Phage endolysins—a novel class of antibacterial agents with a wide range of applications
Evgeny A. Idelevich, Karsten Becker

TL;DR
Phage endolysins are new antibacterial enzymes that can be used in medicine and food safety, but more research and regulatory approval are needed.
Contribution
Phage endolysins are presented as a novel class of antibacterial agents with diverse applications.
Findings
Endolysins can lyse bacteria by targeting their cell walls and are effective against gram-positive bacteria.
Genetic modifications help endolysins overcome the outer membrane of gram-negative bacteria.
Endolysins are being explored for use in human and veterinary medicine, food safety, and biotechnology.
Abstract
Endolysine stellen als „Enzybiotika“ eine neue Klasse antibakterieller Wirkstoffe dar, die natürlicherweise am Ende des lytischen Zyklus in den Bakteriophagen-infizierten Bakterienzellen produziert werden, um den gebildeten Phagenpartikeln vom Zellinneren heraus die Freisetzung aus der Wirtszelle zu ermöglichen. Ihre enzymatische Wirkung auf das Zellwandpeptidoglykan, die zur Lyse der befallenen Bakterien führt, können sie als applizierte Substanz auch von außen entfalten. Während die Endolysinaktivität bei grampositiven Bakterien direkt wirksam werden kann, muss bei gramnegativen Bakterien das Endolysin so modifiziert werden, dass es die äußere Zellmembran überwinden kann. Deshalb sowie zur Optimierung ihrer Spezifität und Stabilität werden Endolysine zunehmend gentechnisch modifiziert und rekombinant produziert, was aufgrund ihres modularen Aufbaus aus lytischen und Bindedomänen…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
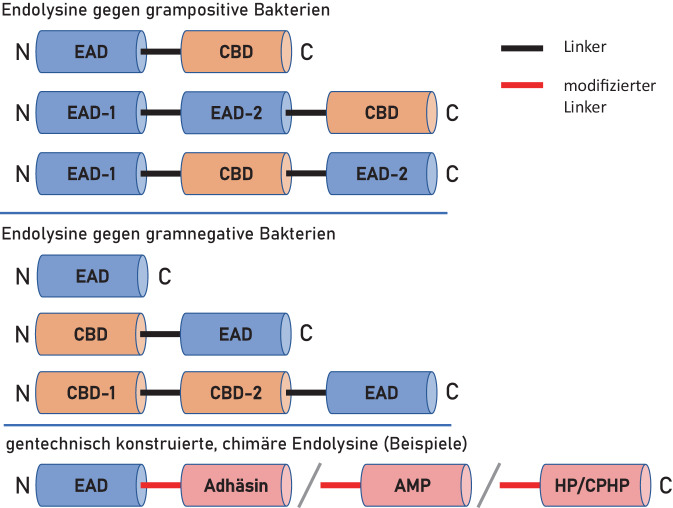 Figure 1
Figure 1- —Universitätsmedizin Greifswald (8976)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Enterobacteriaceae and Cronobacter Research · Probiotics and Fermented Foods
Einleitung
Antimikrobielle Substanzen mit enzymatischer Aktivität (sog. Enzybiotika [1]) stellen neben der Phagentherapie eine weitere Alternative zu den klassischen Antibiotika dar [2]. Neben Enzybiotika mit bakterieller Herkunft, wie z. B. das bakteriolytisch wirkende Lysostaphin von Staphylococcus simulans, haben in den letzten Jahren insbesondere Bakteriophagen-assoziierte Enzybiotika an Interesse gewonnen [3]. Zwei Gruppen sind hier zu unterscheiden, Endo- und Exolysine, die beide lytisch auf die bakterielle Zellwand wirken, wobei die Endolysine zum Zelltod führen. Während Endolysine gegen Ende des viralen Replikationszyklus gebildet werden und generalisiert lytisch von innen („lysis from within“) auf die Zellwand mit dem Ziel der Freisetzung der gebildeten viralen Partikel wirken, sind Exolysine (Ektolysine) viriongebunden und attackieren lokalisiert die Zellwand von außen („lysis from without“), um dem Phagen zu ermöglichen, sein genetisches Material in die Bakterienzelle zu injizieren [4]. Sie werden deshalb auch als Virion- oder „tail“-assoziierte bzw. strukturelle Lysine (VALs, „virion-associated lysins“, bzw. VAPGHs, „virion-associated peptidoglycan hydrolases“) bezeichnet (Details zu Wirkung, Einsatz und Studienlage, siehe [4]). Jedoch können auch Endolysine ihre enzymatische und damit lytische Wirkung auf die bakterielle Zellwand von außen entfalten, wenn sie entsprechend appliziert werden.
