Comprehensive genome-wide identification and expression analysis of the EPF/EPFL gene family in oat
Qingxue Jiang, Lin Ma, Siyuan Guo, Xinyue Zhou, Zhipeng Zhang, Yanhong Cui, Jun Tang, Dengxia Yi, Xuemin Wang

TL;DR
This study identifies and analyzes EPF/EPFL genes in oats, revealing their roles in stress responses and tissue-specific expression patterns.
Contribution
The first genome-wide identification and expression analysis of the EPF/EPFL gene family in oat (Avena sativa).
Findings
33 AsEPF/EPFL genes were identified, with 28 containing the EPF domain and 5 the stomagen domain.
Expression patterns show tissue-specificity and stress-induced expression under salt and drought conditions.
Phylogenetic analysis grouped genes into five clades with monocot and dicot homologues, indicating early evolutionary divergence.
Abstract
Epidermal pattern factor-like (EPF/EPFL) genes are a unique class of small, secreted peptides found in plants that play crucial roles in plant stress responses. A genome-wide analysis revealed 33 AsEPF/EPFL genes in oats (Avena sativa), with 28 containing the conserved EPF domain and 5 harbouring the stomagen domain. These proteins share 2–6 conserved motifs, reflecting functional modularity. The phylogenetic classification grouped these genes into five evolutionarily conserved clades containing both monocot and dicot homologues, indicating early divergence prior to monocot–dicot speciation. Expression profiling revealed distinct tissue-specific patterns: preferential expression in roots (12 genes), stems (6 genes), leaves (5 genes), and spikes (7 genes), with 3 genes showing dual peak expression in stems and leaves. Further analysis of gene expression under salt and drought stress…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
