Changes of Microbiome in Human Papillomavirus Infection and Cervical Cancer: A Systematic Review and Meta‐Analysis
Wei Zhang, Yan Ge, Lihe Yao, Qingchun Yan, Jiuju Wei, Yanfei Yin, Bin Liu

TL;DR
This study reviews how the microbiome changes in HPV infection and cervical cancer, finding significant differences in microbial diversity and composition between infected and healthy individuals.
Contribution
The study provides a comprehensive meta-analysis of microbiome changes across different stages of HPV infection and cervical cancer.
Findings
HPV-infected patients show significantly different microbial diversity compared to healthy controls.
Gardnerella is prevalent during HPV infection but decreases with cervical lesions, while Prevotella, Porphyromonas, and Dialister increase in cancer stages.
Microbial diversity varies with the severity of cervical lesions and is influenced by sample collection regions.
Abstract
We aimed to conduct a systematic review and meta‐analysis of high‐throughput sequencing studies to assess changes in microbiome alpha, beta diversity, and composition differences in patients with human papillomavirus (HPV) infection and cervical cancer. The PubMed, Embase, Web of Science, and Cochrane Library databases were systematically searched to include original studies. The effect size estimates with a 95% confidence interval were combined using a random effects model. The meta‐analysis was performed using the Stata MP16 software. A total of 64 studies were included, with a meta‐analysis of the diversity index performed on a subset of seven studies. Microbial diversity of patients infected with HPV was observed to be significantly different from that of healthy controls (CHAO index: 95% CI 0.42, 5.03, I 2 = 99.18%, p < 0.05). Subgroup analysis based on the sample collection…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
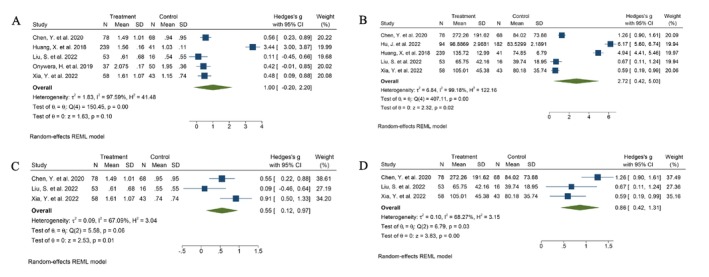 FIGURE 1
FIGURE 1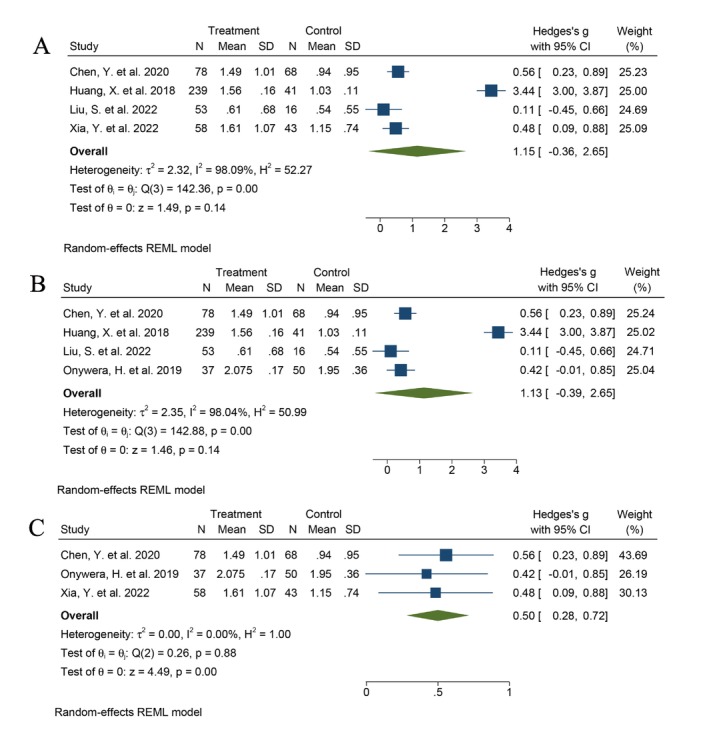 FIGURE 2
FIGURE 2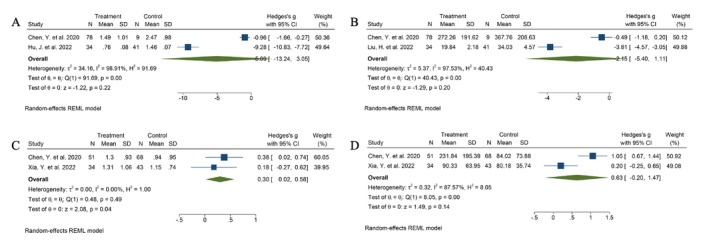 FIGURE 3
FIGURE 3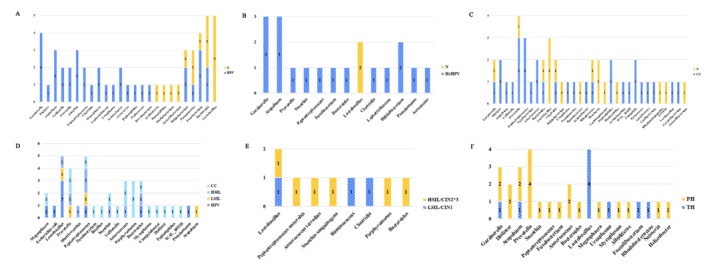 FIGURE 4
FIGURE 4| Study ID | Country | Study design | Study period | Total number | Intervention | Age | Disease type | Sample collection | Method to determine microbiota | Sequencing platform |
|---|---|---|---|---|---|---|---|---|---|---|
| Selvaraj Arokiyaraj et al. 2018 | Korea | CS | 2006–2013 |
41 N = 10 TH = 15 PH = 16 | — |
44.6 ± 9.0 44.4 ± 13.3 44.6 ± 13.0 | HPV | Vagina |
16S rRNA — | Roche 454 |
| Audirac et al. 2016 | Mexico | CCS | 2008–2011 |
32 NCL = 17 SIL = 4 CC = 8 | — |
34 ± 8 40 ± 14 43 ± 11 | CC | Vagina |
16S rRNA V3‐V4 | Roche 454 |
| Berggrund et al. 2020 | Sweden | CS | 2013–2015 |
96 N = 32 TH = 26 PH = 38 | — |
36.7 ± 5.2 35.9 ± 5.2 37.5 ± 6.0 | HPV | Vagina | Metagenomes | Ion Torrent |
| Bi et al. 2021 | China | CCS | 2018.01–2018.06 |
15 PH = 6 TH = 4
| — |
52.17 ± 10.19 43.00 ± 4.74 40.00 ± 6.89 | HPV | Vagina |
16S rRNA V4 | Ion S5XL |
| Borgogna et al. 2021 | USA | CCS | 2010–2011 | 40 | — | 29 ± 9.1 | HPV | vagina |
16S rRNA V3‐V4 | Roche 454 |
| Camargo et al.2022 | Colombia | CS | 2007–2010 | 66 | — | 42.5 (20) | HPV | Vagina |
16S rRNA V4 | Illumina Novaseq PE250 |
| Chao et al. 2020 | China | CCS | 2018.07–2019.03 |
329PH =59 TH = 139
| — | 41.68 ± 11.58 | HPV | Vagina |
16S rRNA V4 | Illumina HiSeq2500 |
| Chao et al. 2021 | China | CCS | 2018.07–2019.03 |
272 CIN2 + =83 HPV = 86
| — | 38.93 ± 9.59 |
HPV HSIL | Vagina |
16S rRNA V5 | Illumina HiSeq2500 |
| Chao et al.2019 | China | CCS | 2018.5–2018.5 |
151 HPV = 65
| — | 37.86 ± 9.99 | HPV | Vagina |
16S rRNA V5 | Illumina HiSeq2500 |
| Chen et al. 2020 | China | CCS | 2015.5–2016.11 |
229 HPV = 78 LSIL = 51 HSIL = 23 CC = 9
| — |
43.00 ± 8.69 47.78 ± 9.63 46.00 ± 10.19 43.70 ± 10.74 56.11 ± 9.02 |
HPV CC | Vagina |
16S rRNA V3‐V4 | Illumina MiSeq |
| Cheng et al.2020 | Sweden | CCS | — |
257 HPV = 144
| — | 14–29 | HPV | Vagina |
16S rRNA V3‐V4 | Illumina MiSeq |
| Cheng et al. 2020 | China | CCS | 2016.9–2019.6 |
131 LSIL = 26 HSIL = 40 CC = 32
| — | 21–65 |
HPV CC | Vagina |
16S rRNA V4 | Illumina HiSeq2000 |
| Chorna et al. 2020 | USA | CCS | — |
19 HrHPV = 11
| — | — | HrHPV |
Cervix vagina Urine | Metagenomes | — |
| Dareng et al. 2016 | Nigeria | CCS | 2012.4–2012.8 |