Ein erster Nachweis eines lytischen Faktors aus Phagenfiltraten gelang W. R. Maxted im Jahr 1957 [5], gefolgt von der Aufreinigung des C1-Lysins aus dem „Evans B563 (C1)-Phagen“ durch R. Krause [6]. Der Begriff „Endolysin“, der 1958 von Jacob und Fuerst für eine bakteriolytische Substanz vom λ‑Phagen geprägt wurde [7], wird heute für Enzyme verwendet, die die glykosidischen bzw. Peptidbindungen lösen, die dem Peptidoglykan der bakteriellen Zellwand die Stabilität verleihen [8]. Vor allem in älterer Literatur wurden Endolysine auch als Lysozyme (z. B. T7-Lysozym) bezeichnet. Erste therapeutische Anwendungen erfolgten nach der Jahrtausendwende insbesondere gegen grampositive Erreger, da bei diesen die Endolysinaktivität direkt wirksam werden kann. Bei gramnegativen Bakterien muss das Endolysin so modifiziert werden, dass es erst die für diesen Zellaufbau typische äußere Membran überwinden kann. Zu diesem Zweck sowie u. a. zur Optimierung der Endolysinspezifität und Verbesserung ihrer Stabilität werden Endolysine zunehmend gentechnisch modifiziert und rekombinant produziert. Ihre auch nach arzneimittel- bzw. medizinproduktrechtlichen Erfordernissen relativ einfache Herstellbarkeit und Modulierbarkeit führten rasch zu einem breiten Interesse an therapeutischen Einsatzmöglichkeiten in der Humanmedizin, die im Artikel vorrangig betrachtet werden sollen. Darüber hinaus sind vielversprechende Anwendungen in der Veterinärmedizin, Lebensmittelproduktion, Diagnostik und Biotechnologie bereits bekannt.
Im Gegensatz zur Therapie mit natürlichen bzw. gentechnisch veränderten Phagen lassen Endolysine eine Reihe von Vorteilen erwarten, wie eine universellere Anwendbarkeit über Stammgrenzen hinweg, unwahrscheinlichere Resistenzentwicklung, einfachere Modifizierbarkeit, unkompliziertere Formulierung, simplere Pharmakokinetik (Tab. 1) und nicht zuletzt weniger komplizierte regulatorische Aspekte (siehe Beitrag von T. Faltus und T. Buss et al. in diesem Themenheft). In der Literatur ist in den letzten Jahrzehnten eine Vielzahl von Endolysinen unterschiedlichster Phagenherkunft und gentechnischer Modifikation beschrieben worden [9–11], jedoch nur sehr wenige haben das Stadium der klinischen Erprobung erreicht, auf die sich der vorliegende Beitrag fokussiert.Tab. 1Aspekte zum therapeutischen Einsatz von Phagen-Endolysinen, intakten Phagen und Antibiotika im VergleichAspektPhagen-EndolysineBakteriophagenAntibiotikaBeschreibungNatürlich vorkommende Enzyme der Bakteriophagen; gentechnisch modifizierbarNatürlich vorkommende infektiöse Agenzien (Bakterien-befallende Viren), gentechnisch modifizierbarNatürliche, semisynthetische oder synthetische Wirkstoffe mit antibakterieller WirkungWirkspektrumSehr schmal (zumeist stamm- bzw. speziesspezifisch); gentechnisch zuschneidbarSehr schmal Phagotyp-spezifisch (häufig stamm- bzw. speziesspezifisch)Breit bis sehr breit (je nach Wirkungsweise)Einfluss auf die MikrobiotaSehr geringSehr geringHoch (je nach Wirkspektrum)WirkprinzipBakterizid (bakteriolytisch)Bakterizid (bakteriolytisch)Bakteriostatisch oder bakterizidWirkung auf BiofilmeMöglich; gentechnisch modifizierbarMöglich; gentechnisch modifizierbarSubstanzabhängigResistenzentwicklungUnwahrscheinlichSchnell (innerhalb einer Applikation möglich)Langsam bis schnell (substanzabhängig)Horizontaler ResistenzgentransferNicht möglichNicht beschriebenHäufig (über mobile genetische Elemente)Vertikaler ResistenzgentransferNicht möglichRegelmäßigRegelmäßigAntibiotika-KreuzresistenzKeineKeineEinzel‑, Multi- bis Panresistenz (je nach Resistenzmechanismus)WirkungskinetikSehr schnell (Minuten-Stunden)Langsam (nach Etablierung des lytischen Zyklus; ggf. erst nach einigen Zyklen)Schnell (Stunden; abhängig von bakterizider bzw. bakteriostatischer Wirkung)PharmakokinetikKontrollierte Dosierung mit kalkulierbarer Bioverfügbarkeit, Aktivität, Metabolisierung und AusscheidungKontrollierte Dosierung, weiterer Verlauf (Selbstreplikation) abhängig vom Vorhandensein von Target-Organismen und deren Quantität und VerteilungKontrollierte Dosierung mit kalkulierbarer Bioverfügbarkeit, Aktivität, Metabolisierung und AusscheidungUnerwünschte WirkungenUnwahrscheinlich bei topischer Applikation; bei systemischer Applikation potenzielle ImmunogenitätRelativ gering (bei lytischen Phagen), ansonsten potenzielle Gefahr durch von Prophagen kodierte Toxingene und Wirtszell-BeimengungenGering bis hoch (je nach Substanzeigenschaften)GMP-Herstellung^a^Unproblematisch biotechnologischKaum umgesetzt (aufwendig)Unproblematisch chemisch bzw. biotechnologischNotwendigkeit der ProduktaktualisierungUnwahrscheinlichHäufig aufgrund von Resistenzentwicklung und Erreger(stamm)wechselKeine (allerdings Unwirksamkeit durch Resistenzentwicklung)Regulative Aspekte und ZulassungInternational etabliert (Arzneimittel oder Medizinprodukt)In einigen Ländern/Regionen etabliert (auch als individueller Heilversuch), aktuelle Aufnahme in das Europäische Arzneibuch (Pharmacopoeia Europaea)International etabliert (Arzneimittel)^a^GMP Good Manufacturing Practice (Gute Herstellungspraxis)
