texture.
Histopathology
Histopathological examination of lung tissue samples from all animals in the SCS, CS, and RS groups was used to identify lesions; tissue samples from the CS group exhibited the most severe lesions. Representative histological images from the Con_July group and each case group (SCS, CS, and RS) are shown in Fig. 1.Fig. 1. Histological images of SARS-CoV-2 infected and recovering mink. Histological findings in SARS-CoV-2 infected mink with severe clinical disease (a,** b**,** c**) and in recovery (d). a Severe luminal secretion in a bronchi (asterix), submucosal inflammation (arrow), and severe diffuse alveolar damage (HE, bar = 100 µm). b Acute heterogeneous fibrin thrombi in a pulmonary artery consisting of organized fibrin (red) and erythrocytes (yellow) (MSB, bar = 100 µm). c SARS-CoV nucleocapsid positive cells in the alveolar lumen and wall (red) (SARS-CoV nucleocapsid IHC, bar = 25 µm). d Type II pneumocyte proliferation (arrowheads) in (HE, bar = 25 µm). SARS-CoV-2 severe acute respiratory syndrome coronavirus 2, HE hematoxylin and eosin, MSB martius scarlet blue
Regarding the scores, the CS group exhibited significantly higher scores across all main categories compared to the corresponding Con_Nov group. A similar trend was observed in the SCS group when compared to Con_July group; however, there was no significant difference between these two groups for the"vascular system"category. The RS group scores were significantly lower than the CS group scores for all categories except the"interstitium"category, where there was no significant difference. The RS group had significantly higher scores than the Con_Nov group for the"vascular system"and"interstitium"categories but significantly lower scores than the Con_Nov group for the"bronchioles"category. The Con_July group had significantly lower scores than the Con_Nov group for the “bronchi,” “bronchioles,” and “alveolar lumen” categories, but there were no significant differences between these groups for the remaining categories (Fig. 2). One control mink from the Con_July group was excluded due to the presence of histological lesions that indicated bronchopneumonia associated with bacterial infection.Fig. 2. Histopathological scores for six main categories assigned to SARS-CoV-2 infected, uninfected, and recovering mink. Scatter plots with bars showing the mean and standard deviation of the sum of scores assigned to each group: Con_July (n = 9), SCS (n = 14), Con_Nov (n = 30), CS (n = 13), and RS (n = 10). For each category, a semiquantitative scoring protocol was used: a bronchi (epithelial damage, luminal secretion and hemorrhage, and submucosal inflammation); b glands (inflammation and degeneration); c bronchioles (epithelial damage, luminal secretion and hemorrhage, and submucosal inflammation); d vascular system (vasculitis, perivasculitis, perivascular edema, and perivascular accumulation of mononuclear cells); e interstitium (cell infiltration, type II pneumocyte proliferation, and alveolar damage); and f alveolar lumen (inflammation, fibrin, hemorrhage, edema, and overall reduction of airspace). Statistical analysis was performed using the Kruskal–Wallis test and Dunn’s correction for multiple comparisons. A P-value < 0.05 was considered statistically significant (ns ≥ 0.05, * < 0.05, ** < 0.01,*** < 0.001). ns not significant, SARS‑CoV‑2 severe acute respiratory syndrome coronavirus 2, Con_July healthy controls from July, SCS subclinical SARS-CoV-2, Con_Nov healthy controls from November, CS clinical SARS-CoV-2, RS recovering from SARS-CoV-2
There was significantly more intracellular hemosiderin in macrophages from the CS group (11/13; 85%) than in those from the Con_Nov (11/30; 37%) and RS (3/10; 30%) groups, whereas no intracellular hemosiderin was observed in macrophages from either the SCS or Con_July group. Giant cells were observed in the SCS (2/14; 14%), Con_Nov (8/30; 27%), and CS (9/13; 69%) groups, with significantly more being present in the CS group than in the Con_Nov group. Bacteria, mainly cocci, were only observed in three cases, with no significant differences between the Con_Nov (1/30; 3%) and CS (2/13; 15%) groups. Thrombi were observed in the SCS (7/14; 50%), CS (8/13; 62%), and RS (2/10; 20%; Fig. 1) groups but not in the corresponding control groups (Con_July and Con_Nov). There was no significant difference in thrombi formation between