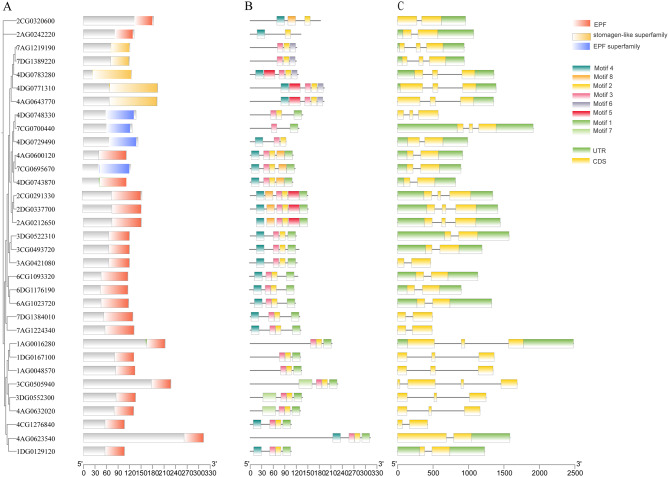 Figure 1
Figure 1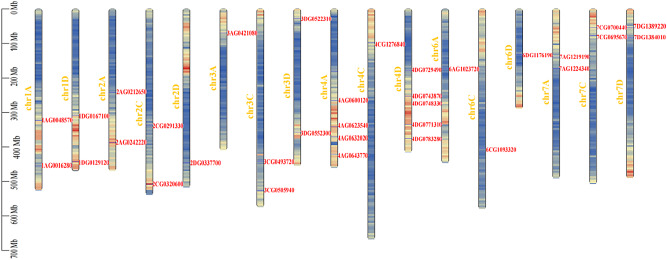 Figure 2
Figure 2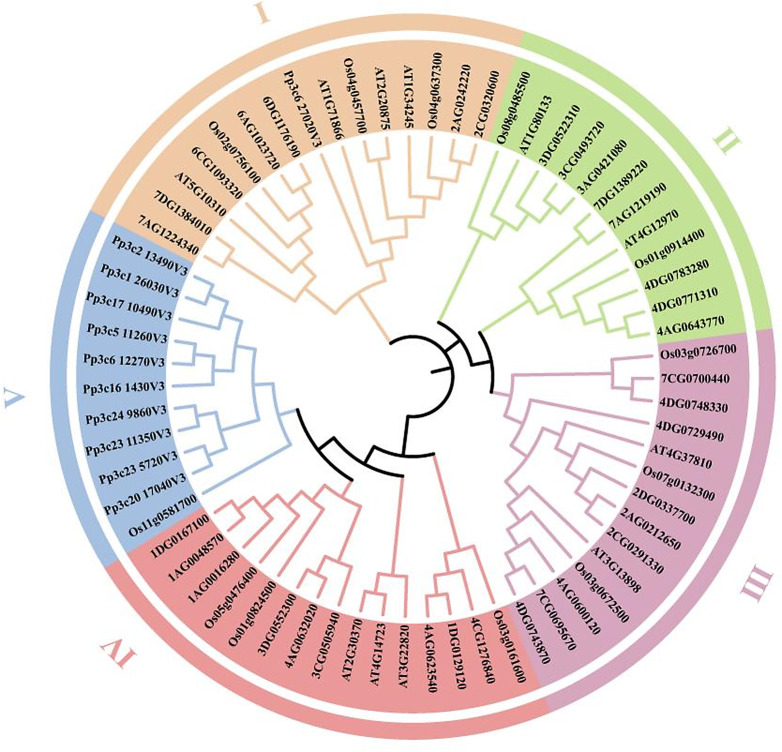 Figure 3
Figure 3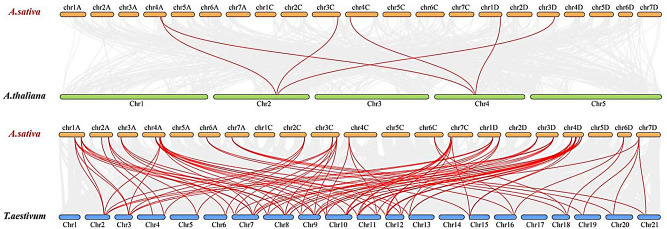 Figure 4
Figure 4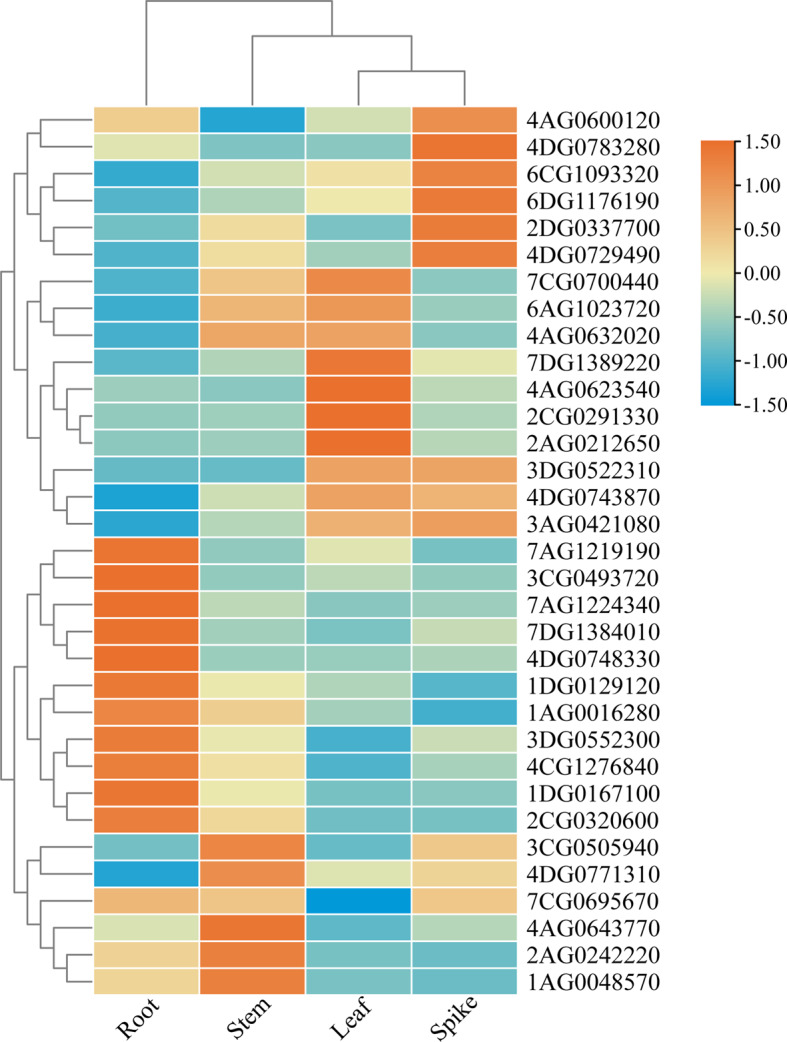 Figure 5
Figure 5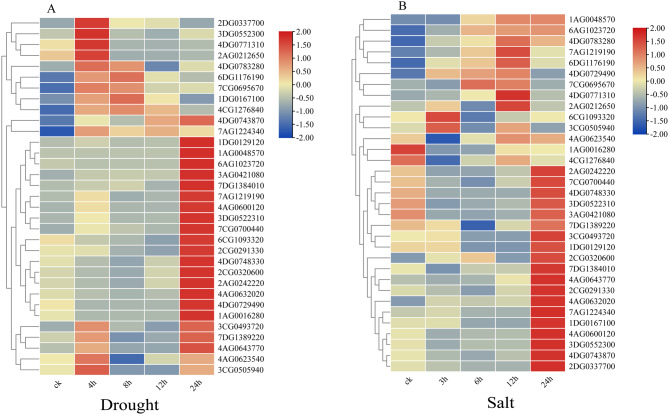 Figure 6
Figure 6- —The earmarked fund for CARS,China
- —The National Crop Germplasm Resources Center,China
- —https://doi.org/10.13039/501100012421Agricultural Science and Technology Innovation Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Reproductive Biology · Plant Molecular Biology Research · Plant biochemistry and biosynthesis
Background
Plants are immobile, which means they must endure abiotic stresses such as drought, salinity, and extreme temperatures. These stressors significantly limit their distribution and impact their growth and development [1]. Most of the water absorbed by plants is lost to the atmosphere through stomata, and regulating transpiration through stomatal control is an effective strategy to increase plant drought resistance [2]. Stomata play key roles in regulating carbon dioxide uptake and water loss [3]. In recent years, small peptides encoded by the epidermal patterning factor (EPF/EPFL) gene family have been identified as key factors in plant responses to abiotic stress [4]. Naoki et al. revealed, through modelling, that EPF/EPFL peptides share a common three-dimensional structure composed of a framework and a loop [5]. These peptides interact with signalling pathways associated with stomatal development to regulate stomatal density and opening/closing, thereby increasing plant adaptability under stress conditions.