Struktur und Wirkmechanismus
Natürliche Endolysine
Endolysine degradieren überwiegend als Hydrolasen („phage-derived peptidoglycan hydrolases“ – PGH) die bakterielle Zellwand, deren Hauptkomponente Peptidoglykan (Murein) in Form eines dicken 3‑dimensionalen Netzes ist; hinzukommen bei grampositiven Bakterien „sekundäre Zellwandpolymere“ („secondary cell wall polymers“ – SCWPs). Weiterhin können auch lytische Phagen-Transglykosidasen das Peptidoglykan durch nichthydrolytische Mechanismen strukturell schwächen. Die Peptidoglykanstruktur ist innerhalb einer Gattung in hohem Maße konserviert, die genaue molekulare Beschaffenheit des Peptidoglykans und der SCWPs kann jedoch bis hinunter auf Spezies- und Stammebene spezifisch sein [12]. Somit entstand im Laufe der Bakterien-Phagen-Koevolution im Gegenzug eine recht große Vielfalt an Endolysinen, die sich in ihrer Struktur, katalytischen Aktivität, Enzymkinetik und insbesondere in ihrer Spezifität unterscheiden [13]. Endolysine von Phagen, die Bakterien mit grampositivem Zellwandaufbau (einschließlich Mykobakterien) befallen, unterscheiden sich strukturell von denjenigen, deren Phagen gramnegative Bakterien infizieren (Abb. 1; [8]).Abb. 1Schematische Darstellung natürlicher und chimärer Endolysine von Bakteriophagen mit Spezifität gegen grampositive oder gramnegative Bakterien bestehend jeweils aus enzymatisch/katalytisch aktiven Domänen („enzymatically active domain“ – EAD) und Zellwand-bindenden Domänen („cell wall-binding domain“ – CBD) bzw. weiteren, gentechnisch eingefügten Bestandteilen, die durch molekulare Linker verbunden sind. Chimäre Endolysine können u. a. mit Adhäsinen, antimikrobiellen Peptiden (AMP) oder „homing peptides“ (HP) bzw. „cell-penetrating homing peptides“ (CPHP) fusioniert sein
Wie anhand der Aufklärung der Kristallstruktur bestätigt werden konnte, sind Endolysine modular aufgebaut [14]. Die Endolysine von Phagen mit grampositiver Wirtsspezifität besitzen zumeist ein Molekulargewicht von 15–40 kDa. Sie bestehen aus einer oder zwei enzymatisch aktiven Domänen („enzymatically active domains“ – EADs; auch „catalytic domain“ – CD), die N‑terminal bzw. zusätzlich zentral angeordnet sind, sowie einer C‑terminalen Zellwand-bindenden Domäne („cell wall-binding domain“ – CBD). Die Domänen sind durch einen kurzen, flexiblen Linker verbunden. Die EAD können Aktivitäten verschiedener Enzymklassen (Glykosidasen, Amidasen und Endopeptidasen) aufweisen und somit unterschiedliche Bindungen der Peptidoglykanstruktur der bakteriellen Zellwand attackieren. Beispielsweise handelt es sich bei der N‑terminalen EAD von Staphylokokken-Phagen in den meisten Fällen um eine cystein- und histidinabhängige Amidohydrolase/Peptidase (CHAP), die als D‑Ala-Gly-Endopeptidase die Bindung zwischen D‑Ala des Peptidstrangs und der Pentaglycinbrücke des Peptidoglykans spaltet. Bei Endolysinen mit mehr als zwei Domänen können auch Molekulargewichte von > 50 kDa erreicht werden. Sowohl die Architektur des Peptidoglykans anderer grampositiver Erreger als auch der Endolysine der diese Arten befallenden Phagen weisen hierzu Unterschiede auf. Die bis vor kurzem bekannten Endolysine der gramnegative Bakterien befallenden Phagen sind leichter (15–20 kDa), globulär strukturiert und sind mit einer alleinigen EAD simpler aufgebaut. Neue Studien zeigen, dass bei diesen Phagen auch eine modulare Endolysinkonfiguration vorkommen kann, die ein oder zwei CBDs am N‑Terminus aufweisen, während sich das EAD-Modul am C‑Terminus befindet, also eine im Vergleich zu Phagen mit grampositiver Spezifität inverse Ausrichtung der molekularen Struktur [15].
Die CBDs sind für die nichtkovalente Bindung an verschiedene Liganden der bakteriellen Zellwandoberflächen, wie z. B. Peptidoglykanbestandteile, Lipopolysaccharide, Teichonsäuren, Peptide oder Kohlenhydrate, verantwortlich und vermitteln die enzymatische Aktivität der EAD [16, 17]. Ihre Bindung ist wirtsspezifisch, von speziesübergreifend bis spezies- und stammspezifisch [13]. Die auf Basis umfangreicher Phagenbibliotheken früher übliche Methodik zur Stammtypisierung von Staphylokokken, Listerien und anderen Bakterien mittels Phagentypisierung (Phagotyping; [18]) beruht hingegen auf der spezifischen Bindung der Phagenrezeptorbindeproteine, z. B. der Phagenschwanzfasern, an bakterielle Wirtsoberflächenrezeptoren.
