Impact of embryo size on apoptosis in C. elegans
Madiha Javeed Ghani, Jens Van Eeckhoven, Barbara Conradt

TL;DR
This study shows that smaller cells in C. elegans embryos are more likely to undergo programmed cell death.
Contribution
The study demonstrates that cell size directly influences apoptosis in genetically manipulated C. elegans embryos.
Findings
Knock-down of C27D9.1 increases inappropriate cell survival in apoptosis-compromised embryos.
Reducing embryo size via ima-3 RNAi partially suppresses inappropriate cell survival.
Smaller cells are more likely to undergo apoptosis in C. elegans embryos.
Abstract
During C. elegans development 131 somatic cells reproducibly undergo programmed cell death. Many of these 131 cells ‘programmed to die' are the smaller daughter of a neuroblast that divides asymmetrically and die through apoptosis. To determine whether cell size impacts the ability of cells programmed to die to undergo apoptosis, we increased or decreased embryo size by RNA interference-mediated knock-down of the genes C27D9.1 or ima-3 , respectively. We found that in apoptosis-compromised genetic backgrounds, C27D9.1 ( RNAi ) enhances and ima-3 ( RNAi ) partially suppresses inappropriate survival of cells programmed to die. This supports the notion that in C. elegans embryos, an increase in cell size compromises and a decrease in cell size promotes the ability of cells programmed to die to undergo apoptosis.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
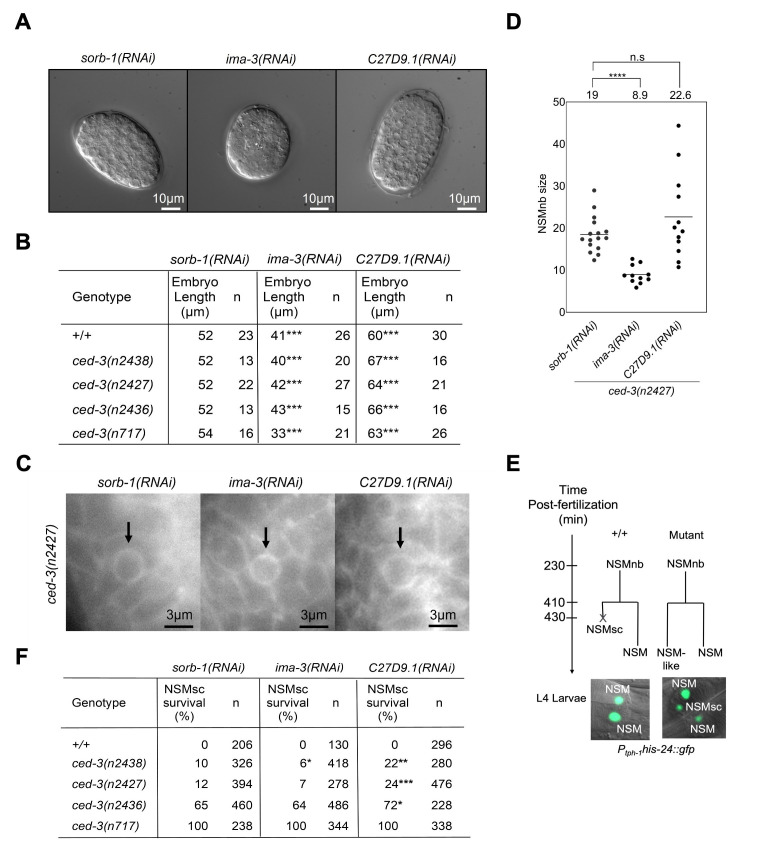 Figure 1
Figure 1|
Strain |
Genotype |
Source |
|
|
|
Yan et al., 2013 |
|
|
|
This study |
|
|
|
Chakraborty et al., 2015 |
|
|
|
Chakraborty et al., 2015 |
|
|
|
Yan et al., 2013 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsReproductive Biology and Fertility
Description
Many of the 131 cells that die during * Caenorhabditis elegans * development are the smaller daughter of a neuroblast (referred to as ‘mother' cell) that divides asymmetrically by both fate and size (Sulston and Horvitz, 1977; Sulston et al., 1983). Loss-of-function mutations of * pig-1 * , which encodes a PAR-1-like kinase orthologous to human MELK (maternal embryonic leucine zipper kinase), cause mother cells to divide symmetrically by size, thereby increasing the physical size of the normally smaller daughter cell i.e. the cell programmed to die (Cordes et al., 2006). In addition, these mutations cause inappropriate survival of cells programmed to die, indicating that they compromise the ability of these cells to undergo apoptosis (Cordes et al., 2006). Conversely, gain-of-function mutations of * ect-2 * , which encodes the * C. elegans * ortholog of the human RhoGEF Ect2 (Epithelial Cell Transforming 2), cause mother cells to divide more asymmetrically by size, thereby decreasing the physical size of the already smaller daughter cell i.e. the cell programmed to die (Sethi et al., 2022). In addition, these * ect-2 * gain-of-function mutations partially suppress the inappropriate survival of cells programmed to die caused by partial loss-of-function mutations of * ced-3 * (which encodes a caspase