The Curli Accessory Protein CsgF of Salmonella Typhimurium Influences the in vitro Aggregation of Human Islet Amyloid Polypeptide
Osmar Meza-Barajas, Clayton Connelly, Alejandra Lopez, Isamar Aranda, Ashwag Binmahfooz, Allison Newell, Sajith Jayasinghe

TL;DR
A bacterial protein called CsgF from Salmonella prevents the clumping of a human amyloid protein in laboratory experiments.
Contribution
This study reveals a novel role for CsgF in inhibiting human islet amyloid polypeptide aggregation.
Findings
CsgF prevents hIAPP aggregation as shown by no increase in Thioflavin T fluorescence.
The C-terminal half of CsgF is sufficient to inhibit hIAPP aggregation.
The N-terminal 65 residues of CsgF are not required for this inhibitory effect.
Abstract
The Curli secretion gene product F (CsgF) is a critical component of the assembly of Curli, proteinaceous filaments, found on the outer surface of gram-negative bacteria such as E. Coli and Salmonella . Herein we describe the ability of CsgF to influence the in-vitro aggregation of human islet amyloid polypeptide (hIAPP), an amyloidogenic polypeptide that is unrelated to Curli. In the presence of CsgF no increase in Thioflavin T fluorescence was observed for freshly solubilized hIAPP monitored as a function of time, suggesting that CsgF prevents the aggregation of hIAPP during the period of observation. A variant of CsgF lacking the first 65 residues in the N-terminus of CsgF retained the ability to inhibit the aggregation of hIAPP suggesting that the ability of CsgF to inhibit the aggregation of hIAPP is mediated by the C-terminal half of the protein.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
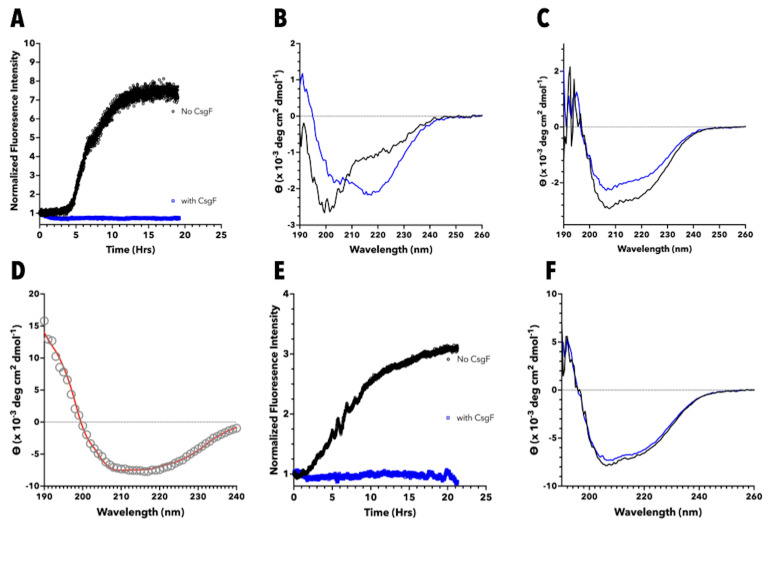 Figure 1
Figure 1- —National Institute of General Medical Sciences (United States)https://ror.org/04q48ey07
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBotanical Research and Applications · Polysaccharides and Plant Cell Walls · Marine and coastal plant biology
Description
Curli, proteinaceous filaments found on the outer surface of bacteria such as E. Coli and Salmonella , are involved in biofilm formation and bacterial attachment to surfaces (Collinson et al., 1997) (Sukupolvi et al., 1997) (Austin et al., 1998) (Römling et al., 1998). Investigation of Curli has shown that these fimbriae possess characteristics similar to amyloid fibers found in a number of diseases such as Alzheimer’s disease, Parkinson’s disease, and type II diabetes. Similar to amyloid, Curli were found to change the spectral properties of Congo Red and Thioflavin T (ThT), as well as contain significant β-sheet character (Chapman et al., 2002). In contrast to amyloid found in disease, which are the result of protein misfolding and are toxic to cells, Curli has a functional role in bacteria. Given these observations Curli have been classified as a functional amyloid: a protein aggregate with amyloid like properties that also serves a functional role. Curli assembly involves six proteins, Csg A, B, C, E, F, and G (Chapman et al., 2002) . CsgA is the major protein component of Curli filaments while CsgB is thought to function as a nucleator of CsgA aggregation as well as to anchor CsgA to the outer surface of the bacterium (Hammar et al., 1996) (Bian & Normark, 1997) . CsgC, CsgE, CsgF, and CsgG provide a variety of important support functions. CsgC and CsgE, periplasmic proteins, prevent the intracellular aggregation of CsgA and/or CsgB (Gibson et al., 2007) (Taylor et al., 2011) (Evans et al., 2015). CsgF, found on the outer surface of bacteria, facilitates the extracellular assembly of CsgA into Curli. In the absence of CsgF, CsgA and CsgB do not localize to the cell surface and are secreted away (Chapman et al., 2002) (Robinson et al., 2006) (Nenninger et al., 2009). CsgG, an outer-membrane channel, secretes the extracellular proteins to the outside milieu (Cao et al., 2014) (Goyal et al., 2014).