Bei den meisten Phagen werden für die natürliche Endolysinwirkung von innen zusätzlich noch kleine hydrophobe Proteine, sog. Holine, benötigt, um zuerst die Zytoplasmamembran perforieren zu können, bevor die Zellwand für die Endolysine erreichbar wird. Für die gramnegativen Bakterien wird noch eine andere Gruppe von Proteinen, die sog. Spanine, erforderlich, um auch die äußere Membran penetrieren zu können [19]. Diese stellt für natürliche Endolysine, wenn sie von außen wirken sollen, also z. B. im therapeutischen Einsatz, eine lyseverhindernde Barriere bei den gramnegativen Bakterien dar.
Gentechnisch modifizierte Endolysine
Mit der Aufklärung der Endolysinstruktur und der Funktion der einzelnen Strukturkomponenten [14] sowie den gentechnischen Möglichkeiten zur Modifizierung der Struktur der natürlichen Endolysine eröffnete sich eine Reihe von Möglichkeiten, um Limitationen für den Einsatz von Endolysinen insbesondere im medizinischen Bereich zu überwinden. Hierzu zählen in erster Linie eine Ausdehnung der oft zu schmalen Target-Spezifität und die Anwendungsausweitung auf gramnegative Bakterien sowie Verbesserungen der Endolysinstabilität und -pharmakokinetik. Anstrengungen in dieser Richtung führten zur gentechnischen Konstruktion chimärer Endolysinstrukturen (Chimäre von altgriech. Χίμαιρα – Mischwesen der griechischen Mythologie; [20]), die auch als „Chimeolysine“ („chimeolysins“; [21]) bezeichnet werden. Diese sind dadurch charakterisiert, dass adhäsions- und lysevermittelnde Bestandteile verschiedener Speziesherkunft (z. B. Phagen- und bakterielle Anteile) eingesetzt werden, die durch einen Linker verbunden sind. Weitere Funktionalisierungen der Endolysine, wie die Fusion mit zellpenetrierenden Peptiden zur Eliminierung intrazellulärer Bakterien oder mit „homing peptides“ als zielerkennende kurze Peptide zur gerichteten Anreicherung in bestimmten Geweben (auch in Kombination als „cell-penetrating homing peptides“ – CPHP), erweitern die zukünftigen therapeutischen und/oder diagnostischen Einsatzmöglichkeiten von Endolysinen in vielfältigster Weise [22].
Für chimäre Endolysine ist bereits eine Vielzahl von Kombinationen mit unterschiedlichster Spezifität beschrieben worden. Eine der ersten Anwendungen basierte auf der synergistischen Wirkung von Lysostaphin und LysK [23]. Lysostaphin wirkt als Zink-Endopeptidase auf die Polyglycinvernetzungen in der Peptidoglykanschicht der Staphylokokkenzellwand. Es handelt sich um ein Bakteriocin, das von S. simulans gebildet wird und als sogenanntes Staphylococcin mit hoher Aktivität Staphylococcus aureus eliminiert, während die Wirkung auf koagulasenegative Staphylokokken deutlich geringer ausfällt. Beim LysK handelt es sich um ein hochwirksames Endolysin [24] vom Bakteriophagen K aus der Myoviridae-Familie, einem Phagen mit vergleichsweise breiter Wirtsaktivität gegen koagulasepositive und -negative Staphylokokken, wobei die CHAP-Domäne für die Wirkung essenziell ist [25]. Das rekombinante PRF-119 (Hyglos GmbH, Bernried, Deutschland) und dessen stabilitätsverbesserte Weiterentwicklung HY-133 (Hyglos, später HYPharm GmbH, Bernried, Deutschland) wurden als erste Lysine nach diesem Prinzip als Chimäre aus der CHAP-Domäne vom Bakteriophagen K (CHAP_K_) und der CBD von Lysostaphin generiert und umfangreich auf deren antibakterielle Aktivität getestet [26, 27]. PRF-119/HY-133 zeigten sich hoch aktiv gegenüber mehr als 1000 S.-aureus-Isolaten, einschließlich Hospital‑, Community- und Livestock-associated MRSA (Methicillin-resistenter S. aureus) der verschiedensten klonalen Linien, weiteren Resistenzphänotypen, einschließlich Mupirocinresistenz und Small-Colony-Variant-(SCV-)Phänotypen, die intrazellulär persistierende Varianten darstellen [26–28]. Untersuchungen zur Bakterizidie ergaben Werte der minimalen bakteriziden Konzentrationen (MBK) in der Größenordnung der minimalen Hemmkonzentrationen (MHK; MHK_50/90_ und MBK_50/90_ jeweils im Bereich von 0,12–0,5 mg/L) bei sehr schnellem, konzentrationsabhängigem Wirkungseintritt innerhalb von wenigen Minuten bis ca. 2 h [29].