278 HrHPV = 66
| — | ≥ 18 | HrHPV | Vagina |
16S rRNA V4 | Illumina MiSeq |
| Di Paola et al.2017 | Italy | CS | — |
73 TH = 27 PH = 28
| — | 26–64 | HPV | Vagina |
16S rRNA V3‐V5 | Roche 454 |
| Fang et al. 2022 | China | CCS | 2020.12–2021.9 |
40 HrHPV = 20
| — |
33.75 ± 6.78 35.55 ± 4.12 | HrHPV | Cervix |
16S rRNA V3‐V4 | Illumina NovaSeq |
| Godoy‐Vitorino et al. 2018 | USA | CCS | — |
62 HrHPV = 27 LrHPV = 3 Both HrHPV and LrHPV = 22
| — | — | HPV |
Cervical vaginal rectum |
16S rRNA V4 | Illumina |
| Guo et al.2022 | China | CCS | — |
149 HPV + NoSIL = 40 HPV + LSIL = 28 HPV + HSIL = 51
| — | 19–50 |
HPV SIL | Vagina |
16S rRNA V4‐V5 | Illumina NovaSeq |
| Hu et al. 2022 | China | CCS | 2019 |
276 HPV = 94
| — | 44.68 ± 10.94 | HPV | Cervix |
16S rRNA V3‐V4 | Illumina HiSeq |
| Huang et al. 2018 | China | CCS | 2015.4–2016.11 |
280 16LSIL = 30 16HSIL = 43 52LSIL = 42 52HSIL = 42 58LSIL = 42 58HSIL = 40 N = 41 | — | — | HPV | Cervix |
16S rRNA V4‐V5 | Illumina MiSeq |
| Ivanov et al. 2023 | Russia | CCS | — | 241 |
Different cancer therapies CRT; RT; ST; CT | — |
HPV CC | Cervix |
16S rRNA V3‐V4 | Illumina MiSeq |
| Kang et al. 2021 | Korea | CCS | — |
23 CC = 8 CIN = 8
| — |
47 ± 10.2 43.4 ± 12.8 47.4 ± 5.38 | CC | Vagina |
16S rRNA V3 | Ion Torrent |
| Laniewski et al. 2018 | USA | CCS | — |
100 N = 20 HPV = 31 LSIL = 12 HSIL = 27 CC = 10 | — |
39.55 ± 7.35 |
HPV CC | Vagina |
16S rRNA V4 | Illumina MiSeq |
| Laniewski et al. 2019 | USA | CCS | — |
78 N = 18 HPV = 11 LSIL = 12 HSIL = 27 CC = 10 | — |
40.38 ± 6.98 36.36 |
HPV CC | Vagina |
16S rRNA V4 | Illumina MiSeq |
| Lee et al. 2013 | Korea | CCS | 2005–2009 |
68 N = 23 HPV = 45 | — | — | HPV | Vagina |
16S rRNA V2‐V3 | Roche 454 |
| Lee et al. 2020 | Korea | CCS | — |
66 CIN1− =24 CIN2 + =42 | — | 45.1 ± 11.7 | HPV | Vagina |
16S rRNA V3 | Ion Torrent |
| Lin et al. 2022 | China | CCS | 2018.6–2020.3 |
28 HPV = 23
| — |
39.5 ± 11.1 36.6 ± 9.3 | HPV | Cervix |
16S rRNA V4 | Illumina MiSeq |
| Liu et al. 2022 | China | CCS | — |
82 Zang = 42 Naxi = 13 Yi = 7 Bai = 4 Lisu = 6 Han = 10 | — | 39.7 ± 7.1 | HPV |
Cervical vaginal rectum |
16S rRNA — | Illumina MiSeq |
| Liu et al. 2022 | China | CCS | 2020.6–2021.3 |
115 HPV = 34 CIN = 40 CC = 41 | — | — |
HPV CIN CC | Cervix | Metagenomes | Illumina HiSeq2500 |
| Liu et al. 2022 | China | CCS | 2016.11–2017.7 |
69
SHPV = 31 MHPV = 22 | — |
35.75 ± 7.94 39.55 ± 10.05 37.82 ± 9.21 | HPV | Vagina |
16S rRNA V4 | Illumina MiSeq |
| Liu et al. 2023 | China | CCS | 2018.7–2019.1 |
1015 HPV = 271
| — | 45.1 ± 7.20 | HPV | Vagina |
16S rRNA V4 | Illumina MiSeq |
| Ma et al. 2023 | China | CCS | 2021.4–2021.7 |
160 NCL = 22 LSIL = 45 HSIL = 36 CC = 27
| — | 30 (29–75) |
HPV SIL CC | Vagina |
16S rRNA — | Illumina MiSeq |
| McKee et al. 2020 | USA | CCS | — |
308 ACC = 109 HrHPV = 110
| — | 26 (21–39) | HPV | Vagina |
16S rRNA V4 | Illumina MiSeq |
| Mei et al. 2022 | China | CS | 2015.1–2021.3 |
100
PH = 28 TH = 30 | — |
37.29 ± 7.49 38.47 ± 7.37 38.86 ± 11.44 | HrHPV | Vagina |
16S rRNA V3‐V4 | Illumina MiSeq |
| Mitra et al. 2015 | UK | CCS | — |
169 LSIL = 52 HSIL = 92 CC = 5
| — | 31 ± 5.08 |
HPV SIL CC | Vagina |
16S rRNA V1‐V2 | Illumina MiSeq |
| Mitra et al. 2020 | UK | CS | 2002–2007 |
87 PH = 45 TH = 42 | — | 20.5 ± 2.4 |
HPV CIN2 | Cervix |
16S rRNA V1‐V2 | Illumina MiSeq |
| Nieves‐Ramirez et al. 2021 | Canada | CCS | 2003.12–2006.07 |
228 SIL = 121
| — |
37.26 ± 10.87 42.83 ± 7.92 |
HPV SIL | Vagina |
16S rRNA V3 | Illumina HiSeq2000 |
| Onywera et al. 2019 | South Africa | CCS | — |
87 HPV = 37
| — | 32.0 (25.0–39.0) | HPV | Cervix |
16S rRNA V3‐V4 | Illumina MiSeq |
| Piyathilake et al. 2016 | USA | CCS | — |
430 CIN 1 = 90 CIN 2+ = 340 | — | 26.1 ± 4.7 |
HPV CIN | Cervix |
16S rRNA V4 | Illumina MiSeq |
| Ritu et al. 2019 | China | CS | 2016.8–2017.8 |
133 N = 42 TH = 56 PH = 35 | — | 43.8 ± 8.9 | HPV | Cervix |
16S rRNA V4 | Illumina Hiseq2500 |
| Sasivimolrattana et al. 2022 | Thailand | CCS | — |
48 HPV = 43 N = 5 | — | 37.77 ± 1.25 | HrHPV | Cervix |
16S rRNA V1‐V9 | Illumina MiSeq |
| Shi et al. 2022 | China | CS | 2015.4–2016.10 |
73 TH = 45 PH = 28 | — | 40.1 ± 11.5 |
HPV SIL | Cervix |
16S rRNA V4‐V5 | Illumina MiSeq |
| Sims et al. 2020 | Botswana | CCS | 2018.7–2019.2 |
31 Dysplasia = 21 CC = 10 | — |