The in vitro aggregation of CsgA, the major protein component of Curli, is inhibited by CsgF (Schubeis et al., 2018) and we sought to determine if CsgF is able to inhibit the aggregation of other amyloid forming proteins, and investigated its ability to influence the aggregation of human Islet Amyloid Polypeptide (hIAPP), a 37-residue polypeptide which aggregates to form amyloid deposits found in the pancreas of type II diabetics.
** Salmonella ** ** Typhimurium CsgF Inhibits The Aggregation of hIAPP ** : To determine the influence of CsgF on the aggregation of hIAPP the real time fluorescence intensity of ThT at 482 nm was measured in the absence and presence of S. typhimurium wild type CsgF (CsgF ST-WT ). In the absence of CsgF ST-WT freshly prepared hIAPP exhibited a sigmoidal increase in ThT intensity ( Figure 1A, black symbols). We also monitored the secondary structure of hIAPP immediately after initiating aggregation and at the end of the ThT time course. The circular dichroism (CD) spectrum of freshly dissolved hIAPP exhibited a peak with negative ellipticity at ~ 198 nm suggestive of a predominantly unstructured backbone ( Figure 1B, black line), while the spectrum obtained after the increase in ThT intensity contained a peak with negative ellipticity at ~ 218 nm, suggestive of a predominantly β-sheet backbone structure ( Figure 1B, blue line). These observations are similar to those described in the literature for hIAPP aggregation (Kapurniotu, 2001) (Padrick & Miranker, 2002) (Jayasinghe & Langen, 2004) . Incubating hIAPP with CsgF ST-WT at a 1:1 mole ratio completely abolished the increase in ThT intensity ( Figure 1A, blue symbols). We confirmed the impact of CsgF ST-WT on the aggregation of hIAPP using five independent measurements (see extended data). For each experiment a single solution of hIAPP was separated into two cuvettes and monitored simultaneously with ThT in the absence and presence of CsgF ST-WT . This allowed us to be more confident that any impact we observed with CsgF was due to the protein itself and not because the hIAPP preparation did not aggregate. We also monitored structural changes using CD spectroscopy. Unlike in the case of hIAPP alone no significant change was observed between the CD spectrum obtained immediately after mixing CsgF ST-WT with hIAPP and the spectrum obtained at the end of the ThT time course ( Figure 1C ). Although it is difficult to attribute structural changes to a specific polypeptide contained in a mixture, we expect any conversion of hIAPP from a predominantly random coil to a 𝛽-sheet structure to manifest as a reduction in the negative ellipticity at ~ 200 nm and an increase in negative ellipticity at ~ 218 nm. The ratio of intensities at 208 and 222 nm are 1.28 for the CD spectra obtained immediately after mixing hIAPP with CsgF ST-WT and at the end of the observation period, suggesting the absence of any significant secondary structure change during the period of observation. Taken as a whole these data suggest that CsgF is able to inhibit the aggregation of hIAPP.
** *The N-terminal residues of * CsgF ST-WT
- are not needed to inhibit the aggregation of hIAPP* ** : CsgF ST-WT is a predominantly unstructured protein (Green et al., 2016). The solution structure of E. Coli CsgF indicates that the entire N-terminal half of the protein is unstructured while the C-terminal half of the protein contains an 𝛼-helix followed by a 𝛽-sheet comprised of four antiparallel 𝛽-strands (Schubeis et al., 2018). It has been speculated that the N-terminus may contain an export signal (Schubeis et al., 2018), and the structure of the Curli secretion channel CsgG in complex with CsgF suggests that the N-terminus of CsgF interacts with CsgG via 27 N-terminal residues (Zhang et al., 2020). We analyzed the CD spectrum of a variant of CsgF ST-WT lacking the first 66 residues of CsgF (CsgF Δ66 ) using DichroWeb (Whitmore & Wallace, 2004, 2008) and found a secondary structure distribution of approximately 15 % α-helix, 30% β-strand, 20% turn, with 35% of the protein being unstructured ( Figure 1D ). This distribution compares favorably with the secondary structure distribution calculated fron the solution NMR structure of E. Coli CsgF (approximately 21% 𝛼-helix, 38% 𝛽-strand, 11% turn, and 29% unstructured), suggesting that removal of the N-terminal unstructured region does not alter the overall secondary structure distribution of the remaining amino acid sequence. To determine if the N-terminal region of CsgF plays a role in the protein’s ability to inhibit the aggregation of hIAPP, aggregation was monitored in the presence of CsgF Δ66 . Freshly rehydrated hIAPP incubated in the presence of CsgF Δ66 ( Figure 1E, blue symbols) did not exhibit any increase in ThT intensity at 482 nm. We repeated these experiments with five independent preparations of hIAPP and in each case observed no increase in ThT intensity when hIAPP was incubated with CsgF _Δ66 _ (see extended data). We also monitored changes in secondary structure. Again, no significant change was observed between the CD spectrum obtained immediately after mixing CsgF Δ66 with hIAPP and the spectrum obtained at the end of the ThT timecourse ( Figure 1F ), which is consistent with a lack of aggregation of hIAPP. Thus, it appears that CsgFD66 is still able to inhibit the aggregation of hIAPP, suggesting that the N-terminal unstructured region of CsgF is not crucial to this activity.