required for apoptosis in * C. elegans * ), indicating that they promote the ability of these cells to undergo apoptosis (Sethi et al., 2022). However, at least in the NSM neuroblast lineage ( Figure 1 ; see below), mutations in both * pig-1 * and * ect-2 * also affect the unequal inheritance of cellular components during asymmetric mother cell division (Wei et al., 2020; Sethi et al., 2022). For instance, during asymmetric NSM neuroblast division, the loss of * pig-1 * disrupts the unequal segregation - into the surviving NSM – of the anti-apoptotic transcription factor CES-1 , an ortholog of human SCRT1 and 2 (scratch family transcriptional repressor). As a result, the NSM sister cell, which is programmed to die, now inherits this anti-apoptotic factor (Wei et al., 2020). For this reason, it has been difficult to discern whether the inappropriate survival of cells programmed to die observed in * pig-1 * loss-of-function mutants is the result of their increased size, the inappropriate inheritance of anti-apoptotic factors or both. The rationale for the study described below was to change the physical size of cells programmed to die using genetic tools other than mutations that affect the ability of mother cells to divide asymmetrically and to determine the impact of this change on the ability of the resulting cells to undergo apoptosis.
The * C. elegans * genes * ima-3 * and * C27D9.1 * encode an importin alpha protein and a protein predicted to be involved in fucosylation, respectively (Geles and Adams, 2001; WormBase). The knock-down by RNA interference (RNAi) of * ima-3 * and * C27D9.1 * decreases or increases embryo size, respectively (Sönnichsen et al., 2005; Hara and Kimura, 2009; Weber and Brangwynne, 2015; Hubatsch et al., 2019). Using * sorb-1 (RNAi) * as a control (Haeussler et al., 2021), we confirmed this in wild type and in different * ced-3 * caspase mutant backgrounds, in which apoptosis is compromised with increasing degrees i.e. * ced-3 ( n2438 ) * , * ced-3 ( n2427 ) * , * ced-3 ( n2436 ) * and * ced-3 ( n717 ) * ( ** Figure 1A, B ** ; see below) (Ellis and Horvitz, 1986; Shaham et al., 1999). In addition, by measuring the size of the NSM neuroblast just prior to its division (metaphase; performed in the * ced-3 ( n2427 ) * background), we found that * ima-3 (RNAi) * reduces NSM neuroblast size whereas * C27D9.1 (RNAi) * increases NSM neuroblast size albeit not significantly ( ** Figure 1C, D ** ). The two NSM neuroblasts divide and each gives rise to a larger NSM, which survives, and a smaller NSM sister cell (NSMsc), which is programmed to die through apoptosis ( ** Figure 1E ** ). Next, using a reporter that visualizes the two NSMs as well as inappropriately surviving NSM sister cells in L4 larvae (P * _ tph-1 _ his-24 ::gfp * ; transgene bcIs66 ) (Yan et al., 2013) ( ** Figure 1E ** ), we determined ‘NSM sister cell survival' (% NSMsc survival) in * ima-3 (RNAi) * and * C27D9.1 (RNAi) * larvae in a wild-type background (+/+) and in the background of * ced-3 ( n2438 ) * , * ced-3 ( n2427 ) * , * ced-3 ( n2436 ) * or * ced-3 ( n717 ) * , respectively. In a