41.8 ± 7.5 50.7 ± 12 | CC | Cervix |
16S rRNA V4 | Illumina MiSeq |
| So et al. 2020 | Korea | CCS | — |
50 HrHPV = 40 NHrHPV = 10 | — | 20–50 | HPV | Vagina |
16S rRNA V3‐V4 | Illumina MiSeq |
| Tango et al. 2020 | Korea | CCS | 2006.3–2020 |
92 CIN2/3‐CC = 42
| — |
45.7 ± 11.7 45.1 ± 11.6 | CC | Cervix |
16S rRNA — | Roche 454 |
| Teka et al. 2023 | Ethiopia | CCS | 2019.10–2020.2 |
120 CC = 60 Dysplasia = 25
| — | ≥ 18 | CC | Cervix |
16S rRNA V4 | Illumina MiSeq |
| Tosado‐Rodríguez et al. 2023 | USA | CCS | 2020.3–2020.10 |
91
LSIL = 18 HSIL = 17 | — | 39 ± 11.3 |
HPV SIL | Cervix |
16S rRNA V4 | — |
| Usyk et al. 2020 | USA | CS | — |
273 THr = 70 PHr = 170 CHr = 33 | — |
23.4 ± 2.8 22.6 ± 2.3 22.70 ± 2.9 | HrHPV | Cervix |
16S rRNA V4 | Illumina MiSeq |
| Vikramdeo et al. 2022 | USA | CCS | — |
36 AA = 12 CA = 12 HIS = 12 | — |
35.25 ± 8.00 44.42 ± 12.32 32.92 ± 9.62 |
CIN CC | Cervix |
16S rRNA V4 | Illumina MiSeq |
| Wei et al. 2022 | China | CCS | — |
59
HPV = 13 LSIL = 15 HSIL = 10 CC = 11 | — |
38.50 ± 5.68 37.08 ± 7.05 38.33 ± 7.35 40.50 ± 5.95 43.45 ± 5.50 |
HrHPV SIL CC | Vagina |
16S rRNA V3‐V4 | Illumina MiSeq PE300 |
| Wei et al. 2021 | China | CCS | 2018.9–2018.12 |
60 HrHPV = 30
| — |
36.70 ± 6.30 34.13 ± 6.43 | HrHPV |
Cervix Vagina |
16S rRNA V3‐V4 | Illumina MiSeq PE300 |
| Wu et al. 2020 | China | CCS | — |
69 NILM = 31 LSIL = 22 HSIL = 16 | — | 38 ± 9.04 |
HrPV SIL | Vagina |
16S rRNA V3‐V4 | Illumina MiSeq |
| Wu et al. 2021 | China | CCS | 2018.1–2019.12 |
94 CC = 13 HSIL = 31 LSIL = 10 HPV = 12
| — | 38.6 ± 7.6 |
HPV SIL CC | Vagina |
16S rRNA V4 | Illumina NovoSeq6000 |
| Xia et al. 2022 | China | CCS | 2021.4–2021.10 |
135
HrHPV = 58 LSIL = 34 | — | 32.27 ± 5.88 | HPV | Vagina |
16S rRNA V3‐V4 | Illumina NovoSeq6000 |
| Xie et al. 2020 | China | CCS | 2018.7–2019.8 |
72
CIN = 22 CC = 23 | — | 34.57 ± 2.665 |
CIN CC | Vagina |
16S rRNA V4 | Illumina MiSeq |
| Xu et al. 2022 | China | CCS | — |
40
LSIL = 10 HSIL CC = 10 | — | 48.28 ± 12.76 |
SIL CC |
Cervix Vagina |
16S rRNA V3‐V4 | Illumina MiSeq |
| Xu et al. 2023 | China | CCS | — |
87 HRCD = 50
| — | 18–75 |
HPV CIN CC | Vagina |
16S rRNA V3 | Illumina MiSeq |
| Yang et al. 2020 | China | CCS | 2017.2–2018.11 |
52 HPV16 = 27
| — |
34.5 ± 4.5 36.9 ± 4.9 | HPV | Vagina | Metagenomes | Illumina HiSeq |
| Zeng et al. 2023 | China | CS | 2016.1–2018.6 |
135
HrHPV16/18 = 28 Non‐HrHPV16/18 = 62 |
Vaginal probiotics (mainly lactobacilli) interferon therapy |
36 (17) 37 (18) 41.5 (10.75) | HPV | Cervix |
16S rRNA — | Ion S5XL |
| Zeng et al. 2023 | China | CCS | 2019.9–2020.1 |
60
CIN1 = 15 CIN2/3 = 15 CC = 15 | — | — |
HPV CIN | Vagina |
16S rRNA V3‐V4 | Illumina MiSeq |
| Zhai et al. 2021 | China | CCS | 2019.1–2019.12 |
168
HrHPV = 29 LSIL = 32 HSIL = 40 CC = 38 | — |
40.08 ± 4.83 42.17 ± 5.18 40.63 ± 4.55 40.64 ± 5.57 42.43 ± 5.31 |
HrHPV SIL CC | Vagina |
16S rRNA V3‐V4 | Ion S5XL |
| Zhang et al. 2018 | China | CCS | 2014–2015 |
166 CIN1− = 64 CIN1 = 62 CIN2+ = 19 CIN3 = 21 | — | — |
HPV CIN | Cervix |
16S rRNA V3‐V5 | Illumina HiSeq2500 |
| Zhang et al. 2022 | China | CCS | 2020.11–2021.5 |
356
HPV = 159 CIN = 84 | — | 20–70 |
HPV CIN CC |
Cervix Vagina |
16S rRNA — | Illumina Novaseq PE250 |
| Zhang et al. 2021 | China | CCS | — |
100
Other HrHPV = 32 HPV16/18 = 38 CC = 10 | — |
38.35 ± 3.72 36.75 ± 6.15 35.73 ± 6.89 38.80 ± 3.08 |
HPV CC |
Cervix vagina |
16S rRNA V3‐V4 | Illumina MiSeq |
| Study ID | Software | Alpha | Beta | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Alpha richness and diversity | Shannon | Simpson | Chao | Observed_species | ACE | PD whole tree | Beta diversity | PCA | PCoA | Others | ||
| Selvaraj Arokiyaraj et al. 2018 | Mothur | Higher PH and TH than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | Difference between N, TH, and PH | ─ | ⊕ | ─ |
| Audirac et al. 2016 | QIIME | Higher CC and SIL groups than NCL | ⊙ | ─ | ─ | ─ | ─ | ⊕ | No difference between CC, SIL groups and NCL | ─ | ⊕ | ─ |
| Berggrund et al.2020 | QIIME | 1. No significant difference between N, TH and PH at baseline | ⊙ | ─ | ─ | ─ | ─ | ─ | NR | ─ | ─ | ─ |
| 2. No significant difference between TH and PH at second sample | ⊙ | |||||||||||
| Bi et al. 2021 | QIIME | Higher PH than TH and N | ⊕ | ⊕ | ⊕ | ⊕ | ⊕ | ─ | NR | ─ | ─ | ─ |