Genetic and physiological analyses have shown that EPF/EPFL genes have great potential for improving plant WUE and drought tolerance by regulating stomatal development [6]. Recent studies have shown that regulating stomatal development through molecular methods can significantly increase plant drought resistance. Compared with the wild type strain, the Dunn team reported a dual effect after the TaEPF1B gene was overexpressed in wheat: the stomatal density decreased by 39%, whereas the water use efficiency increased by 27% [7]. Research on poplar has revealed a more complex regulatory mechanism. In the 84 K variety, the overexpression of PdEPFL6 not only reduced the stomatal density by 42% but also significantly increased leaf water retention by balancing the transpiration rate (decreased by 58%) and photosynthetic efficiency (maintained at 85% of normal levels) [8]. In potato, overexpression of the StEPF2 gene led to unique phenotypic characteristics. Under drought stress, the stomatal density decreased by 35%, but the optimization of stomatal opening and closing rhythms successfully maintained the leaf water content at more than 90% of normal levels [9]. Notably, heterologous expression of the poplar PdEPF3 gene in Arabidopsis revealed the feasibility of cross-species regulation. Transgenic plants presented a 22% increase in the photosynthetic rate and a 41% reduction in the transpiration rate under drought conditions, indicating excellent drought resistance phenotypes [10]. Li et al. found that hydrogen sulfide (H₂S) can modify EPF2 and EPFL9 through direct cysteinylation, affecting their protein activities and thereby influencing stomatal development in Arabidopsis, increasing the stomatal density in the lower epidermis of leaves [11]. These results indicate that the EPF/EPFL gene family plays an important role in regulating stomatal density, thereby affecting plant drought tolerance and water use efficiency. In addition, the EPF/EPFL gene family has been shown to participate in the plant inflorescence structure, leaf morphology, pollen tube elongation and other morphogenesis processes [12]. Sun et al. performed a qRT‒PCR analysis, which revealed that the expression of the TaEPFL1 gene in the stamens (PS) of the wheat male sterile mutant hts-1 (exhibiting homeotic transformation of anthers into pistils) was significantly upregulated. Specifically, TaEPFL1 levels in PS stamens were approximately 20-fold higher than those in normal pistils (P) and 15-fold higher than those in normal stamens (S). These results suggest that TaEPFL1 overexpression may contribute to the homeotic transformation of stamens into pistils in wheat [13].
Given the crucial role of EPF/EPFL gene family members in regulating plant stress responses, members of the EPF/EPFL gene family have been systematically identified in various plants. HARA et al. first identified 11 EPF/EPFL gene family members in Arabidopsis thaliana, namely, AtEPF1, AtEPF2, AtEPFL1, AtEPFL2, AtEPFL3, AtEPFL4, AtEPFL5, AtEPFL6, AtEPFL7, AtEPFL8, and AtEPFL9 [14]. EPF/EPFL gene family members have subsequently been identified in different plants, such as tomato (Solanum lycopersicum, 14 genes) [15], rye (Secale cereale, 12 genes) [16], barley (Hordeum vulgare, 11 genes) [17], rice (Oryza sativa, 12 genes) [18], maize (Zea mays, 17 genes) [18], and wheat (Triticum aestivum, 35 genes) [19]. While the complete oat genome was not fully sequenced and published until 2022 due to its large size (10.56 Gb) and complex hexaploid structure, the comprehensive genome-wide identification of EPF/EPFL family genes in Avena sativa remains to be completed [20].
Oat is an annual herbaceous plant of the Poaceae family that is widely grown as a dual-purpose crop for grain and fodder, particularly in cool temperate regions [21]. Oats possess excellent characteristics, such as tolerance to poor soils, salinity, cold, and drought; high forage yields; and high nutritional contents, making them valuable in livestock production [22]. Nevertheless, under the agricultural context of ensuring food security and maintaining a red line of 1.8 billion mu (120 million hectares) of arable land in China, oats are often cultivated in marginal lands prone to drought and salinity. Existing varieties struggle to meet the industrial demands. The EPF/EPFL gene family plays an important role in plant adaptation to adverse stress and growth and development processes. Therefore, identifying and exploring drought- and salt tolerance-related EPF/EPFL genes in oats is crucial for utilizing biotechnological approaches to breed new oat germplasms, supporting the development of drought-resistant and salt-tolerant oat varieties.
This study employed bioinformatics methods to systematically identify EPF/EPFL gene family members in the oat genome. The physicochemical properties, chromosomal localization, conserved domains, and gene structures of these family members were analysed. Furthermore, a clustering analysis was performed to elucidate the phylogenetic relationships of these family members in plants. Additionally, qRT‒PCR was employed to investigate the tissue-specific expression patterns of AsEPF/EPFL gene family members and their responses to drought and salt stress. These findings lay a foundation for a deeper understanding of the biological functions of the EPF/EPFL gene family in oats and provide important candidate genes for the discovery of key genes related to drought and salt tolerance in oats.
Materials and methods
Plant materials and methods
Seeds of the Avena sativa L. cultivar ‘Mengyan 1’ were selected for this study. Seeds were germinated in Petri dishes, transferred to seedling pots, and cultivated in a greenhouse with the relative humidity maintained between 65% and 75%. When the seedlings reached the three-leaf stage, they were subjected to drought and salt stress treatments. Drought stress was simulated using 20% PEG 6000, and samples were collected at 0 h, 4 h, 8 h, 12 h, and 24 h after treatment. Salt stress was induced by adding 150 mM NaCl, and samples were collected at 0 h, 3 h, 6 h, 12 h, and 24 h after treatment. Additionally, tissues (roots, stems, leaves, and spikes) from oats at the heading stage were sampled. All the samples were immediately frozen in liquid nitrogen and stored at -80 °C for subsequent RNA extraction. Each sample had three biological replicates.
Identification and analysis of the EPF/EPFL gene family
The protein sequences of 11 AtEPF/EPFL genes were downloaded from the Arabidopsis genome database TAIR (https://www.arabidopsis.org/). Using TBtools v1.121 software, BLASTP searches were performed against the oat genome to identify AsEPF/EPFL proteins (E value < 1e^− 5^). Whole-genome data, CDSs, protein sequences, and annotation files for oats were downloaded from the Ensembl Plants database (https://plants.ensembl.org/). The Hmm search tool in TBtools v1.121 was used to further search for and identify oat EPF/EPFL proteins. The protein sequences obtained via both methods were merged, redundant sequences were removed, and the final set of oat EPF/EPFL gene family members was obtained. The Pfam (http://pfam.xfam.org/) and NCBI-CDD (https://www.ncbi.nlm.nih.gov/cdd) websites were used for additional protein identification. TBtools v1.121 software was used to predict physicochemical properties such as the molecular weight and isoelectric point of the AsEPF/EPFL protein sequences.
Conserved motif and gene structure analyses
TBtools v1.121 was used to analyse the gene structures of AsEPF/EPFL family members. MEME online software (http://meme.nbcr.net/meme) was used to analyse conserved motifs in AsEPF/EPFL proteins, with the maximum motif count set to 7. The results were visualized using TBtools v1.121.