wild-type background, NSM sister cells always die (0% NSMsc survival), and we found that * ima-3 * ( RNAi ) has no effect on this ( ** Figure 1F, ** * sorb-1 (RNAi) * vs * ima-3 (RNAi) * ). In the background of the weak * ced-3 * loss-of-function mutations * ced-3 ( n2438 ) * and * ced-3 ( n2427 ) * , * ima-3 (RNAi) * suppresses NSM sister cell survival from 10% to 6% and from 12% to 7%, respectively. Therefore, in these mutant backgrounds, * ima-3 (RNAi) * causes increased NSM sister cell death, which indicates that * ima-3 * has ‘anti-apoptotic' function. However, * ima-3 (RNAi) * has no effect on NSM sister cell survival in the background of the stronger * ced-3 * loss-of-function mutation * ced-3 ( n2436 ) * or the putative * ced-3 * null mutation * ced-3 ( n717 ) * , in which 65% or 100% of the NSM sister cells inappropriately survive ( ** Figure 1F ** ). In the wild-type background, * C27D9.1 * ( RNAi ) also has no effect on NSM sister cell survival ( ** Figure 1F, ** * sorb-1 * ( RNAi ) vs * C27D9.1 * ( RNAi )). However, we found that * C27D9.1 (RNAi) * enhances NSM sister cell survival in * ced-3 ( n2438 ) * , * ced-3 ( n2427 ) * and * ced-3 ( n2436 ) * animals but not in * ced-3 ( n717 ) * animals, in which NSM sister cell survival is already 100% ( ** Figure 1F ** ). Therefore, in these mutant backgrounds, * C27D9.1 (RNAi) * causes decreased NSM sister cell death, which indicates that * C27D9.1 * has ‘pro-apoptotic' function.
At least to our knowledge, * ima-3 * and * C27D9.1 * have not been implicated in apoptosis or non-apoptotic forms of cell death. While we cannot rule out that they have not yet described roles in cell death, the simplest explanation for our results is that * ima-3 * is anti-apoptotic and * C27D9.1 * pro-apoptotic as a result of their opposing roles in embryo size and NSM neuroblast size and therefore, presumably, the size of the NSM sister cell. Specifically, we propose that – at least in the background of partial loss-of-function mutations of * ced-3 * caspase - the * ima-3 (RNAi) * -induced reduction in cell size promotes and the * C27D9.1 (RNAi) * -induced increase in cell size compromises the ability of the NSM sister cell to undergo apoptosis. Hence, our results support the notion that in * C. elegans * embryos, cell size and apoptosis inversely correlate. Our results also suggest that the effects of * pig-1 * and * ect-2 * mutations on the ability of the NSM sister cell to undergo apoptosis are in part due to their impact on NSM sister cell size. Finally, our finding that in a wild-type background, * ima-3 (RNAi) * and * C27D9.1 (RNAi) * do not affect cell death in the NSM neuroblast lineage suggests that the * ima-3 (RNAi) * -induced reduction in cell size is not sufficient to cause inappropriate apoptosis (of the NSM neuroblast or the NSM) and that the * C27D9.1 (RNAi) * -induced increase in cell size is not sufficient to cause the NSM to inappropriately survive; however, we cannot exclude that the * ima-3 (RNAi) * -induced reduction in cell size and the * C27D9.1 (RNAi) * -induced increase in cell size were not large enough to impact apoptosis.