| Camargo et al. 2022 | QIIME | 1. Higher low VL than high VL and medium VL | ⊕ | ⊕ | ─ | ─ | ⊕ | ─ | 1. Difference between low VL, high VL and medium VL | ─ | ⊕ | ─ |
| 2. Higher increase VL than equal VL | ⊙ | ⊙ | ─ | ─ | ⊕ | ─ | 2. No difference between increase VL, equal VL and decrease VL | ─ | ⊙ | ─ | ||
| Chao et al. 2020 | — | No difference between PH, TH, and N | ⊙ | ─ | ─ | ─ | ─ | ─ | 1. Difference between PH and TH | ─ | ⊕ | ─ |
| 2. Difference between PH and N | ─ | ⊕ | ─ | |||||||||
| 3. No difference between TH and N | ─ | ⊙ | ─ | |||||||||
| Chao et al. 2021 | — | 1. Higher CIN2+ than HPV | ⊕ | |||||||||
| 2. Higher CIN2+ than N | ⊕ | ─ | ─ | ─ | ─ | ─ | Difference between CIN2+, HPV and N | ─ | ⊕ | ─ | ||
| 3. No difference between P and N | ⊙ | |||||||||||
| Chen et al. 2020 |
Mothur R | 1. Higher HPV‐positive (HPV; LSIL; HSIL; CC) than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | Difference between HPV, LSIL, HSIL, CC and N | ─ | ⊕ | ─ |
| 2. CC were highest than HPV, LSIL, HSIL | ⊕ | |||||||||||
| Cheng et al. | R | Higher HPV than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | No difference between HPV and N | ─ | ⊙ | ─ |
| Cheng et al. 2020 | — | 1. Higher HPV than N | ⊕ | ─ | ─ | ─ | ─ | ⊙ | NR | ─ | ─ | ─ |
| 2. Higher CC than LSIL, HSIL, N | ⊕ | ─ | ─ | ─ | ─ | ⊕ | ||||||
| Chorna et al. 2020 | QIIME | 1. Higher HrHPV than N at vaginal | ─ | ─ | ⊕ | ─ | ─ | ─ | No difference between HrHPV and N at vaginal and cervical | ─ | ⊙ | ─ |
| 2. No difference between HrHPV and N at cervical | ─ | ─ | ⊙ | ─ | ─ | ─ | ||||||
| Dareng et al. 2016 | QIIME | ─ | ─ | ─ | ─ | ─ | ─ | ─ | Difference between HrHPV and N | ─ | ⊕ | ─ |
| Di Paola et al.2017 | R | Higher PH, TH than N | ⊙ | ─ | ⊙ | ⊙ | ─ | ─ | NR | ─ | ─ | ─ |
| Fang et al. 2022 | QIIME | Higher HrHPV than N | ⊕ | ⊕ | ─ | ─ | ─ | ─ | Difference between HrHPV and N | ─ | ⊕ | ─ |
| Godoy‐Vitorino et al. 2018 | R | Higher CIN3 than CIN1 at cervix and vaginal | ⊕ | ─ | ─ | ─ | ─ | ─ | Difference between cervix, vaginal and anal | ─ | ⊕ | ─ |
| Guo et al.2022 | R | 1. Higher HSIL than LSIL | ⊕ | ─ | ─ | ─ | ─ | ─ | Differences between N, HPV + NoSIL, LSIL andHPV+HSIL | ─ | ⊕ | ─ |
| 2. No differences between HPV + LSIL and HPV + NoSIL | ⊙ | ─ | ─ | ─ | ─ | ─ | ||||||
| 3. No difference between HPV + NoSIL and N | ⊙ | |||||||||||
| Hu et al. 2022 | QIIME | Higher HPV than N | ─ | ─ | ⊕ | ─ | ─ | ─ | NR | ─ | ─ | ─ |
| Huang et al. 2018 | QIIMER | Higher SILs than N | ⊕ | ─ | ⊕ | ─ | ⊕ | ─ | Difference between SILs and N | ─ | ⊕ | ─ |
| Ivanov et al. 2023 | QIIME | 1. Higher CC than N | ⊕ | ─ | ─ | ⊕ | ─ | ─ | ||||
| 2. Higher HSIL than N | ⊕ | NR | ─ | ─ | ─ | |||||||
| 3. Lower in PT+ than PT— | ⊙ | ─ | ─ | ⊙ | ─ | ─ | ||||||
| Kang et al. 2021 | QIIME | Higher CC, CIN than N | ⊙ | ⊙ | ─ | ─ | ─ | ─ | Difference between CC, CIN and N | ─ | ⊕ | ─ |
| Laniewski et al. 2018 | USEARCH | NR | ─ | ─ | ─ | ─ | ─ | ─ | No difference between HPV, CIN1, CIN2/3, CC and N | ─ | ⊙ | ─ |
| Laniewski et al. 2019 | USEARCH | NR | ─ | ─ | ─ | ─ | ─ | ─ | Difference between HPV, LSIL, HSIL, CC and N | ⊕ | ─ | ─ |
| Lee et al. 2013 | QIIME | NR | ─ | ─ | ─ | ─ | ─ | ─ | NR | ─ | ─ | ─ |
| Lee et al. 2020 |
QIIME VSEARCH | NR | ─ | ─ | ─ | ─ | ─ | ─ | No difference between CIN1− and CIN2+ | ⊙ | ─ | ─ |
| Lin et al. 2022 | R | Higher HPV than N | ⊕ | ⊕ | ⊙ | ─ | ⊙ | ─ | NR | ─ | ─ | ─ |
| Liu et al. 2022 | Mothur | 1. Similar among all samples from the six ethnic groups | ─ | ─ | ─ | ─ | ─ | ─ | 1. Difference between rectal and cervical, vaginal | ⊕ | ─ | ─ |
| 2. Highest in the rectal and was higher in cervical than in vaginal | ─ | 2. No difference between cervical and vaginal | ⊙ | ─ | ─ | |||||||
| 3. No difference between HPV and N at rectal, cervical, vaginal | ⊙ | ─ | ─ | |||||||||
| Liu et al. 2022 | R | 1. Higher CIN and CC than HPV | ⊕ | ─ | ⊕ | ─ | ─ | ─ | Difference between CIN and CC, HPV | ─ | ⊙ | ─ |
| 2. No differences between CIN and CC | ─ | ─ | ⊙ | ─ | ─ | ─ | ||||||
| Liu et al. 2022 | Mothur | 1. No differences between SHPV, MHPV and N at genus level | ⊙ | ⊙ | ⊙ | ─ | ⊙ | ─ | Difference between MHPV and SHPV, N | ─ | ⊕ | ─ |
| 2. Higher SHPV than MHPV and N | ⊙ | ⊙ | ⊕ | ─ | ⊙ | ─ | ||||||