Das Prinzip der Ergänzung von CHAP_K_ oder anderer katalytischer CHAP-Domänen (z. B. LysRODI und LysC1C) um die Lysostaphin-CBD oder andere Domänen (z. B. M23-Endopeptidase oder SH3b) wurde ebenfalls von anderen Arbeitsgruppen u. a. zur Aktivitäts- und Spezifitätsverbesserung für Staphylokokken-wirksame Endolysine eingesetzt [30–33]. Auch gegen andere bakterielle Pathogene wurde bereits eine Reihe chimärer Lysine konstruiert. Mit ClyR konnte ein Chimeolysin mit sehr guter Aktivität und insbesondere einem erweiterten Wirtsspektrum gegen die meisten Streptokokkenarten (u. a. S. pyogenes, S. agalactiae, S. dysgalactiae, S. mutans, S. pneumoniae) sowie einige Enterokokken- und Staphylokokkenarten konstruiert werden [34]. Obwohl bisher keine Gardnerella-spezifischen Bakteriophagen bekannt sind, konnten DNA-Regionen in den Genomen identifiziert werden, die auf einen Prophagenursprung hindeuten [35]. Endolysin-typische EAD- und CBD-Sequenzen wurden als rekombinante Proteine exprimiert und die Kombination mit der höchsten Bakterizidie (PM-477) wurde weiter untersucht und zeigte eine hohe Aktivität gegen G. vaginalis, selbst in Biofilmen [36].
Eine Möglichkeit zur Überwindung der Gramnegativenproblematik ist die Fusion eines Endolysins mit Molekülen, die in der Lage sind, die äußere Membran zu permeabilisieren. Ein Beispiel ist das gentechnisch konstruierte Hybridtoxin aus der FyuA-(„ferric-yersiniabactin-uptake“-)Transporter-Bindungsdomäne von Pesticin und dem N‑Terminus vom T4-Lysozym [37]. Das von Yersinia pestis produzierte Pesticin wird unter Stressbedingungen als Bakterizidin produziert, um verwandte Stämme zu eliminieren. Erst die Wechselwirkung zwischen dem Outer-Membrane-Protein FyuA und Pesticin ermöglicht den Import von Pesticin durch die äußere Membran in das Periplasma. Dort entfaltet es seine bakterizide Wirkung, indem es das Peptidoglykan durch seine Muramidase-(Lysozym‑)Aktivität abbaut. Pesticin-exprimierende Stämme schützen sich allerdings selbst, indem sie ein Immunitätsprotein („pesticin immunity protein“ – Pim) exprimieren, das die Aktivität des Toxins hemmt. Das Hybridtoxin konnte spezifisch Yersinia- und pathogene Escherichia-coli-Stämme lysieren und außerdem die Pim-Wirkung umgehen, was ihm einen deutlichen Vorteil gegenüber dem alleinigen Pesticineinsatz verschafft [37]. Die Aktivität konnte durch einen verlängerten Linker zwischen dem Peptid und dem Endolysin oder durch eine Kombination von polykationischen und hydrophoben/amphipathischen Peptiden weiter gesteigert werden [38]. Für derartige Fusionsproteine aus Endolysinen und antimikrobiellen Peptiden mit membranpenetrierenden Eigenschaften wurde die Bezeichnung „Artilysine“ eingeführt [38].
Interessant sind Fusionen mit Cecropinen, die als antimikrobielle Peptide zum innaten Immunsystem von Insekten gehören und durch ihre amphipathische Natur die Permeabilisierung der äußeren Membran erleichtern. Das Artilysin eLysMK34 als Fusionsprodukt aus dem Acinetobacter-baumannii-Phagen „MK34-Lysin“ und Cecropin A zeigte u. a. eine verbesserte Abtötungskinetik im Vergleich zum Wildtyplysin [39]. Weitere Möglichkeiten zur Überwindung der äußeren Membran sind Endolysinfusionen mit rezeptorbindenden Phagenproteinen (Innolysine; [40]) und Pyocindomänen (Lysocine; [41]).
Einsatzmöglichkeiten und deren Wertung
Ihre relativ einfache gentechnische Modifizierbarkeit macht Endolysine zu aussichtsreichen Kandidaten für die unterschiedlichsten Anwendungen nicht nur zu therapeutischen, kosmetischen und diagnostischen Zwecken in der Human- und Veterinärmedizin, sondern auch in der Lebensmittelindustrie für Biokonservierung und Lebensmittelsicherheit, in der Landwirtschaft und Umwelt zur biologischen Schädlingsbekämpfung und Bioremediation sowie zu vielfältigen Zwecken in der Biotechnologie.
Präventive und therapeutische Anwendungen
Die im Gegensatz zu den klassischen Antibiotika quasi phylogenetisch ableitbare und damit klar definierbare Spezifität auf Genus‑, Spezies- oder Stammebene ertüchtigt Endolysine zu idealen Wirkstoffen im Sinne des Konzepts einer ultimativ schmalen, nur auf das kausale Agens ausgerichteten Therapie bakterieller Infektionen, im Idealfall ohne jegliche Beeinträchtigung der Mikrobiota des Wirtes. Dieses Prinzip einer möglichst weitgehenden Vermeidung von Antibiotikaresistenzselektionsdrücken ist von noch größerer Bedeutung im präventiven Bereich, z. B. zur Vermeidung postoperativer Wundinfektionen. Auch aus dieser Sicht sind Endolysine vielversprechende Präparate, wie u. a. zur Eradizierung einer nasalen MRSA/S.-aureus-Kolonisation als Quelle nachfolgender nosokomialer Infektionen [42].