Chromosomal localization analysis of the EPF/EPFL gene family
Chromosome density files were extracted using TBtools v1.121 Chromosome Heatmap Data. The locations of AsEPF/EPFL family genes on chromosomes were determined using the oat genome database, and their distribution on chromosomes was visualized using TBtools v1.121 Gene Location Visualize from GTF/GFF.
Evolutionary analysis of the AsEPF/EPFL system
The protein sequences of 56 EPF/EPFL genes from oats, rice, Arabidopsis and Physcomitrella patens were aligned using MUSCLE in MEGA-X software. A neighbour-joining (NJ) phylogenetic tree was constructed using MEGA-X, and the resulting tree was visualized and beautified using the iTOL website (https://itol.embl.de).
Synteny and homologous gene pairs
The collinearity of EPF/EPFL genes in A. thaliana, T. aestivum, and A. sativa was analysed using the One Step MCScanX module in TBtools v1.121. The Dual Synteny Plot plugin in TBtools was employed to visualize the collinearity of the AsEPF/EPFL genes. This visualization revealed the chromosomal distribution of AsEPF/EPFL genes, their collinear relationships, and their correspondence with EPF/EPFL genes in other species.
Expression pattern of the EPF/EPFL genes in oat
Total RNA was extracted from different tissues and from samples subjected to drought and salt stress treatments using the Fast Pure Plant Total RNA Isolation Kit (TIANGEN, Beijing, China). cDNA was synthesized using the Trans Script II One-Step gDNA Removal and cDNA Synthesis Super Mix Kit (Vazyme, Nanjing, China). qRT‒PCR was performed using Hieff^®^ qPCR SYBR Green Master Mix (Yeasen, Shanghai, China) in a 20 µL reaction volume, which included 10.0 µL of SYBR Green Master Mix, 0.4 µL each of forwards and reverse primers, 8.2 µL of sterile water, and 1 µL of cDNA template. The PCR program included initial denaturation at 95 °C for 3 min, followed by 40 cycles of 95 °C for 15 s, 60 °C for 30 s, and 72 °C for 20 s. A melting curve analysis was performed from 60 °C to 95 °C. AsActin was used as the reference gene, and relative expression levels were calculated using the 2^−ΔΔCt^ method. All the experiments were performed with three biological replicates. The sequences of the primers used for qRT‒PCR are listed in Table 1.
This methodology section outlines the detailed procedures used for the identification, characterization, and expression analysis of the AsEPF/EPFL gene family in oats under various conditions.
Data analysis
The qRT‒PCR data were processed using Excel 2019. GraphPad Prism software was used to generate heatmaps depicting the relative expression levels of the 33 AsEPF/EPFL genes across different tissues and under various abiotic stress conditions. Additionally, a clustering analysis was performed to analyse the expression patterns.
Table 1qRT‒PCR primers for the AsEPF/EPFL genesPrimer nameForward primer (5′-3′)Reverse primer (5′-3′)4AG0623540CGCATGATGGAGGAAGGGAAGAGTGTTGTTGTCAAGGGAATTTC3DG0522310CTCTTCTTCCTGTCGCATCTGCCCTCCTCGTCTCATCCAG7AG1219190GCTGCCGTTGTTGTGCTATCTCGCTCCGCTCCGTCATCGC4AG0600120GCGGAAGAAGAAGAAGGAGGAGACGGCTTGTAGTTGGAGTGG4DG0743870CGCCGAGGAGAAGAGCATTGTAGTTGGAGTGGTCGTCGTAG3AG0421080CTCTTCTTCCTGTCGCACCTGATCCTCGGCACGCACCAG6CG1093320CCTGTCCACTTTTCAGAACCTATCCACCCGAGCGGCTTGTAG7DG1384010GACTAGCAGCCGCACTACTTGCGCACCTTCTCCTCCTCTCC3DG0552300GTGCTGACGGTGGTGGTGCAAGGTGCTGGTGTTCGCC2DG0337700TCTTCCTCTCCTTGCTCCTCTTCTCCTCCTCCTGCTCCTTATTGG3CG0493720GCTTGGCTCTTCTTCCTATCGCATCCTCGGCACGCACCAGG7AG1224340GACTAGCAGCCGCACTACTTCGCACCTTCTCCTCCTCTC1DG0129120CGCTCCTCCTCCTCCTCTTCAGCCTGTCGCCGCACTTG1DG0167100CGTCGTCGCCTCCCTCTGCGCTGCTCCTGACATCTTCC6DG1176190CCCTCTCCACTTTTCAGAACCCGAGCGGCTTGTAGTTGG4DG0783280TGCTCCTCTTCTTCCTCCTCTCGGGTGGTTTCTCCTCGTTCAG4DG0729490GCTTCTCCTCTTCTCCTTCCTGTCCTCCTCCTCGGCAATGG1AG0016280CCTTCCATCCATCCATCCACAGGACGACGAGCAGCCTTAGC2CG0320600CGCAACACCTCATAGACATAGCGAACACAGCCACGAGAAGAAG7CG0695670CTGCGGAAGAAGAAGAAGAAGAAGTGGTCGTCGTAGGAAGAAGG4CG1276840CGCTCATCCTCCTCTTCCTCCAGCCTGTCGCCGCACTTG2AG0242220GGAGGCGGCGGAGATGAGGGCGATGGAGCACTTGAAGC4DG0748330AACTCCTGCTACTTCACTCCTCATCACCACCGCCGACATC7CG0700440CTGCTCCTCCTTCTGCTACCATCATCACCGCCGACACC1AG0048570GCGTCGTCGCCTCCCTCTGGCTGCTCCTGACATCTTCC7DG1389220CTGCCGTTGTTGTGCTATCTCGCTCCGCTCCGTCATCGC6AG1023720AGAACCTGGCGGAGGACAAGGAGCGTCGTGGCAACCTG4AG0632020GTGCTGACGGTGGTGGTGGCGCAAGGTGCTGGTGGTC4AG0643770GCTCCTCTTCTTCCTCCTCTCCCTATATGTTCTCCTGTTAATCCTTGG3CG0505940CTACAGTGGCGAGTGAGACCAGCGGCAGCAACAGTATGG2CG0291330TCTTCCTCTCCTTGCTCCTCTTCCTCCTCCTGCTCCTTACTG4DG0771310GCTCCTCTTCTTCCTCCTCTCTATATGTTCTCCTGTTAATCCTTGG2AG0212650CTTCCTCTCCTTGCTCCTCTTCCCTCCTCCTGCTCCTTACTGGAsActinCACTGCCGAGCGGGAAATTGTGATGGAAGGCTGGAAGAGGAC
Results
Identification and analysis of the physicochemical properties of the EPF/EPFL gene family
In the oat genome, a total of 33 nonredundant EPF/EPFL gene family members were identified (Table 2). The analysis of the physicochemical properties analysis the following characteristics of the AsEPF/EPFL proteins: (1) the length of the AsEPF/EPFL proteins ranged from 106 to 213 amino acids; (2) the molecular weights varied between 11,418.25 and 32,984.15 Da, with an average of 15,386.06 Da; (3) the isoelectric points (pI) ranged from 7.09 to 10.82; (4) the instability index of the AsEPF/EPFL proteins ranged from 44.8 to 87.54, indicating their stability; and (5) the aliphatic index ranged from 59.65 to 89.67. The hydrophilicity index ranged from − 0.447 to 0.02, with the average suggesting hydrophilic properties. These results indicate that AsEPF/EPFL proteins are predominantly composed of basic amino acids and exhibit hydrophilic characteristics (Table 2).