| Liu et al. 2023 | QIIME | 1. Higher BV & HPV than N | ⊕ | ─ | ─ | ⊕ | ─ | ─ | ||||
| 2. N, non‐BV and HPV, BV and HPV− to BV and HPV+, an increasing trend of alpha diversities | ─ | ─ | ─ | ─ | ─ | ─ | 1. Difference between BV and HPV and N | ─ | ⊕ | ─ | ||
| 3. Higher SHPV than N | ⊕ | ─ | ─ | ⊕ | ─ | ─ | 2. Difference between MHPV and SHPV | ─ | ⊕ | ─ | ||
| 4. Higher MHPV than N | ⊕ | ─ | ─ | ⊕ | ─ | ─ | ||||||
| 5. Higher MHPV than SHPV | ⊙ | ─ | ─ | ⊕ | ─ | ─ | ||||||
| Ma et al. 2023 | QIIME | Higher CC than LSIL, NV+ and N | ⊕ | ⊕ | ─ | ─ | ─ | ─ | NR | ─ | ─ | ─ |
| McKee et al. 2020 | Mothur | No difference between ACC, HrHPV and N | ⊙ | ⊙ | ─ | ─ | ─ | ─ | Difference between ACC, HrHPV and N | ─ | ─ | ⊕ |
| Mei et al. 2022 |
Mothur R | 1. Higher N than TH | ─ | ─ | ⊕ | ─ | ─ | ─ | ||||
| 2. No difference between PH and N | ─ | ─ | ⊙ | ─ | ─ | ─ | Difference between PH, TH and N | ─ | ⊕ | ─ | ||
| 3. No difference between PH and TH | ─ | ─ | ⊙ | ─ | ─ | ─ | ||||||
| Mitra et al. 2015 | MothurR | Higher CC, HSIL, LSIL than N | ⊙ | ⊙ | ─ | ⊙ | ─ | ─ | NR | ─ | ─ | ─ |
| Mitra et al. 2020 | MothurR | Higher PH than TH | ─ | ⊙ | ─ | ─ | ─ | ─ | NR | ─ | ─ | ─ |
| Nieves‐Ramirez et al. 2021 | Mothur | 1. Higher SIL than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | 1. Difference between SIL and N | ─ | ⊕ | ─ |
| 2. No difference between HPV_Lesion HPV‐no_Lesion | ⊙ | ─ | ⊙ | ─ | ─ | ─ | 2. No difference between HPV_Lesion HPV‐no_Leson | ─ | ⊕ | ─ | ||
| 3. Higher HPV_Lesion than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | 3. Difference between HPV and N | ─ | ⊕ | ─ | ||
| 4. Higher HPV than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | ||||||
| Onywera et al. 2019 | QIIME | 1. No difference between HPV and N | ⊙ | 1. No difference between HPV and N | ─ | ⊙ | ─ | |||||
| 2. No difference between HrHPV and N | ⊙ | ⊙ | ─ | ─ | ─ | ─ | 2. No difference between HrHPV and N | ─ | ⊙ | ─ | ||
| Piyathilake et al. 2016 | QIIME | No difference between CIN1 and CIN2+ | ─ | ─ | ─ | ─ | ─ | ─ | No difference between CIN1 and CIN2+ | ─ | ⊙ | ─ |
| Ritu et al. 2019 | QIIME | 1. Higher N than HPV | ⊙ | ⊙ | ⊕ | ⊕ | ─ | ⊙ | ||||
| 2. No difference between TH and PH | ⊙ | ⊙ | ⊙ | ⊙ | ─ | ⊙ | NR | ─ | ─ | ─ | ||
| Sasivimolrattana et al. 2022 | — | Higher NLD than LD | ⊕ | ─ | ─ | ─ | ─ | ─ | Difference between NLD and LD | ─ | ─ | ─ |
| Shi et al. 2022 | R | No difference between PH and TH | ⊙ | ─ | ⊙ | ─ | ─ | ─ | No difference between PH and TH | ─ | ⊙ | ─ |
| Sims et al. 2020 | USEARCH | 1. Higher CC than Dysplasia | ⊕ | 1. Difference between CC and Dysplasia | ─ | ⊕ | ─ | |||||
| 2. Higher CIN3 than CIN2 | ⊕ | ─ | ─ | ─ | ─ | ─ | 2. Difference between CIN3 and CIN2 | ─ | ⊕ | ─ | ||
| So et al. 2020 | QIIME | Higher HPV than N | ⊕ | ─ | ─ | ─ | ─ | ─ | Difference between CC, CIN and N | ─ | ⊕ | ─ |
| Tango et al. 2020 | USEARCH | Higher CIN2/3‐CC than N | ⊙ | ⊙ | ⊕ | ─ | ─ | ─ | No difference between CIN2/3‐CC and N | ─ | ⊙ | ─ |
| Teka et al. 2023 | QIIME | 1. No difference between HPV and N | ⊙ | ⊙ | ─ | ─ | ─ | ─ | 1. Difference between HPV and N | ─ | ⊕ | ─ |
| 2. Higher CC than Dysplasia and N | ⊕ | ⊕ | ─ | ─ | ─ | ─ | 2. Difference between CC and Dysplasia, N | ─ | ⊕ | ─ | ||
| Tosado‐Rodríguez et al. 2023 | QIIME | 1. No difference between HPV and N | ⊙ | ─ | ⊙ | ─ | ─ | ─ | Difference between HSIL and LSIL | ─ | ⊕ | ─ |
| 2. Higher HSIL than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | ||||||
| Usyk et al. 2020 | VSEARCH | 1. No difference between THr and PHr and CHr at visit 1 | ⊙ | ─ | ─ | ─ | ─ | ─ | NR | ─ | ─ | ─ |
| 2. Higher CHr than PHr and THr at visit 2 | ⊕ | ─ | ─ | ─ | ─ | ─ | ||||||
| Vikramdeo et al. 2022 | USEARCH | 1. Higher CIN than N | ⊙ | ─ | ─ | ⊕ | ─ | ─ | ||||
| 2. No difference between HPV and N | ⊙ | ─ | ─ | ⊙ | ─ | ─ | NR | ─ | ─ | ─ | ||
| Wei et al. 2022 | QIIMER | 1. Higher CC than N | ⊕ | |||||||||
| 2. Higher HSIL than N | ⊕ | ⊕ | ─ | ─ | ─ | ─ | 1. Difference between CC and N | ─ | ⊕ | ─ | ||
| 3. Higher LSIL than N | ⊕ | |||||||||||
| 4. Higher HPV than N | ⊕ | |||||||||||
| 5. No difference between CC and HSIL, LSIL, HPV | ⊙ | ⊙ | ─ | ─ | ─ | ─ | 2. No difference between HSIL and LSIL | ─ | ⊙ | ─ | ||
| Wei et al. 2021 | QIIME | No difference between HPV and N | ⊙ | ─ | ⊙ | ⊙ | ─ | ─ | Difference between HPV and N | ⊕ | ─ | ─ |