Perfekt ergänzend zur bakteriziden Wirkung ist die Fähigkeit einiger Endolysine, mit bakteriellen Biofilmen von grampositiven und gramnegativen Erregern zu interagieren und diese abzubauen [43]. Es konnte z. B. für Biofilme von Klebsiella pneumoniae und Acinetobacter baumannii gezeigt werden, dass enzymatische Endolysinaktivität mit Biofilm-protektiven Verbindungen wie extrazellulärer DNA und polyanionischen Kohlenhydraten interagiert und Biofilm-zerstörende Effekte auslöst [44]. Für detaillierte Auflistungen der Vielzahl bereits beschriebener Endolysine und anderer Phagenproteine für einen potenziellen präventiven und/oder therapeutischen Einsatz, die sich noch im experimentellen oder frühen klinischen Stadium befinden, sei hier auf aktuelle Übersichtsarbeiten verwiesen [45–50].
Da der lytische Wirkmechanismus von Endolysinen unabhängig von bakteriellen Resistenzmechanismen gegen Antibiotika ist, sind Kreuzresistenzen mit Antibiotika ohne Relevanz. Das Potenzial von Resistenzentwicklungen gegen eingesetzte Endolysine ist gleichfalls niedrig. Hinzu kommt die ultraschnelle bakterizide Wirkung von Endolysinen innerhalb weniger Minuten bis Stunden, die den Zeitraum für einen hypothetischen Selektionsdruck extrem kurz hält [29]. Der schnelle Wirkungseintritt ist von besonderem Interesse für präventive Zwecke, da z. B. präoperative Wartezeiten bis zu einer erfolgreichen MRE-Dekolonisierung von derzeit mehreren Tagen drastisch reduziert werden könnten. Topisch eingesetzt lässt sich ihre Aktivität ohne systemische Wirkung streng lokal begrenzen.
Phänotypische Varianten mit reduziertem Metabolismus, oft in Kombination mit intrazellulärer Persistenz oder Biofilmeinbettung, sind eine der Hauptursachen für eine funktionelle Resistenz gegenüber in vitro als empfindlich getesteten Antibiotika und können die Ursache für chronisch persistierende und rezidivierende Infektionsverläufe bilden. Derartige Persister‑, Dormanz- oder Small-Colony-Variant-(SCV-)Stadien können durch Phagenendolysine erfolgreich lysiert werden [28, 51]. Im Gegensatz zum Einsatz von Phagenendolysinen ist für die Therapie mit kompletten Phagen bekannt, dass diese die Entstehung von Persisterzellen sogar induzieren können [52, 53].
Weitere vorteilhafte Faktoren umfassen die geringe Wahrscheinlichkeit von Nebenwirkungen sowohl bei topischer als auch parenteraler Applikation aufgrund ihrer sehr geringen Toxizität [54, 55]. Translations- und anwendungsrelevante Vorteile wie die hohe Vielfalt natürlich vorkommender Endolysine und deren unkomplizierte gentechnische Modifizierbarkeit, die einfache Produktion, einschließlich unter Bedingungen der Guten Herstellungspraxis (Good Manufacturing Practice – GMP), ihre gute Stabilität (ggf. nach gentechnischer Optimierung), die vielfältigen Formulierungsmöglichkeiten (z. B. als Lösungen, Salben oder Sprays) runden das Bild einer potenziell erfolgreichen, neuen Antiinfektivasubstanzklasse ab.
Bei Proteinbiopharmazeutika besteht grundsätzlich die Möglichkeit einer unerwünschten Auslösung von Immunreaktionen, die bei parenteraler Applikation über die Generierung neutralisierender Antikörper unter Umständen zu einem Verlust der Wirksamkeit des Medikaments sowie zur Autoimmunität und zu Überempfindlichkeitsreaktionen führen können. In einer Studie zur möglichen Endolysininaktivierung durch Antikörper konnte gezeigt werden, dass LysGH15-spezifische Antikörper die Abtötungseffizienz des Endolysins gegen MRSA weder in vitro noch im Mausexperiment in vivo beeinträchtigten [56].