Table 2. Physical and chemical properties of AsEPF/EPFLsIDChr LocationAmino AcidsMolecular WeightTheoretical pIInstability IndexAliphatic IndexGrand Average of HydropathicityAVESA.00010b.r1.4AG06235404 A:341,292,372,341,293,95631232984.510.8280.4369.33-0.23AVESA.00010b.r1.3DG05223103D:26,890,21126,891,78311913208.210.0367.0680.25-0.099AVESA.00010b.r1.7AG12191907 A:138,739,243–138,740,18112013231.278.7973.7472.5-0.236AVESA.00010b.r1.4AG06001204 A:265,140,156–265,141,07311112086.139.6962.579.28-0.232AVESA.00010b.r1.4DG07438704D:253,105,873–253,106,68911011928.969.6960.980.91-0.174AVESA.00010b.r1.3AG04210803 A:69,461,343–69,461,80811913111.3710.1564.2877.82-0.206AVESA.00010b.r1.6CG10933206 C:402,160,455–402,161,58611512368.179.4653.3162.17-0.187AVESA.00010b.r1.7DG13840107D:84,111,192–84,111,68112813356.338.661.7479.38-0.091AVESA.00010b.r1.3DG05523003D:357,151,732–357,152,97813515116.6510.0587.5469.93-0.253AVESA.00010b.r1.2DG03377002D:445,938,958–445,940,37015016521.388.7550.8889.670.007AVESA.00010b.r1.7AG12243407 A:173,108,213–173,108,69913113509.498.8560.7480.53-0.062AVESA.00010b.r1.3CG04937203 C:441,104,824–441,106,01411913148.3710.478.2167.98-0.316AVESA.00010b.r1.1DG01291201D:442,739,192–442,740,41910611476.379.7975.3477.55-0.076AVESA.00010b.r1.6DG11761906D:131,330,530–131,331,42611512259.989.6154.0859.65-0.276AVESA.00010b.r1.1DG01671001D:307,877,586–307,878,94713113518.629.2563.968.47-0.087AVESA.00010b.r1.4DG0729494D:177,100,140–177,101,12814115669.049.4667.7878.94-0.377AVESA.00010b.r1.4DG07832804D:375,659,485–375,660,84412413680.557.095570.89-0.346AVESA.00010b.r1.1AG00162801 A:454,487,915–454,490,39521222026.489.7155.9375.47-0.005AVESA.00010b.r1.7CG06956707 C:77,733,004–77,733,89711612753.0910.258.2676.72-0.398AVESA.00010b.r1.2CG03206002 C:508,222,649–508,223,60718219158.448.9358.2884.340.232AVESA.00010b.r1.4CG12768404 C:98,248,152–98,248,57310611418.259.7981.4271.13-0.146AVESA.00010b.r1.2AG02422202 A:387,377,357–387,378,42913113949.378.849.4676.790.007AVESA.00010b.r1.4DG07483304D:268,883,169–268,883,74113614330.348.946.3376.84-0.222AVESA.00010b.r1.7CG07004407 C:54,446,612–54,448,52412613596.649.7244.879.84-0.245AVESA.00010b.r1.1AG00485701 A:322,903,882–322,905,22713413877.989.172.5163.96-0.151AVESA.00010b.r1.7DG13892207D:52,814,691–52,815,63011913200.298.787177.98-0.15AVESA.00010b.r1.6AG10237206 A:172,085,798–172,087,12411712432.229.6154.3860.34-0.218AVESA.00010b.r1.4AG06320204 A:369,998,489–369,999,65113014531.999.7888.5466.62-0.231AVESA.00010b.r1.4AG06437704 A:420,590,561–420,591,91319120834.688.756.6664.97-0.406AVESA.00010b.r1.3CG0505943 C:526,163,238–526,164,92422724469.279.769.6276.48-0.152AVESA.00010b.r1.2CG02913302 C:336,260,669–336,262,01014916356.128.5251.9683.09-0.073AVESA.00010b.r1.4DG07713104D:334,947,335–334,948,72319221118.018.8360.3864.11-0.447AVESA.00010b.r1.2AG02126502 A:242,310,696–242,312,14215016511.348.7555.52890.02
Analysis of the conserved motifs and structures of the EPF/EPFL gene family
The results of the conserved domain analysis are shown in Fig. 1A. The conserved domain of 24 AsEPF/EPFL members is EPF; the conserved domain of 4 AsEPF/EPFL members (4DG0748330, 7CG0700440, 4DG0729490, and 7CG0695670) is stomagen; and the conserved domain of 5 AsEPF/EPFL members (7AG1219190, 7DG1389220, 4DG0783280, 4DG0771310, and 4AG0643770) is the stomagen-like superfamily. Further analysis of the conserved motifs of the AsEPF/EPFL proteins revealed 7 conserved motifs, named Motif 1 to Motif 7 (Fig. 1B). Among these, 17 AsEPF/EPFLs contain 4 motifs; 5 AsEPF/EPFLs contain 3 motifs; 2AG0242220 contains only 2 motifs, namely, Motif 4 and Motif 8; and 7CG0700440 contains only 2 motifs, namely, Motif 3 and Motif 1. In addition, by comparing the genomic sequences of AsEPF/EPFL genes with their corresponding CDSs, the structures of AsEPF/EPFL genes was obtained (Fig. 1C). Most (31) AsEPF/EPFL genes contain 2 exons and 1 intron, whereas 2CG0320600 and 4AG0643770 have only one exon. These results indicate that the AsEPF/EPFL gene family in oats is highly conserved.