| Wu et al. 2020 | — | Higher NILM than LSIL and HSIL | ⊙ | ⊙ | ─ | ─ | ─ | ─ | NR | ─ | ─ | ─ |
| Wu et al. 2021 | QIIME | 1. Higher CC than N | ⊕ | |||||||||
| 2. Higher HSIL than N | ⊕ | ⊕ | ─ | ─ | ─ | ─ | Difference between CC, HSIL, LSIL, HPV and N | ─ | ⊕ | ─ | ||
| 3. Higher LSIL than N | ⊕ | |||||||||||
| 4. Higher N than HPV | ⊕ | |||||||||||
| Xia et al. 2022 | QIIMER | 1. Higher HrHPV than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | No difference between HrHPV and LSIL, N | ─ | ⊙ | ─ |
| 2. Higher HrHPV than LSIL | ⊙ | ─ | ⊕ | ─ | ─ | ─ | ||||||
| Xie et al. 2020 | QIIME | 1. No difference between CIN and CC | ⊙ | ⊙ | ─ | ─ | ─ | ─ | Difference between CIN, CC and N | ─ | ⊕ | ─ |
| 2. Higher CIN and CC than N | ⊙ | ⊙ | ─ | ─ | ─ | ─ | ||||||
| Xu et al. 2022 | QIIME | Higher CC than N | ⊕ | ⊕ | ─ | ─ | ─ | ─ | Difference between CC, HSIL, LSIL and N | ─ | ⊕ | ─ |
| Xu et al. 2023 | QIIME | Higher HRCD than N | ⊕ | ⊕ | ─ | ─ | ⊙ | ─ | No difference between HRCD and N | ─ | ⊙ | ⊙ |
| Yang et al. 2020 | R | No difference between HPV16 and N | ⊙ | ⊙ | ─ | ─ | ─ | ─ | No difference between HPV16 and N | ⊙ | ─ | ─ |
| Zeng et al. 2023 |
QIIME VSEARCH | 1. Higher HrHPV16/18 than N | ⊙ | ⊙ | ⊕ | ⊕ | ⊕ | ─ | 1. No difference between HrHPV16/18, HrHPV16/18 and N | ─ | ⊙ | ─ |
| 2. Higher MHPV and twoHPV than N | ⊙ | ⊙ | ⊙ | ⊙ | ⊙ | ⊙ | 2. Higher MHPV and twoHPV than N | ─ | ⊙ | ─ | ||
| 3. Higher PH than TH | ⊙ | ⊙ | ⊕ | ⊕ | ⊙ | ⊙ | ||||||
| Zeng et al. 2023 | USEARCHMothur | 1. Higher CC than N | ⊕ | ⊕ | ⊕ | ⊕ | ─ | ⊕ | 1. No difference between CIN1 and N | ─ | ⊙ | ─ |
| 2. Higher CC than CIN1 | ⊙ | ⊙ | ⊕ | ⊕ | ─ | ⊕ | 2. Difference between CC and CIN1, CIN2/3 N | ─ | ⊕ | ─ | ||
| 3. Higher CC than CIN2/3 | ⊕ | ⊕ | ⊕ | ⊕ | ─ | ⊕ | 3. No difference between CIN2/3 and CC | ─ | ⊙ | ─ | ||
| Zhai et al. 2021 | QIIME | 1. Higher N than HrHPV | ⊕ | ⊕ | ⊕ | ⊕ | ⊕ | ⊙ | Difference between CC, HSIL, LSIL, HrHPV and N | ─ | ⊕ | ─ |
| 2. No difference between SIL and CC | ⊙ | ⊙ | ⊙ | ⊙ | ⊙ | ⊙ | ||||||
| Zhang et al. 2018 | UPARSEQIIME | 1. No difference between CIN1− and CIN2+ | ⊙ | ─ | ─ | ─ | ─ | ─ | 1. No difference between CIN1− and CIN2+ | ─ | ⊙ | ─ |
| 2. No difference between CIN1− and CIN3 | ⊙ | ─ | ─ | ─ | ─ | ─ | 2. No difference between CIN1− and CIN3 | |||||
| Zhang et al. 2022 | MothurQIIME | 1. Higher CIN than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | No difference between HPV, CIN and N | ─ | ⊕ | ─ |
| 2. Higher HPV than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | ||||||
| 3. Higher SHPV than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | ||||||
| 4. Higher MHPV than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | ||||||
| 5. Higher CIN than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | ||||||
| 6. Higher HrHPV than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | ||||||
| 7. Higher LrHPV than N | ⊕ | ─ | ⊕ | ─ | ─ | ─ | ||||||
| Zhang. et al. 2021 |
CutadaptPEAR Prinseq Usearch Uchime Muscle FastTree Mothur R | 1. Higher cervial N than vaginal N | ⊕ | |||||||||
| 2. Higher cervial Other Hrhpv than vaginal Other Hrhpv | ⊕ | ─ | ─ | ─ | ─ | ─ | Difference between the cervical and vaginal microbiota | ─ | ⊕ | ─ | ||
| 3. Higher cervial Hrhpv than vaginal Hrhpv | ⊕ | |||||||||||
| 4. No difference between cervial CC and vaginal CC | ⊙ | ─ | ─ | ─ | ─ | ─ | ||||||
- —Lanzhou Science and Technology Bureau 10.13039/501100007764
- —Natural Science Foundation of Gansu Province 10.13039/501100004775
- —Health Commission of Gansu Province 10.13039/100017956
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCervical Cancer and HPV Research · Reproductive tract infections research · Gut microbiota and health
Introduction
1
Cervical cancer (CC) is the fourth most common cancer threatening women's health worldwide, with approximately 600,000 cases and 300,000 deaths annually [1]. Over 99% of precancerous lesions are associated with human papillomavirus (HPV) infection, with high‐risk HPV types 16, 18, 31, 33, and 45, which are identified in approximately 97% of CC cases [2]. Recently, many studies have found that an imbalance in the cervicovaginal microbiota is closely related to HPV infection and CC [3, 4, 5, 6].