Methods
Strain maintenance and genetics
Strains were maintained at 15°C or 20°C on Nematode Growth Media (NGM) plates and fed with OP50 * Escherichia coli * bacteria according to standard protocols (Brenner, 1974). Bristol N2 strain was used as wild type. The following transgenes and alleles were used in this study: LGIII bcIs66 ( * P _ tph-1 _ his-24 ::gfp * ) (Yan et al., 2013); LGIV * ced-3 ( n2438 ), ced-3 ( n2427 ), ced-3 ( n2436 ), ced-3 ( n717 ) * (Ellis and Horvitz, 1986; Shaham et al., 1999); LGV * ltIs44 * ( * P _ pie-1 _ mCherry:: ph ^PLCΔ^ * ) (Audhya et al., 2005). LGII bcIs104 ( * P pie1 gfp:: tac-1 * ) (Bellanger and Gonczy, 2003). RNAi clones from the Ahringer RNAi library were used for the following genes (Kamath and Ahringer, 2003; Kamath et al., 2003): * ima-3 * ( * F32E10.4 * ) (Sönnichsen et al., 2005), * C27D9.1 * (Sönnichsen et al., 2005), * sorb-1 * ( * Y45F10D.13 * ) (Haeussler et al., 2021), and * pop-1 * ( * W10C8.2 * ) (Rueyling et al., 1998). Throughout our studies, we used information and tools available on WormBase ( https://wormbase.org/#012-34-5 ) (Fire et al., 1998; Sternberg et al., 2024).
RNA interference ( RNAi) by feeding
All RNAi bacterial clones were picked from the Ahringer library (Kamath and Ahringer, 2003; Kamath et al., 2003). This library is distributed by Source BioScience Ltd. ( https://sourcebioscience.com ). To verify the clones, genomic fragments of the targeted genes cloned into the L4440 vector were amplified by PCR using L4440-F ( TGGATAACCGTATTACCGCC ) and L4440-R ( GTTTTCCCAGTCACGACGTT ) oligos, sequenced and sequences verified using the WormBase BLAST tool ( https://wormbase.org/tools/blast_blat ). Verified RNAi bacterial clones were inoculated in 200 µL of LB broth containing 100 μg/mL carbenicillin and grown overnight at 37°C in a shaking incubator. 100 µL of each bacterial culture was seeded onto NGM plates containing 6 mM IPTG and 100 µg/mL carbenicillin. Plates were kept at room temperature overnight in the dark. RNAi by feeding was performed by transferring L4 larvae onto seeded IPTG plates and kept at 20°C for 24-96 hours depending on the gene (Timmons and Fire, 1998; Burton et al., 2011). F1 embryos and/or L4 larvae were harvested for embryo or cell size analyses and for determining NSM sister cell survival. * sorb-1 (RNAi) * was used as negative control (Haeussler et al., 2021) and * pop-1 (RNAi) * as positive control (Rueyling et al., 1998) for RNAi. Loss of * pop-1 * resulted in nearly 100% dead embryos.
Microscopy
Measurements of embryo length and NSM neuroblast (NSMnb) size
24-48 hours post-RNAi feeding, embryos were harvested for embryo size measurements. Embryos were mounted on a glass slide with 2% agarose pads containing M9 buffer. For measuring the lengths of the embryos, Nomarski Z-stacks were taken. The central Z-stack was used to measure the length of each embryo. Using a line selection tool in ImageJ software, a straight line was drawn in the centre of the embryo from anterior to posterior (egg shell), and the length was measured by using ImageJ software (https://imagej.net/ij/) (Schneider et al., 2012; v1.54m). Average embryo length was calculated for each RNAi condition.
The size of the NSMnb under different RNAi conditions was estimated using the * P _ pie-1 _ mCherry::ph ^PLCΔ^ * plasma membrane reporter ( * ltIs44 * ) (Audhya et al., 2005) essentially as described (Sethi et al., 2022). The NSMnb was identified based on its morphology and position and its size was estimated when it had reached metaphase. A Z-stack (0.5 μm step size) was taken through it, and a region of interest (ROI) was drawn around the plasma membrane of the NSMnb in all Z-slices to determine NSMnb area. To estimate NSMnb size, the sum was taken of all NSMnb areas.
A Zeiss Axio Imager.M2 equipped with Differential Interference Contrast (DIC) and epifluorescence was used for imaging.