Fig. 1A Evolutionary tree and conserved domains of AsEPF/EPFL proteins. B. Conserved motifs of the EPF/EPFL gene family. C. Gene structure of the EPF/EPFL gene family
Chromosomal localization of the EPF/EPFL gene family
The chromosomal localization analysis revealed that the 33 AsEPF/EPFL genes are located on 17 chromosomes of the oat genome (Fig. 2). Further analysis indicated that these genes are unevenly distributed across the 17 chromosomes. Chromosome 4D contains 5 AsEPF/EPFL genes, namely, 4DG0743870, 4DG0783280, 4DG0729490, 4DG0748330, and 4DG0771310; chromosome 4 A contains 4 AsEPF/EPFL genes, namely, 4AG0623540, 4AG0600120, 4AG0632020, and 4AG0643770; chromosomes 1 A, 1D, 2 A, 2 C, 3 C, 3D, 7 A, 7 C, and 7D each contain 2 AsEPF/EPFL genes; and chromosomes 2D, 3 A, 4 C, 6 A, 6 C, and 6D each contain only 1 AsEPF/EPFL gene. These results indicate that no gene duplication occurred during the formation of the AsEPF/EPFL genes in the oat genome.
Fig. 2. Chromosomal locations of AsEPF/EPFL genes.Gene density is depicted in a color gradient, with red indicating regions of higher gene density and blue indicating regions of lower gene density
Phylogenetic analysis of the EPF/EPFL gene family
Multiple sequence alignments were performed on the amino acid sequences of the 33 AsEPF/EPFL proteins from oats, 12 OsEPF/EPFL proteins from rice, 11 PpEPF/EPFL proteins from moss plants (Physcomitrella patens) and 11 AtEPF/EPFL proteins from Arabidopsis, totalling 67 EPF/EPFL proteins, to explore the evolutionary relationships of the EPF/EPFL gene family members in oats and other species. A phylogenetic tree was constructed (Fig. 3). The results of the phylogenetic analysis presented in Fig. 3 show that the EPF/EPFL family can be classified into five distinct subgroups. Subgroups II, III, and IV contain conserved EPF/EPFL members from Arabidopsis (eudicot), rice (Oryza sativa; monocot), and oats (Avena sativa; monocot), with AsEPF/EPFL sequences exhibiting closer phylogenetic relationships to the OsEPF/EPFL sequences within each subgroup. Notably, P. patens is exclusively clustered in subgroups I and V.
Fig. 3. Phylogenetic tree of EPF/EPFL genes.Clades I–IV are color-coded to indicate the evolutionary groupings of the EPF/EPFL proteins. Note: The accession numbers for the OsEPF1-12 genes of the rice EPF/EPFL family are as follows: LOC_Os03g0161600, LOC_Os11g0581700, LOC_Os05g0476400, LOC_Os01g0824500, LOC_Os04g0457700, LOC_Os04g0637300, LOC_Os08g0485500, LOC_Os02g0756100, LOC_Os01g0914400, LOC_Os03g0726700, LOC_Os03g0672500, and LOC_Os07g0132300. The accession numbers for the AtEPF1-2 genes of the Arabidopsis EPF/EPFL family are AT2G2075 and AT1G34245. The accession numbers for the AtEPFL1-9 genes are AT5G10310, AT4G37810, AT3G13898, AT4G14723, AT3G22820, AT2G30370, AT1G71866, AT1G80133, and AT4G12970. The accession numbers for the PpEPF genes are Pp3c24_9860V3, Pp3c23_11350V3, Pp3c23_5720V3, Pp3c20_17040V3, Pp3c17_10490V3, Pp3c5_11260V3, Pp3c16_1430V3, Pp3c1_26030V3, Pp3c2_13490V3, Pp3c6_27020V3, and Pp3c6_12270V3
Collinearity analysis of EPF/EPFLs in oat, arabidopsis, and wheat
We further investigated the evolutionary history of the EPF/EPFL gene family across different species by conducting a comprehensive collinearity analysis of the genomes of A. sativa, A. thaliana, and T. aestivum. The results of the analysis, as shown in Fig. 4, revealed significant gene collinearity among these three species. Specifically, six pairs of collinear genes were identified between oat and A. thaliana: 4AG0632020, 4AG0623540, 3CG0505940, 4CG1276840, 1DG0129120, and 3DG0552300. These genes are unevenly distributed across chromosomes 4 A, 3 C, 4 C, 1D, and 3D in oats and other chromosomes.