The vagina is a relatively complex microecosystem mainly composed of vaginal flora, immune mediators, periodically changing endocrine environments, and reproductive tract mucosal epithelial immune cells. The vaginal flora is of great significance in maintaining the stability of microecology, self‐purification of the vagina, and women's health [7]. Women with dysbiosis are at an increased risk of developing tumors [8].
Various tumors meet the nutritional demands of rapid proliferation through changes in metabolic patterns. Metabolic reprogramming is considered a hallmark of cancer that promotes rapid proliferation, survival, invasion, metastasis, and resistance to therapy, thereby supporting tumorigenesis [9]. When dysbiosis occurs, especially colonization by certain obligate anaerobes, enzymes and metabolites are produced, which damage the integrity of mucosal and epithelial cells and promote HPV infection and the continuous progression of cervical lesions [10]. Gardnerella and Streptococcus sp. are involved with CC at the microbial level [11].
Therefore, clarifying the role of cervicovaginal microbiota in HPV infection, cervical lesions, and CC is of great significance for prevention and treatment. The development of high‐throughput sequencing technology, 16S amplicon sequencing, and metagenomics has allowed accurate analysis of vaginal microorganisms through non‐culture methods, providing a new approach for the research, diagnosis, and treatment of HPV infections and cervical lesions [12]. However, high‐throughput sequencing is challenging due to the lack of standardization and harmonization in detection and analysis methods. These include differences in amplification regions, analysis software, calculation methods, and outcome metrics, as well as inconsistencies in the selection of clinical sample types, such as stool, urine, blood, and vaginal secretions. These limitations have led to inconsistent results and hindered the understanding of the roles of microorganisms in the development of CC [13, 14, 15]. Most studies have suggested that HPV infection and CC could lead to an increase in the diversity of the vaginal microbiome [6, 8, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25]. However, other studies have shown a decrease in microbial diversity [5, 26, 27, 28, 29, 30, 31, 32, 33].
The purpose of this review is to summarize the current evidence on the application of high‐throughput sequencing in studying the cervicovaginal microbiota and perform a meta‐analysis to assess changes in microbiome diversity in patients with HPV infection and CC. Evaluation of microbial dominance, abundance, and diversity seemingly implies microbial dynamics reflected by the abundance of Gardnerella, Prevotella, Porphyromonas, and Dialister species that correlate with women's cervicovaginal conditions. This review will provide in‐depth insight into cervicovaginal microbiota associated with cervical health and disease.
Methods
2
Data Sources and Search Strategy
2.1
The systematic review was conducted following PRISMA 2020 guidelines (Checklist S1). This review was registered at PROSPERO (ID: 42023459347) (https://www.crd.york.ac.uk/prospero/). The Cochrane Library, Web of Science, PubMed, and Embase databases were searched for studies published before June 2023. There is no definite time limit for the retrieval of literature. The search strategy includes the combination of MeSH terms and free text, and the MeSH terms include “Microbiota,” “Uterine Cervical Neoplasms,” and “Human Papillomavirus Viruses” to retrieve articles related to human HPV infection, CC, and microbiome (Table S1 for search strategy).
Inclusion and Exclusion Criteria
2.2
The inclusion criteria were as follows: (1) definite diagnosis of HPV infection or CC, aged over 15 years; (2) types of outcomes including the results of diversity analysis or linear discriminant analysis using high‐throughput sequencing; (3) studies published in any country with human subjects; (4) case–control studies, cross‐sectional studies, retrospective and prospective cohort studies, randomized and non‐randomized controlled trials, and exploratory observational studies; and (5) research samples including cervical or vaginal secretions.
Exclusion Criteria: Pregnant women, patients with HIV infection, animal research, conference abstracts, reviews, letters to the editor, case reports, or case series.
Data Selection and Extraction
2.3
Two authors (W.Z. and Y.G.) independently selected the titles and abstracts of the studies and reviewed the full texts and supplementary materials. Any discrepancies were resolved by consensus or arbitration with a third author (Y.Y.). Study characteristics, microbiome diversity, and bacterial groups enriched in diseased conditions were extracted from each included study.
Quality Assessment
2.4
Newcastle‐Ottawa Scale was used to assess the quality of cohort and case–control studies, divided into three domains and eight items, using a star system with a full score of nine stars; the risk of bias was then considered as high (< 5 points), moderate (6–7), or low (8–9). The evaluation results of all articles are shown in Table S2.
Strategy of Data Synthesis
2.5
In this study, we conducted a meta‐analysis of diversity indices. Data were extracted using means and standard deviations to calculate Hedges' g and the 95% confidence intervals (CI) used in the meta‐analysis. A random‐effect model was applied to obtain pooled effect size estimates, 95% CI, and p values by restricting the maximum likelihood estimator. Meta‐analysis was performed using Stata MP16 software, and statistical significance was determined at p < 0.05. Statistical heterogeneity was assessed using I ^2^ statistics. Subgroup analysis was performed when the estimated values showed a heterogeneity > 70%. Publication bias was not tested because fewer than 10 studies were eligible for the meta‐analysis of each outcome.
Results
3
Study Selection
3.1
The detailed study research and selection process are shown in the PRISMA flowchart (Figure S1). A total of 2166 pieces of literature were searched. After duplicate removal and assessment of the title and abstract, a total of 64 articles were eligible for full‐text analysis, and 7 studies were selected for the quantitative synthesis.
Study Characteristics
3.2
The current evidence on the application of high‐throughput sequencing was examined to investigate variation in the cervicovaginal microbiota and perform a meta‐analysis to assess changes in microbiome diversity in patients with HPV infection and CC. The characteristics of the 64 studies, including 54 case–control and 10 cohort studies, were described and published between 2013 and 2023. In total, 41 studies were from Asian countries, 12 studies were from North America, 6 were from Europe, 4 studies were conducted in Africa, and 1 study was from Latin America (Table 1). Most of the studies (60 out of 64) employed 16S rRNA sequencing for the generation of microbiome data, while the remaining four studies utilized metagenomics sequencing. The 16S rRNA sequencing‐based studies considered different amplification regions such as V3–V4 (n = 18), V4 (n = 21), V1–V9 (n = 1), V3 (n = 4), V1–V2 (n = 2), V2–V3 (n = 1), V3–V5 (n = 2), V4–V5 (n = 3), and V5 (n = 2) and six studies omitted amplification regions. The most extensive sequencing platforms were Illumina MiSeq, Illumina HiSeq, and Illumina NovoSeq, with three studies using the Ion Torrent sequencing platform and six studies using Roche 454 sequencing.