the CS and RS groups. There were significantly more neutrophils in the alveolar lumen or septae of the SCS (7/14; 50%), CS (12/13; 92%), and RS (6/10; 60%) groups than in the corresponding Con_July (0%) and Con_Nov (2/30; 7%) control groups. Finally, there were round, solitary, acellular and eosinophilic structures in the alveolar lumen that resembled PCA in all groups, but these were significantly more abundant in the CS and Con_Nov groups than in the RS group. Significant differences between the groups are shown in Fig. 3.Fig. 3. Histological characteristics of SARS-CoV-2 infected, uninfected, and recovering mink. Stacked bar chart showing the histological characteristics of the five study groups: Con_July (n = 9), SCS (n = 14), Con_Nov (n = 30), CS (n = 13), and RS (n = 10). Each graph represents a particular characteristic including: a hemosiderin in macrophages, b giant cells, c bacteria, d thrombi, e neutrophils in lumen or septae, and f pulmonary corpora amylacea. Each bar corresponds to a group, with segments indicating the proportions that are “present” (represented in orange) and “not present” (represented in blue). Fisher’s exact test was used to evaluate differences in outcomes between the groups. A P-value < 0.05 was considered statistically significant (ns ≥ 0.05, * < 0.05, ** < 0.01, *** < 0.001). ns not significant, SARS‑CoV‑2 severe acute respiratory syndrome coronavirus 2, Con_July controls from July, SCS subclinical SARS-CoV-2, Con_Nov controls from November, CS clinical SARS-CoV-2, RS: recovering from SARS-CoV-2
In addition to the scores obtained from the semiquantitative protocol, there were notable differences in the particular histological lesions present in the different groups, including vasculitis, perivasculitis, cell infiltration in the alveolar interstitium, and type II pneumocyte proliferation. For example, mild (9/13; 69%) to moderate (3/13; 23%) vasculitis was exclusively observed in the CS group.
Perivasculitis was only observed in the CS group, with all animals displaying some degree of perivasculitis (mild, (1/13;8%); moderate, (2/13;15%); or severe, (10/13;77%)). All groups exhibited perivascular accumulation of mononuclear cells, especially the RS group. Varying degrees of perivascular edema were observed in the SCS and CS groups.
Cell infiltration of mononuclear cells or neutrophil granulocytes in the alveolar interstitium was observed in the SCS, CS, and RS groups, with 12 out of 13 (92%) of the tissue sections in the CS group showing a diffuse pattern of cell infiltration. The infiltration patterns in the SCS and RS groups were milder, including focal (4/14 (29%) and 1/10 (10%), respectively), multifocal (3/14 (21%) and 2/10 (20%), respectively), and diffuse (5/14 (36%) and 3/10 (30%), respectively) patterns.
Type II pneumocyte proliferation was infrequent and included focal (3/14 (21%) in SCS and 2/10 (20%) in RS) and multifocal (1/14 (7%) in SCS and 4/10 (40%) in RS) proliferation. No proliferation was observed in the CS or control groups. Diffuse alveolar damage was observed to varying degrees in the SCS, CS, and RS groups.
In both the Con_Nov and RS groups, there was multifocal nodular accumulation of mononuclear cells at several locations. Most frequently, the nodules were peribronchiolar and associated with bifurcations; however, in 8 out of 30 animals from the Con_Nov group (27%) and 6 out of 10 animals from the RS group (60%), focal mononuclear nodules were present in the interstitia of the alveolar tissue (Fig. 4).Fig. 4. Secondary lymphoid tissue in an apparently healthy mink. Lung tissue from an apparently healthy mink, showing aggregates of lymphocytes (a), predominantly B cells (b), and T cells (c) in the surrounding regions (all indicated by arrows). a hematoxylin and eosin; b CD79 stain; c CD3 stain. Scale bars: 50 µm for all images
Cranial and caudal lung sections from the CS and Con_Nov groups were evaluated. Upon unblinding, the scores for the cranial and caudal lung lobes of each animal were compared. Because there were no statistically significant differences between these scores, each animal was assigned the higher score from the two lobes in each subcategory.
All lung sections from the RS group and six randomly selected sections from the Con_Nov group were stained with Masson’s trichrome to evaluate lung fibrosis. We observed no differences in fibrosis between the two groups.