Measurement of % NSM sister cell (NSMsc) survival
72-96 hours post-RNAi feeding start, L3/L4 larvae were picked to score inappropriate NSMsc survival using the * P _ tph-1 _ his-24 ::gfp * reporter ( bcIs66 ). Worms were mounted on a glass slide with 2% agarose pads containing sodium azide in M9 buffer (25 mM). NSMsc survival was determined by counting GFP-positive cells in the anterior pharynx as previously described (Thellmann et al., 2003; Yan et al., 2013; Sethi et al., 2022). A Zeiss Axio Imager.M2 equipped with Differential Interference Contrast (DIC) and epifluorescence was used for imaging.
Statistical analyses
Statistical analyses were performed using the R statistical software package. For comparing the embryo sizes, a linear model containing all genotypes and RNAi conditions was constructed (R Core Team, 2024; v4.4.1), and its residuals were visually inspected for deviations from normality and homoscedasticity of variance. Differences between relevant RNAi treatments ( * sorb-1 (RNAi) * controls vs respective RNAi treatments) within genotypes were then tested post-hoc using the least square means method in the emmeans package (Lenth, 2024; v1.10.5). To compare statistical significance of NSMnb size under different RNAi conditions, Mann-Whitney U-tests were used. One sided Fisher's exact tests were used to determine statistical significance of % NSMsc survival under all RNAi conditions. All p-values were corrected for multiple testing using false discovery rate. (Benjamini and Hochberg, 1995). Significance level (α) was set at *p * < 0.05 for all tests.
Reagents
**: **
Complete genotypes of strains used in this study.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Audhya Anjon Hyndman Francie Mc Leod Ian X. Maddox Amy S. Yates John R. Desai Arshad Oegema Karen 20051024 A complex containing the Sm protein CAR-1 and the RNA helicase CGH-1 is required for embryonic cytokinesis in Caenorhabditis elegans The Journal of Cell Biology 17121540-814026727910.1083/jcb.20050612416247027 PMC 2171198 · doi ↗ · pubmed ↗
- 2Bellanger Jean-Michel Gönczy Pierre 200391 TAC-1 and ZYG-9 Form a Complex that Promotes Microtubule Assembly in C. elegans Embryos Current Biology 13170960-98221488149810.1016/s 0960-9822(03)00582-712956950 · doi ↗ · pubmed ↗
- 3Benjamini Yoav Hochberg Yosef 199511 Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing Journal of the Royal Statistical Society Series B: Statistical Methodology 5711369-741228930010.1111/j.2517-6161.1995.tb 02031.x · doi ↗
- 4Brenner S 197451 THE GENETICS OF CAENORHABDITIS ELEGANS Genetics 7711943-2631719410.1093/genetics/77.1.714366476 PMC 1213120 · doi ↗ · pubmed ↗
- 5Burton Nick O. Burkhart Kirk B. Kennedy Scott 20111121 Nuclear RN Ai maintains heritable gene silencing in Caenorhabditis elegans Proceedings of the National Academy of Sciences 108490027-8424196831968810.1073/pnas.1113310108 PMC 324181922106253 · doi ↗ · pubmed ↗
- 6Chakraborty Sayantan Lambie Eric J. Bindu Samik Mikeladze-Dvali Tamara Conradt Barbara 20151210 Engulfment pathways promote programmed cell death by enhancing the unequal segregation of apoptotic potential Nature Communications 612041-172310.1038/ncomms 10126 PMC 468211726657541 · doi ↗ · pubmed ↗
- 7Cordes Shaun Frank C. Andrew Garriga Gian 2006715 The C. elegans MELK ortholog PIG-1 regulates cell size asymmetry and daughter cell fate in asymmetric neuroblast divisions Development 133141477-91292747275610.1242/dev.0244716774992 · doi ↗ · pubmed ↗
- 8Ellis H 1986328 Genetic control of programmed cell death in the nematode C. elegans Cell 4460092-867481782910.1016/0092-8674(86)90004-83955651 · doi ↗ · pubmed ↗