In contrast, the collinearity between oat and wheat was much stronger, with 100 pairs of collinear genes identified. This finding suggests a closer evolutionary relationship of the EPF/EPFL gene family between oat and wheat than in A. thaliana. Further examination revealed that the EPF/EPFL genes in the oat genome are widely distributed across 16 of its chromosomes, whereas no EPF/EPFL genes were detected on the other five chromosomes (5 A, 1 C, 5 C, 2D, and 5D).
Fig. 4. Covariance analysis of EPF/EPFL genes from A. sativa, A. thaliana and T.aestivum. The grey lines in the background indicate blocks of collinearity within A. sativa and the indicated plants, whereas the red lines indicate homozygous EPF/EPFL gene pairs
Expression patterns of EPF/EPFL genes in different oat tissues
The expression patterns of 33 AsEPF/EPFL genes in different oat tissues (roots, stems, leaves, and spikes) were analysed using qRT‒PCR. As shown in Fig. 4, the expression levels of the 33 AsEPF/EPFL genes varied significantly across the four tissues. Twelve genes (7AG1219190, 3CG0493720, 7AG1224340, 7DG1384010, 4DG0748330, 1DG0129120, 1AG0016280, 3DG0552300, 4CG1276840, 1DG0167100, 2CG0320600, and 7CG0695670) presented the highest expression levels in roots; six genes (4AG0632020, 3CG0505940, 4DG0771310, 4AG0643770, 2AG0242220, and 1AG0048570) presented the highest expression levels in stems; five genes (7CG0700440, 7DG1389220, 4AG0623540, 2CG0291330, and 2AG0212650) presented the highest expression in leaves; and seven genes (4AG0600120, 4DG0783280, 6CG1093320, 6DG1176190, 4DG0729490, 2DG0337700, and 3AG0421080) presented significantly higher expression levels in spikes than in other tissues. Additionally, 7CG0700440, 6AG1023720, and 4AG0632020 presented significantly higher expression in stems and leaves than in roots and spikes(Fig. 5). The differing expression levels across tissues suggest that the 33 AsEPF/EPFL genes may perform distinct biological functions in various oat tissues.
Fig. 5. Expression patterns of AsEPF/EPFL genes in different oat tissues
Characteristics of EPF/EPFL expression in response to abiotic stress
The expression patterns of 33 AsEPF/EPFL genes under drought stress were analysed using qRT‒PCR. As shown in Fig. 5A, all 33 AsEPF/EPFL genes presented varying degrees of upregulation under drought stress (Fig. 6A). Among them, 9 AsEPF/EPFL genes (2DG0337700, 3DG0552300, 4DG0771310, 2AG0212650, 4DG0783280, 7CG0695670, 7AG1224340, 4AG0623540, and 3CG0505940) peaked at 4 h poststress; 6DG1176190 and 1DG0167100 peaked at 8 h; and 22 AsEPF/EPFL genes (4DG0743870, 1DG0129120, 1AG0048570, 6AG1023720, 3AG0421080, 7DG1384010, 7AG1219190, 4AG0600120, 3DG0522310, 7CG0700440, 6CG1093320, 2CG0291330, 2CG0320600, 2AG0242220, 2CG0320600, 4DG0748330, 4AG0632020, 4DG0729490, 1AG0016280, 3CG0493720, 7DG1389220, and 4AG0643770) peaked at 24 h. These results suggest that AsEPF/EPFL genes may participate in the oat drought stress response through different regulatory pathways.
In response to salt stress, the qRT‒PCR analysis revealed that only 2 AsEPF/EPFL genes (1AG0016280 and 4CG1276840) were downregulated, whereas the remaining 31 AsEPF/EPFL genes were upregulated to varying degrees (Fig. 6B). Among these, 6CG1093320 and 3CG0505940 peaked at 3 h poststress; 7CG0695670 peaked at 6 h; 8 AsEPF/EPFL genes (1AG0048570, 4DG0783280, 7AG1219190, 6DG1176190, 4DG0729490, 4DG0771310, 2AG0212650, and 4AG0623540) peaked at 12 h; and the remaining 20 AsEPF/EPFL genes peaked at 24 h (Fig. 6B). These results indicate significant differences in the response times of different AsEPF/EPFL genes to salt stress.
Fig. 6(A) Relative expression of the AsEPF/EPFL genes under PEG stress. (B) Relative expression of the AsEPF/EPFL genes under NaCl stress.ck: non-stressed control plants harvested at matching time points
Discussion
Plant epidermal patterning factor-like (EPF/EPFL) peptides are small secretory peptides that are widely present in plants and play crucial roles in growth, development, and stress resistance [23]. Previous studies have shown that some signal peptides are involved in signalling during plant growth and development, whereas others participate in defence responses or symbiotic processes [24, 25]. In this study, we utilized bioinformatics tools to identify 33 AsEPF/EPFL genes in the hexaploid oat genome. Research has revealed significant differences in the number of family members across species, ranging from diploid species (A. thaliana with 11 genes, H. vulgare with 11 genes, and Z. mays with 17 genes) to polyploid species (T. aestivum with 35 genes and O. sativa with 33 genes). The relatively high number of AsEPF/EPFL genes in oat may be attributed to its complex polyploid evolutionary history (2n = 6x = 42, AADDCC), with its genome originating from interspecific hybridization between a tetraploid (CCDD) and a diploid (AA) ancestor [26]. The phylogenetic analysis revealed that AsEPF/EPFLs can be divided into five subgroups. Notably, P. patens was found only in branches I and V, indicating that these two branches may have originated after the evolution of pteridophytes. Although lower bryophytes and ferns have not yet developed stomata and vascular tissues, members of the EPF/EPFL gene family have already emerged. The increase in the number of EPF/EPFL gene family members throughout evolution is likely attributable to gene or genome duplication. Overall, the phylogenetic analysis suggested that the EPF/EPFL family originated before the divergence of monocots and dicots, with the biological functions of orthologous genes being conserved across different species.