Immunohistochemistry
The MAC387 IHC evaluation revealed macrophages in lung tissue from both groups. In the CS group, macrophages infiltrated the vascular wall in cases of vasculitis and often when interstitial inflammation was observed. In the Con_Nov group, there were macrophages at various locations within the lung. Specifically, macrophages were found multifocally infiltrating the alveolar interstitium (up to two cells per alveolar septum), in small numbers in the lumina of the bronchi and bronchioles (and sporadically subepithelially), and in lamina propria of both bronchi and bronchioles.
In both the SCS and CS groups, SARS-CoV nucleocapsid positive cells were observed in the bronchi and bronchiolar epithelium, in the alveolar wall, and in the alveolar lumen. In general, the concentration of SARS-CoV nucleocapsid positive pneumocytes and macrophages was higher near the bronchi and bronchioles than in the peripheral pulmonary tissue. SARS-CoV nucleocapsid positive epithelial cells in the bronchi and bronchioles retained normal morphology with intact cilia in the pseudostratified epithelium and were attached to the basement membrane. SARS-CoV nucleocapsid positive pneumocytes were either located in the alveolar wall or desquamated to the alveolar lumen. SARS-CoV nucleocapsid positive mononuclear cells were found in the lumen of alveoli, bronchi and bronchioles, and in the alveolar interstitium. MAC387 staining indicated that the positive cells in the alveolar lumen were probably macrophages. In three samples from the CS group, glandular cells were SARS-CoV nucleocapsid positive. No endothelia in any vessels or leukocytes involved in vasculitis were positive. No positive cells were associated with thrombi formation. No SARS-CoV nucleocapsid positive cells were observed in the Con_July, Con_Nov, or RS group.
CD3 and CD79 IHC analysis revealed that nodular accumulations of mononuclear cells observed in alveolar interstitia in the Con_Nov group consisted of B lymphocytes at the center and T lymphocytes at the periphery and resembled secondary lymphoid tissue. There was perivascular lymphocytic cuffing in both the Con_Nov and CS groups, predominantly involving T lymphocytes. In the CS group, however, this perivascular cuffing consisted of several layers of T lymphocytes.
Virology
A summary of the results from three different SARS-CoV-2 detection methods including SARS-CoV-2 antigen in throat swabs using RT-qPCR, anti-SARS-CoV-2 antibodies in blood samples using ELISA, and SARS-CoV nucleocapsid IHC on pulmonary histology sections is shown in Table 2.
Discussion
Overall, the histopathological evaluation in this study revealed extensive pulmonary pathology in all case groups. The pathological findings were significantly more severe in the group of mink that developed clinical signs of severe acute respiratory distress and died due to SARS-CoV-2 infection. However, noticeable lesions were also observed in the group of mink that exhibited no clinical disease, as well as in those that were in recovery from a SARS-CoV-2 outbreak. In a previous study, mild to moderate bronchointerstitial pneumonia was reported in asymptomatic SARS-CoV-2-infected mink during an outbreak on a Spanish farm [16]. Additionally, another study found that one in three mink farms with SARS-CoV-2 infections showed no clinical signs of disease [20].
The mandatory SARS-CoV-2 surveillance program in Denmark, as of 2025, focuses on detecting infections linked to clinical disease in mink farms. Farms are subject to public monitoring if there are any suspicions of SARS-CoV-2 infection based on observed clinical signs. Additionally. the program mandates the collection of throat swabs to test for virus from 5 deceased animals every 3 months, from 15 animals no more than 10 days before pelting, and from 60 animals prior to import to Denmark when new animals are purchased abroad [19].