Endolysine in klinischer Erprobung
Im Gegensatz zu der großen Zahl bekannter und charakterisierter Endolysinmoleküle befinden sich nur sehr wenige davon in der klinischen Entwicklung bzw. wurden oder werden in klinischen Studien an Menschen untersucht. Eine Übersicht von bisher stattgefundenen und laufenden kontrollierten klinischen Studien mit Endolysinen ist in der Tab. 2 zusammengefasst. Bisher wurden nur gegen S. aureus gerichtete Endolysine in klinischen Studien untersucht, wobei zwischen zwei Indikationen bzw. Verabreichungswegen unterschieden werden kann. Während einige Substanzen für eine topische Anwendung entwickelt werden, zielen die anderen auf eine intravenöse Verabreichung ab. Interessant könnte auch eine Kombinationstherapie von Endolysinen und klassischen Antibiotika sein [57].Tab. 2Übersicht über randomisierte kontrollierte klinische Studien (RCT) mit EndolysinenPrüfpräparatStudiendesignZielorganismusVerwendungszweckVerabreichungswegRegistrierung^a^LiteraturAP-SA02RCT, verblindet, Phase I/IIS. aureusBakteriämieIntravenösNCT04160468n. a.Exebacase (CF-301)RCT, verblindet, Phase IIIS. aureusEndokarditis, BakteriämieIntravenösNCT04160468Fowler, Jr. et al. (2024; [58])Exebacase (CF-301)RCT, verblindet, Phase IIS. aureusEndokarditis, BakteriämieIntravenösNCT03163446Fowler, Jr. et al. (2020; [57])Exebacase (CF-301)RCT, verblindet, Phase IS. aureusEndokarditis, BakteriämieIntravenösNCT02439359n. a.HY-133RCT, verblindet, Phase I/IIS. aureusEradikation der nasalen KolonisationIntranasalNCT06290557n. a.LSVT-1701 (SAL200)RCT, verblindet, Phase IS. aureusBakteriämieIntravenösNCT03446053Wire et al. (2022; [73])N‑Rephasin (SAL200)RCT, verblindet, Phase IIS. aureusBakteriämieIntravenösNCT03089697n. a.N‑Rephasin (SAL200)RCT, verblindet, Phase IS. aureusBakteriämieIntravenösNCT01855048Jun et al. (2017; [74])P128RCT, verblindet, Phase I/IIS. aureusNasale EradikationIntranasalNCT01746654n. a.Staphefekt (SA100)RCT, verblindetS. aureusAtopische DermatitisHautcremeNCT02840955Totté et al. (2017; [65]),de Wit et al. (2019; [75])RCT randomisierte kontrollierte Studie („randomized controlled trial“), n. a. nicht verfügbar („not available“)^a^National Clinical Trials (NCT) Identifier Number, Identifikationsnummer der klinischen Studie in der Datenbank „ClinicalTrials.gov“ der U.S. National Library of Medicine
Unter den topischen Endolysinen in der klinischen Entwicklung ist das rekombinant hergestellte, chimäre HY-133 zu erwähnen, welches gerade in einer deutschen klinischen Phase-I/II-Studie insbesondere auf seine Sicherheit und Verträglichkeit, aber auch auf Wirksamkeit zur nasalen Eradikation von S. aureus untersucht wird. Diese doppelblinde randomisierte kontrollierte Studie (NCT06290557) befindet sich aktuell (Stand März 2025) in der aktiven Phase der Probandenrekrutierung. Neben einer einzelnen Verabreichung des Wirkstoffs sollen nach einer Interimssicherheitsbewertung multiple Applikationen sowie die Verabreichung der erhöhten Dosierungen des Wirkstoffes erfolgen.
Unter den Endolysinen für eine systemische Anwendung hat der Wirkstoff Exebacase (CF-301), der zur intravenösen Therapie von der durch S. aureus verursachten Endokarditis bzw. Bakteriämie entwickelt wird, die meisten klinischen Studien absolviert. Nach einer Phase-I-Studie (NCT02439359), in der die Sicherheit von Exebacase an gesunden Probanden gezeigt wurde, folgten die US-amerikanischen Studien der Phase II und Phase III. Die Ergebnisse der Phase-II-Studie im Design einer randomisierten, Placebo-kontrollierten Überlegenheitsstudie waren vielversprechend. Die klinische Ansprechrate betrug am Tag 14 in der Gruppe, die Exebacase zusammen mit einem Standardantibiotikum erhielt, 70,4 %, während die Ansprechrate in der Gruppe, die nur Antibiotika appliziert bekam, bei 60,0 % lag (p = 0,31). Signifikant war der Effekt von Exebacase in der MRSA-Subgruppe (74,1 % vs. 31,3 %, p = 0,01; [57]). Allerdings konnte in der Phase-III-Studie (DISRUPT), die gleichfalls als randomisierte Überlegenheitsstudie angelegt worden war, kein positiver Effekt der zusätzlichen Gabe von Exebacase bestätigt werden [58]. Die klinischen Ansprechraten am Tag 14 betrugen in der MRSA-Population 50,0 % bei kombinierter Gabe von Exebacase plus Antibiotikum und 60,6 % bei alleiniger Verabreichung von Antibiotika (p = 0,392). Angesichts der vielversprechenden Ergebnisse der Phase-II-Studie war dieses Ergebnis unerwartet und die Studie musste nach der Zwischenbewertung des Datenüberwachungskomitees vorzeitig abgebrochen werden [58]. Von den Autoren werden eine Heterogenität innerhalb der Studienpopulation und ein relativ geringer Stichprobenumfang diskutiert. Weitere Endolysine befinden sich zurzeit in der Entwicklungs-Pipeline für unterschiedliche klinische Indikationen (Tab. 2).