Although intriguing, the ability of a Curli accessory protein to inhibit the aggregation of an amyloidogenic protein unrelated to Curli formation is not unique. ThT aggregation studies have shown that CsgC is able to prevent the in vitro aggregation of human α-synuclein, an amyloidogenic protein implicated in Parkinson’s disease (Evans et al., 2015). The ability of CsgC to prevent protein aggregation appears to be specific since it was unable to prevent the aggregation of the Alzheimer’s Aβ 42 peptide even at a 1:1 (CsgC:Aβ 42 ) molar ratio. Based on a comparison of the primary structure of CsgA and α-synuclein, it has been suggested that these two proteins interact with CsgC via a common interaction motif (Evans et al., 2015) . In contrast, there is no discernible region of sequence similarity between CsgA and hIAPP, suggesting that the ability of CsgF ST-WT to inhibit the aggregation of hIAPP does not involve the recognition of a specific amino acid sequence.
Conclusion: The observation that CsgF _ST-WT _ can influence the aggregation of hIAPP adds to the growing body of evidence indicating that the Curli accessory proteins CsgC, CsgE, and CsgF can modulate the aggregation of proteins that form amyloid. Future studies aimed at more clearly identifying the regions and residues of CsgF involved in interacting with hIAPP could help our understanding of the mechanism by which CsgF inhibits protein aggregation.
Methods
** Expression and purification of C-terminal hexahistidine tagged Salmonella Typhimurium CsgF: **
- E. Coli* BL21 (DE3) cells transformed with the appropriate pET21 vector containing the sequence for Salmonella Typhimurium CsgF (wild type or lacking the first 66 residues), fused to the plasmid encoded C-terminal hexahistidine tag, were grown, at 37 ^0^ C to a OD (595 nm) of between 0.5 - 1. Protein production was induced with the addition of 1 mM IPTG, and the cells harvested by centrifugation after 16 hours of incubation at 26 ^0^ C. Cells were resuspended in binding buffer (20 mM phosphate, 20 mM imidazole, 500 mM NaCl ) and B-PER, frozen using liquid nitrogen and lysed using manual grinding with a mortar and pestle. Unbroken cells, and cell debris, were removed by centrifugation (5000 x g) for 20 min at 4 ^0^ C, and histidine tagged protein was recovered from the lysate using a HiTrap TALON Crude Co ^2+^ column (GE Healthcare, Piscataway, NJ) as described by the manufacturer. Resin bound protein was eluted from the column using elution buffer (20 mM phosphate, 250 mM imidazole, 500 mM NaCl) and subsequently desalted and buffer exchanged (in to 20 mM phosphate buffer) using a HiPrep 26/10 Desalting column (GE Healthcare, Piscataway, NJ). The purity of affinity purified histidine tagged CsgF was investigated using SDS PAGE and the presence of the histidine tag was confirmed by western blot using an Anti-His (C-Term)-HRP antibody (LifeTechnologies, Carlsbad, CA).
** Preparation of hIAPP : ** Lyophilized human IAPP peptide was dissolved in Hexafluoro Isopropanol (HFIP) to obtain clear solutions. Peptide concentrations were calculated and aliquots of peptide in HFIP were pipetted into 1.5 mL Eppendorf tubes, mixed with 500 μL of deionized distilled water, immediately frozen in liquid nitrogen, and lyophilized overnight. Dry lyophilized peptide was stored in a vacuum desiccator until use.
** Preparation of hIAPP samples to monitor aggregation: ** Dry lyophilized peptide prepared as described above was dissolved in 20 mM buffer at pH 7.4. The solution was separated into two samples of equal volume. To one CsgF or CsgFD66 was added from a stock solution while an equal volume of buffer was added to the other. The final concentration of hIAPP in each sample was 12.5 or 25 μM.