Originally native to the North American continent, the mink is a solitary carnivore with territorial, crepuscular, and nocturnal habits [36]. Its inclination toward solitude and masking signs of pain mean that assessing disease symptoms in mink may be difficult, similar to other domestic carnivores such as cats [37] and ferrets [38]. Good health and freedom from injury are welfare quality principles that have been included in the WelFur assessment program, which has been implemented at mink farms across the European continent and is maintained by impartial assessors. In this context, good health is defined by the following welfare quality criteria: absence of injuries, skin lesions or other injuries to the body; freedom from disease, risk of mortality, diarrhea, lameness, impaired movement, or pain produced as a result of management procedures [39]. A previous study in the Netherlands suggested that outbreaks of SARS-CoV-2 on some mink farms may be missed due to a lack of clinical signs [22]. In this study, the severity of pathology observed in asymptomatic SARS-CoV-2-infected mink, as well as those that were in recovery from the disease, suggests that a surveillance strategy based on testing four times a year and monitoring clinical signs may not adequately meet welfare needs. First, infected mink with subclinical disease may never be tested under the current surveillance system; unless they die from other diseases, they will not be included in the mandatory testing every three months. Second, testing deceased mink with throat swabs every three months appears insufficient, as they may test negative just one month after the initial detection of an outbreak, as previously reported [28]. Danish animal welfare legislation requires that individuals responsible for production animal care monitor each animal at least once daily. While mink farmers routinely observe their animals as part of standard management practices, systematic studies on the presentation of clinical diseases, including signs of pain-related behaviors, have not been reported in mink, unlike other more commonly studied animal models or production animals. This highlights the need for further research to establish reliable behavioral and physiological markers of disease and discomfort in this species. Consequently, SARS-CoV-2 infection on a farm could be overlooked in the present surveillance program. Furthermore, the public health implications of undetected SARS-CoV-2 infection on mink farms should be investigated further. The evidence of the mink-adapted SARS-CoV-2 Cluster 5 variant spilling back into the local community [8, 10, 40] underlines a need for improved biosecurity measures on mink farms in general. Although the mutations in the Cluster 5 variant negatively impacted the replication of the virus in human cells [41] the spread of this variant into the human community underscores the possibility of mink serving as reservoirs of new virus variants with unknown pathogenicity in the future.
Additionally, perivasculitis was frequently observed in the mink group that succumbed to SARS-CoV-2 infection, which corroborates reports from previous studies on mink [11, 16, 22, 42]. Notably, perivascular accumulations consisting of multiple layers of mononuclear cells were also consistently present in the control groups. A previous study found that mononuclear vascular cuffing was common in Danish mink kits, but no infectious cause was identified [43]. This may be similar to the perivascular lymphocytic cuffing documented in clinically unremarkable domestic cats [44]. Consequently, the term “perivasculitis” may be misleading and apparent lesions in lung tissues from farmed mink could represent the proliferation of pre-existing perivascular lymphatic tissue rather than true perivasculitis. Additionally, nodular structures resembling secondary lymphoid tissue, featuring a central B follicle and an outer T-lymphocyte zone, were consistently observed in the control group from November and in the group of mink in recovery from SARS-CoV-2, in which the kits were 6 months and 5 months old, respectively. These lymphoid aggregates share features with the bronchus-associated lymphoid tissue (BALT) described in other species, including cats and humans [45–47]. Additionally, such lymphoid aggregates were not present in younger mink kits (2 months old), which is consistent with studies on rats that report BALT first developing only after 2 weeks postnatally [46]. Although a general understanding of BALT may be extrapolated from related species, detailed studies in mink have not been published. Future research is necessary to investigate whether BALT plays a role in controlling respiratory infections and could be important for evaluation of environmental responses and farming practices.
The presence of lymphocytic aggregates in the lungs of healthy mink reported in this study warrants further investigation to determine any functional role. In addition, previous studies have suggested that mink could be used as a model for human COVID-19 studies [27, 42, 48] and the findings reported here emphasize the need for more detailed descriptions of mink lungs to evaluate suitability.
Furthermore, histological evidence for bacteria in the lungs of mink that died during SARS-CoV-2 outbreaks was minimal, suggesting that mortality is more likely due to damage caused by SARS-CoV-2. By contrast, secondary bacterial infections were reportedly responsible for mortality in Danish mink that succumbed to influenza infections [49].