Although there are few reports on EPF/EPFL genes in oat, the EPF/EPFL gene family has been confirmed in multiple plant species to regulate plant development and growth, especially in morphogenesis processes such as stomata, awns, stamens, and fruit skin formation [27–30]. In this study, we analyzed the tissue-specific expression patterns of 33 AsEPF/EPFL genes in oat and found significant tissue specificity. Most genes were highly expressed in roots and spikes: 12 genes showed the highest expression in roots, while 8 genes exhibited elevated expression in leaves. This expression pattern suggests that AsEPF/EPFL genes may be involved in regulating the development of roots, leaves, and spikes in oat. For example, CmEPF1 is predominantly expressed in melon roots, where it modulates root cortical aerenchyma to regulate growth [31]. After overexpressing AtEPFL9 in Arabidopsis, the overexpressing lines exhibited a higher number of stomata in mature leaves, indicating that AtEPFL9 positively regulates stomatal development in plants [32]. Similarly, overexpression of OsEPF1 reduces the stomatal density and enhances root aerenchyma formation, impacting early leaf sheath development in rice [33], whereas MdEPF2 overexpression in apple significantly decreases the stomatal density [34].In ryegrass, ScEPF1 and ScEPF2 overexpression reduce the stomatal density and improve drought tolerance [35]. Additionally, Lu et al. reported that OsEPFL2 was highly expressed in rice panicles, and CRISPR/Cas9-mediated knockout of OsEPFL2 resulted in transgenic rice with short or no awns [18]. In this study, we detected high expression levels of multiple AsEPF/EPFL genes in oat roots and spikes, suggesting an evolutionarily conserved role of this gene family in root and panicle development within Poaceae plants. Future studies employing CRISPR/Cas9-mediated knockout experiments could further elucidate the molecular mechanisms by which these genes regulate panicle morphogenesis and stomatal development in oats.
With the growing global population and extreme climate events, crop production and food security face significant challenges. Plants must adapt to harsh conditions such as drought and soil salinity, which present significant challenges to crop growth and threaten global food security [36]. Salt stress is a major abiotic factor limiting crop productivity, with plant-specific EPF/EPFL peptide hormone families, playing crucial roles in stress adaptations through their regulation on stomatal development and abiotic stress signaling pathways.In this study, by integrating the transcriptomic data of monocotyledonous plants under salt stress from the GEO database (GSE21651), we found that Os08g0485500 in the salt-tolerant rice variety and its homologous genes in oat (3AG0421080, 3CG0493720 and 3DG0522310) were significantly upregulated in a synchronized manner after 24 h of NaCl treatment, indicating conserved role of the EPF/EPFL family in salt stress adaptation.
Furthermore, we found that the high expression of 2CG0320600 and 2AG0242220 under drought stress may be associated with the enhancement of plant stress tolerance.The oat genes 2CG0320600 and 2AG0242220 were phylogenetically clustered into the same evolutionary subgroup as AtEPF1 and AtEPF2 in A. thaliana. This evolutionary congruence implies functional conservation, particularly in stomatal density regulation, with oat 2CG0320600 and 2AG0242220 likely performing analogous roles to Arabidopsis AtEPF1/AtEPF2 [37]. In Arabidopsis, AtEPF1 and AtEPF2 encode canonical regulators of stomatal patterning. Mechanistically, these EPF family peptides interact with the receptor kinase TMM (TOO MANY MOUTHS) and ERECTA family receptors to suppress stomatal lineage initiation [38]. For example, AtEPF2 overexpression reduces the stomatal density, whereas atepf2 mutants present hyperstomatal phenotypes. The evidence suggests that these genes modulate the stomatal density to mitigate the effects of drought and salinity conditions. By extension, oat 2CG0320600 and 2AG0242220 may orchestrate similar adaptive mechanisms, enhancing stress tolerance through conserved stomatal developmental pathways.Future studies employing overexpression experiments could further elucidate the molecular mechanisms through which these genes regulate stomatal development in oats.
Conclusions
This study identified 33 AsEPF/EPFL genes from the oat genome using bioinformatics data, analysed their gene structures and conserved protein domains, and investigated the evolutionary relationships of EPF/EPFL genes across species. An analysis of the expression patterns of AsEPF/EPFL genes in different tissues (roots, stems, leaves, and spikes) suggested that AsEPF/EPFL genes play important roles in root, leaf, and spike morphogenesis in oats. Additionally, the expression characteristics of 33 AsEPF/EPFL genes under drought and salt stress were analysed, revealing the significant induction of AsEPF/EPFL expression with a prolonged stress duration. Based on the conserved functions of homologous genes in other species, candidate genes potentially involved in the oat drought stress response were identified. This research lays a foundation for further elucidation of the biological functions of the oat EPF/EPFL gene family.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Liu S, Jia F, Jiao, Z,Wang J. Ectopic expression of secretory peptide Pd EPF 3 in Arabidopsis confers drought tolerance with reduced stomatal density. Acta Soc Bot Pol. 2019,88(2).
- 2Zhon L, Fang H. Identification and bioinformatics analysis of EPF/EPFL family genes for epidermal pattern factors in tomato. Mol Plant Breed. 2024,1–13.
- 3Lu J, He J, Zhou X, Zhong J, Li J, Liang YK. Homologous genes of epidermal patterning factor regulate stomatal development in rice. J Plant Physiol. 2019 Mar-Apr;234–235:18–27.10.1016/j.jplph.2019.01.01030660943 · doi ↗ · pubmed ↗
- 4Hunt L, Bailey KJ, Gray JE. The signalling peptide EPFL 9 is a positive regulator of stomatal development. New Phytol. 2010, 186(3).10.1111/j.1469-8137.2010.03200.x 20149115 · doi ↗ · pubmed ↗
- 5Jin L. Preliminary study on the involvement of rice epidermal pattern factor EPF in regulating stomatal development. Wuhan Univ. 2019,2–3.
- 6Qi J. Response of Apple EPFL secreted protein gene to long-term drought stress and functional analysis of Md EPF 2. Northwest Agriculture and Forestry University; 2020.