** ThT fluorescence assay for protein aggregation: ** Aggregation of hIAPP was monitored using the fluorescence intensity increase of ThT. To each aggregation reaction (prepared as described above), a sufficient amount of ThT to yield 25 μM (from 5 mM stock solution in deionized distilled water) was added immediately after dissolving hIAPP in buffer, and real-time emission intensities were measured at 482 nm with excitation at 450 nm. Measurements were performed at room temperature with excitation and emission slit widths of 1 and 10 nm, respectively. Fluorescence measurements were taken using a FluoroMax-3 Spectro-fluorometer (Horiba Jobin Yvon Inc, Edison, NJ).
** *CD Spectroscopy: * ** CD spectra were obtained using a Jaco 810 spectropolarimeter (Jasco Inc., Easton, MD). Measurements were taken every 0.5 nm at a scan rate of 50 nm/min with an averaging time of 1 s. All spectra were collected between 190 and 260 nm using a 2 mm path length quartz cuvette. Spectra were corrected by subtracting an appropriate background, and are presented with intensity units of molar ellipticity.
Reagents
pET21 vectors containing the inserted sequences for CsgF fused to the plasmid-encoded C-terminal hexahistidine tag were obtained from Genscript (Piscataway, NJ). E. Coli BL21(DE) expression competent cells, Bacterial Protein Extraction Reagent (B-PER), and a C-terminal AntiHis antibody were obtained from ThermoFischer Scientific (Grand Island, NY). Hexafluoroisopropanol (HFIP) and ThT were obtained from Sigma-Aldrich (Milwaukee, WI). Synthetic wild-type human Islet Amyloid Polypeptide (h-IAPP) was obtained from Bachem (King of Prussia, PA) or from PolyPeptide Laboratories (Torrence, CA).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Austin John W Sanders Greg Kay William W Collinson S.Karen 199851 Thin aggregative fimbriae enhance Salmonella enteritidis biofilm formation FEMS Microbiology Letters 16220378-109729530110.1111/j.1574-6968.1998.tb 13012.x 9627964 · doi ↗ · pubmed ↗
- 2Bian Zhao Normark Staffan 1997101 Nucleator function of Csg B for the assembly of adhesive surface organelles in Escherichia coli The EMBO Journal 16190261-41895827583610.1093/emboj/16.19.58279312041 PMC 1170214 · doi ↗ · pubmed ↗
- 3Cao Baohua Zhao Yan Kou Yongjun Ni Dongchun Zhang Xuejun Cai Huang Yihua 2014121 Structure of the nonameric bacterial amyloid secretion channel Proceedings of the National Academy of Sciences 111500027-842410.1073/pnas.1411942111 PMC 427332625453093 · doi ↗ · pubmed ↗
- 4Chapman Matthew R. Robinson Lloyd S. Pinkner Jerome S. Roth Robyn Heuser John Hammar Mårten Normark Staffan Hultgren Scott J. 200221 Role of Escherichia coli Curli Operons in Directing Amyloid Fiber Formation Science 29555560036-807585185510.1126/science.106748411823641 PMC 2838482 · doi ↗ · pubmed ↗
- 5Collinson S. Karen Clouthier Sharon C. Doran James L. Banser Pamela A. Kay William W. 1997 Characterization of the Agf BA Fimbrial Operon Encoding Thin Aggregative Fimbriae of Salmonella Enteritidis Advances in Experimental Medicine and Biology 0065-259824724810.1007/978-1-4899-1828-4_379192021 · doi ↗ · pubmed ↗
- 6Evans Margery L. Chorell Erik Taylor Jonathan D. Åden Jörgen Götheson Anna Li Fei Koch Marion Sefer Lea Matthews Steve J. Wittung-Stafshede Pernilla Almqvist Fredrik Chapman Matthew R. 201521 The Bacterial Curli System Possesses a Potent and Selective Inhibitor of Amyloid Formation Molecular Cell 5731097-276544545510.1016/j.molcel.2014.12.02525620560 PMC 4320674 · doi ↗ · pubmed ↗
- 7Gibson D. L White A. P Rajotte C. M Kay W. W 200741 Agf C and Agf E facilitate extracellular thin aggregative fimbriae synthesis in Salmonella Enteritidis Microbiology 15341350-08721131114010.1099/mic.0.2006/000935-017379722 · doi ↗ · pubmed ↗
- 8Goyal Parveen Krasteva Petya V. Van Gerven Nani Gubellini Francesca Van den Broeck Imke Troupiotis-Tsaïlaki Anastassia Jonckheere Wim Péhau-Arnaudet Gérard Pinkner Jerome S. Chapman Matthew R. Hultgren Scott J. Howorka Stefan Fronzes Rémi Remaut Han 2014914 Structural and mechanistic insights into the bacterial amyloid secretion channel Csg G Nature 51675300028-083625025310.1038/nature 1376825219853 PMC 4268158 · doi ↗ · pubmed ↗