In this study, proliferation of type II pneumocytes was predominantly observed in the group of mink in recovery from SARS-CoV-2, whereas the two groups of mink with active infections exhibited minimal proliferation in only a few animals. When assessing type II pneumocyte proliferation in mink lungs, AD status must be considered because the early stages of infection with AMDV can cause type II pneumocyte hyperplasia [50]. The mink included in this study were from farms that had been confirmed as free from AD before and after sampling. During the COVID-19 pandemic when SARS-CoV-2 outbreaks were occurring on mink farms worldwide, AD was widespread on mink farms globally [51]. In Denmark, AD is a notifiable disease and all mink breeders must test their animals several times every year. Strict testing regimes have successfully reduced outbreaks of AD on farms [52]. In a previous study conducted in the Netherlands, moderate to severe proliferation of type II pneumocytes was observed in adult mink from four different SARS-CoV-2-infected farms [22]. All these farms had tested positive for AMDV during the past 10 years, but none showed clinical signs of Aleutian disease. In another study from a SARS-CoV-2 farm outbreak in Utah, infrequent type II pneumocyte proliferation in 20 SARS-CoV-2 infected mink from an AD-positive farm was reported [11]. In two experimental studies of SARS-CoV-2 infected mink, type II pneumocyte proliferation was reported to moderate and varying degrees [42, 53], although the AD status of the mink was not specified. In another experimental study on 11 SARS-CoV-2 infected mink and 6 control mink, type II pneumocyte proliferation was not observed and, interestingly, these mink were AD negative [54]. The previous findings together with our observations, showing that proliferation predominately occurs 1 month after active SARS-CoV-2 infection, suggest that type II pneumocyte proliferation in SARS-CoV-2 infected mink may be a sign of post-infection recovery rather than active infection.
This study facilitated a comparative analysis of pulmonary histological lesions in SARS-CoV-2 infected mink kits with and without clinical disease, as well as mink in recovery from the disease. The inclusion of age matched controls was a marked strength of this study, given the lack of descriptions of normal mink pulmonary anatomy found in previous reports.
In all study groups, PCA were observed in the alveolar lumen of many animals. The presence of PCA in humans with pneumonia has been documented for decades [55–57], and PCA also occur in alveolar spaces in sheep with chronic and acute pneumonia [58, 59]. In general, PCA are considered incidental findings [60] and their specific function remains unknown [61], although their presence may indicate previous injury or pre-existing lesions wherein cellular debris and secretions have accumulated [56]. To our knowledge, this is the first study to document the presence of PCA in mink, and our observations suggest that the presence of PCA may be a common incidental finding in mink lungs.
Conclusion
Severe pulmonary lesions were observed in SARS-CoV-2 infected mink kits with subclinical and clinical disease and in kits in recovery from infection. These lesions included epithelial damage, luminal secretion and hemorrhage, subepithelial inflammation of the bronchi and bronchioles, diffuse alveolar damage with hyaline membrane formation, alveolar hemorrhage, mononuclear cell infiltration of interstitia, and the formation of fibrin, edema, and thrombi. These findings suggest that the impact of SARS-CoV-2 farm outbreaks may be more serious than records of clinical signs indicate. The current surveillance system on Danish mink farms does not effectively detect repeated SARS-CoV-2 outbreaks that show no clinical signs or cause no mortality, which can lead to unnoticed welfare issues. The severity of the observed lesions reveal hidden health and welfare challenges in mink, underscoring the need for improved prevention measures, surveillance and understanding of long-term impact of SARS-CoV-2 infection in mink.
Supplementary Information
Additional file 1. Semiquantitative scoring protocol for evaluating minklung tissue. Semiquantitative scoring protocol for histological evaluation of pulmonary tissue from farm mink infected with SARS-CoV-2. The protocol comprises six main categories with subcategories.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1World Health Organization. Coronavirus disease (COVID-19)—situation reports [Internet]. 2024 [cited 2024 Nov 11]. https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports
- 2World Organization for Animal Health. SARS-Co V-2 [Internet]. WOAH—World Organisation for Animal Health. 2024 [cited 2024 Nov 11]. https://www.woah.org/en/disease/sars-cov-2/
- 3Statistics Denmark. Fakta om minkbranchen i Danmark [Internet]. Danmarks stat. 2021 [cited 2024 Nov 19]. https://www.dst.dk/da/Statistik/nyheder-analyser-publ/bagtal/2020/2020-10-28-fakta-om-minkbranchen-i-Danmark
- 4Danish Veterinary and Food Administration. CHR [Internet]. 2024 [cited 2024 Nov 11]. https://chr.fvst.dk/chri/faces/frontpage
- 5Chriél M, Jensen TH, Hjulsager C, Larsen LE, Jørgensen PH, Harslund JLF, et al. Consequences of outbreaks of influenza A virus in farmed mink (Neovison vison) in Denmark in 2009 and 2010. In: Larsen PF, Møller SH, Clausen T, Hammer AS, Lássen TM, Nielsen VH, et al., editors. Proceedings of the Xth International Scientific Congress in fur animal production. Wageningen: Academic Publishers; 2012. p. 186–9.