Zu erwähnen sind neben den in Tab. 2 aufgeführten kontrollierten Endolysinstudien eine nichtkontrollierte klinische Studie an Patienten mit atopischer Dermatitis, die mit Staphefekt™ SA.100 behandelt wurden [59], sowie Fallberichte zur Behandlung von Hauterkrankungen, die mit S. aureus assoziiert werden [60], und Knieprotheseninfektionen durch S. epidermidis [61].
Marktverfügbar sind bereits Medizinprodukte als rezeptfrei erhältliche Hautcremes oder Gele mit gentechnisch modifizierten Endolysinen als Inhaltsstoff, die als Mikrobiom-freundlich/-unterstützend beworben werden. Das betrifft Staphefekt™ SA.100 (auch als Endobioma™ vermarktet), das zur topischen Behandlung oder deren Unterstützung bei atopischem Ekzem/Neurodermatitis und Rosacea angeboten wird. Die Studienlage zu diesen Produkten ist sehr begrenzt und die Zulassung von Phagenendolysinen als Medizinprodukt anstatt als Arzneimittel ist zu hinterfragen.
Weitere Endolysinanwendungen
Neben einem therapeutischen bzw. präventiven Einsatz der Endolysine in der Medizin ist eine Reihe weiterer Einsatzgebiete bereits beschrieben. Ein vielversprechender Bereich ist das sog. Mikrobiom-Engineering. Obwohl die Bedeutung des Mikrobioms für die menschliche Gesundheit zunehmend erkannt und berücksichtigt wird, fehlt es bisher an einer breiten Palette maßgeschneiderter Werkzeuge, um gezielt die Zusammensetzung der Mikrobiota zu modulieren, insbesondere wenn es gilt, einzelne Stämme oder Spezies ohne „Kollateralschaden“ für andere Mikroorganismen zu eradizieren. Gerade hier können Endolysine im Gegensatz zu den Antibiotika ihre Vorteile bezüglich Spezifität und relativ einfacher gentechnischer Modifizierbarkeit zum Tragen bringen und machen sich so zu aussichtsreichen Kandidaten für diesen Zweck [62]. Erste Ergebnisse aus In-vitro‑, Ex-vivo- und In-vivo-Studien untermauern mögliche Einsatzgebiete, wie u. a. die Therapie einer Infektion mit Clostridioides (Clostridium) difficile im Rahmen einer Antibiotika-assoziierten Durchfallerkrankung zur Remodulierung der Darmmikrobiota [63], die präoperative Eradizierung einer S.-aureus/MRSA-Besiedlung der Nasenhöhle bzw. von geschädigter Haut [64–66] oder die spezifische Beseitigung von Gardnerella vaginalis aus der Vaginalmikrobiota bei bakterieller Vaginose [36].
Die gleichen Vorteile einer hohen Spezifität und gentechnischen Modifizierbarkeit lassen Endolysine auch als ideale Biosonden zum Einsatz in der mikrobiologischen Diagnostik erwarten. Neben CBDs von Endolysinen stellen rezeptorbindende Phagenproteine („tail fibers“ und „tailspikes“) einen Schlüssel für die Entwicklung von diagnostischen Werkzeugen u. a. für die Markierung, die Immobilisierung und den Nachweis definierter bakterieller Targets dar [40]. Hierzu können – als Ersatz von diagnostisch eingesetzten Antikörpern – fluoreszenz- oder enzymmarkierte CBDs bzw. rezeptorbindende Phagenproteine z. B. zur immunomagnetischen Separation von Pathogenen eingesetzt werden. Hauptsächlich für den Einsatz in der Lebensmittelproduktion ist bereits eine Reihe von Testsystemen mit Wildtyp- oder modifizierten Endolysinen beschrieben, u. a. zum Nachweis von S. aureus [67], Listerien [68], Salmonella Typhimurium [69] und Clostridium perfringens [70].
In der Lebensmitteltechnologie werden Endolysine bereits erfolgreich als Mittel zur unterstützenden Biokonservierung von Lebensmitteln eingesetzt [71]. Insbesondere thermophile Endolysine (> 65 °C) können hier in Zukunft breite Verwendung finden [72]. Für Anwendungsmöglichkeiten im One-Health-Bereich sei auf die Literatur verwiesen [72].
Fazit
Die Identifizierung von neuen Phagenendolysinen und deren Charakterisierung mit dem Ziel eines möglichen Einsatzes in der Human- und Veterinärmedizin zu diagnostischen, präventiven und therapeutischen Zwecken sowie in vielen anderen Bereichen, wie der Lebensmitteltechnologie oder unter One-Health-Gesichtspunkten, stellen ein rasant wachsendes Forschungsfeld dar. Ihre natürliche Wirkungsbeschränkung auf definierte Taxa, ihre ultraschnelle Wirksamkeit und ihre relativ einfache gentechnische Modifizierbarkeit machen ihren Einsatz als zukünftige, maßgeschneiderte Produkte für die hochspezifische, Kollateralschäden-vermeidende Bekämpfung bakterieller Infektionen vielversprechend. Klinische Studien müssen jedoch noch zeigen, dass der in anderen Bereichen (z. B. Diagnostik und Lebensmitteltechnologie) bereits gelungene Einsatz rekombinanter Endolysine auch als Arzneimittel oder Medizinprodukt für die Human- und Veterinärmedizin umsetzbar ist.