A total of 22 studies explored the differences between microbial changes in the long‐term process of HPV infection in different degrees of cervical lesions, followed by progression to CC [5, 6, 17, 18, 19, 23, 25, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47]. A total of 11 studies [3, 10, 15, 29, 48, 49, 50, 51, 52, 53, 54] have reported differences between persistent and transient HPV infections. Persistent HPV infection makes a complex microbial community, indicating that the microbial flora differs according to the infection state. High‐risk HPV (HrHPV) infection impacts the role of microbiota in the existence of cervical lesions [13, 23, 29, 32, 46, 53, 55, 56, 57, 58]. Four studies investigated whether infection with a single HPV genotype or multiple HPV genotypes was associated with diverse communities composed of vaginal microbiota [25, 54, 59, 60]. Different treatment methods were applied for CC and compared differences in cervicovaginal microorganisms after treatment [17]. Interferons combined with probiotics (Lactobacillus) were used to treat the HPV infection and attempted to determine the mechanism by which Lactobacillus protects the vagina [54].
Microbiome Diversity
3.3
Alpha diversity included Shannon (n = 53), Simpson (n = 24), CHAO (n = 23), observed species (n = 12), ACE (n = 8), and phylogenetic diversity whole tree (n = 6) indices. Beta diversity included principal component analysis (n = 5), principal coordinate analysis (n = 40), and other methods (n = 2) (Table 2).
Changes of Alpha Diversity in Patients Infected With HPV
3.4
Nine studies [8, 16, 20, 23, 25, 35, 36, 61, 62] showed that microbial diversity was higher in patients infected with HPV than in healthy controls, two studies [6, 51] showed the opposite result, and five studies [5, 21, 30, 32, 63] found no difference. No correlation was observed between alpha diversity and HPV infection in South African women [30], Ethiopian women [21], Hispanic/Latina (HIS), African Americans (AA) [5], and Hispanics living in Puerto Rico [63]. In contrast, among Asian women, although statistical uniformity is not complete, there is a consistent trend that the invasion of HPV increases the diversity of cervical and vaginal microorganisms [16, 20, 23, 25, 32, 35, 36, 61, 62]. Therefore, region and race cannot be ignored in such studies. Similarly, patients infected with HrHPV have a higher microbial diversity than the normal Asian population [25, 54, 56, 64], and only one study found no significant difference in microbial diversity between the two groups [46]. In addition, the sample collection site was one of the factors in microbial diversity; vaginal samples from patients with HrHPV had higher microbial diversity than those from healthy controls [64]; however, there was no difference in the cervix [13].
Another interesting phenomenon is observed in patients with persistent and transient HPV infections (HPV clearance status). Cervicovaginal microbial diversity shows no significant difference between those with persistent and transient HPV infection [29, 48, 51, 52, 53], but five studies showed the diversity of patients with persistent HPV infection was more complex than that of patients with transient HPV infection [10, 15, 49, 50, 54], although not statistically uniform. These results were affected by age and duration of HPV infection; the population in these cohort studies ranged from 18 to 70 years, and the duration of the studies ranged from 6 months to 2 years. Therefore, age‐dependent variations in microbiome makeup need to be considered while evaluating the correlation between HPV infection type/severity and microbiome diversity and comparing the same across various age groups.
Changes of Alpha Diversity in Patients With CC
3.5
The microbial diversity of patients with CC was not only higher than that of HPV‐infected patients [18, 35] but also significantly higher than that of healthy controls [6, 17, 21, 22, 23, 24, 34, 35, 36, 37]. In a study with participants of two age groups, when the age was > 50 years, the diversity of patients with CC was higher than that of healthy people. When the age was less than 50 years, the significant difference between the two groups was absent [31]. The diversity of CC samples from different sites (including cervix and vagina) showed no differences [47]. Although the microbial diversity of patients with cervical dysplasia and cervical intraepithelial neoplasia (squamous intraepithelial lesions) was also increased, the observed differences were not statistically significant [18, 23, 43, 46]. Moreover, the microbial diversity of patients with CC was higher than that of patients with cervical dysplasia [19, 21] and cervical intraepithelial neoplasia [24, 35, 36].
Meta‐Analyses
3.6
Shannon and Chao indices were used for quantitative analysis (n = 7). The Shannon index evaluates species evenness, and the Chao index focuses more on species richness. The Shannon index showed that the microbial diversity between patients infected with HPV and healthy controls was statistically insignificant (Hedges' g = 1.00, 95% CI 0.20 to 2.20, I ^2^ = 97.59%). The Chao index indicated statistical significance (Hedges' g = 2.72, 95% CI 0.42 to 5.03, I ^2^ = 99.18%) (Figure 1A,B). It can be inferred that the total number of HPV‐infected species was higher than that in healthy people. However, the difference in species evenness was not obvious.
Forest plot of the differences in alpha diversity between patients with HPV infection and healthy control: (A) Shannon index, (B) CHAO index; Subgroup analysis at vagina (C) Shannon index, (D) CHAO index.
The subgroup analysis was conducted according to the sample collection regions, and the I^2^ value decreased (Shannon index: Hedges' g = 0.55, 95% CI 0.12 to 0.97, I ^2^ = 67.09%, CHAO index: Hedges' g = 0.86, 95% CI 0.42 to 1.31, I ^2^ = 68.27%) with significant heterogeneity. The results showed that the Shannon and Chao index were statistically significant in the vagina (Figure 1C,D). Subgroup analysis was performed according to the country, and no statistical difference between patients infected with HPV and the healthy controls was observed (Shannon index: Hedges' g = 1.15, 95% CI −0.36 to 2.65, I ^2^ = 98.09%) (Figure 2A). Subgroup analysis was performed based on the sequencing platform (Figure 2B) and primer pair (variable region) selection (Figure 2C). When the primer pair was selected on V3–V4 the statistical difference (Shannon index: Hedges' g = 0.50, 95% CI 0.28 to 0.72, I ^2^ = 0.00%) was observed. This suggests that the site of sample collection and the choice of the primer pair may be responsible for heterogeneity.
Subgroup analysis of Shannon index between patients with HPV infection and healthy control: (A) China, (B) Illumina MiSeq, (C) primer pairs.
The microbial diversity between patients with HPV infection and CC was statistically insignificant (Shannon index: Hedges' g = −5.09, 95% CI −13.24 to 3.05, I ^2^ = 98.91%, Chao index: Hedges' g = −2.15, 95% CI −5.40 to 1.11, I ^2^ = 97.53%) (Figure 3A,B). Chao index (Hedges' g = 0.63, 95% CI 0.20 to 1.47, I ^2^ = 87.57%) and Shannon index (Hedges' g = 0.30, 95% CI 0.02 to 0.58, I ^2^ = 0.00%) revealed significant differences in microbial diversity between patients with low‐grade cervical lesions and healthy controls (Figure 3C,D). These results suggest that cervical lesions can disrupt the structure and function of the residing microbiome.
Forest plot of the differences in alpha diversity between patients with HPV infection and cervical cancer: (A) Shannon index, (B) CHAO index; alpha diversity between patients with low‐grade squamous intraepithelial lesion and healthy control: (C) Shannon index, (D) CHAO index.
Beta Diversity
3.7
Eight studies reported differences between the microorganisms in patients with HPV and HrHPV infections and healthy controls [8, 21, 28, 32, 48, 55, 56, 60], and a considerable number of reports also showed no difference between the two groups [13, 16, 25, 30, 33, 45, 54, 64, 65]. Two studies reported differences in beta diversity between patients with persistent and transient HPV infections [15, 66], while two showed no difference [48, 52]. The cervicovaginal microbiome exhibits differences between single HPV and multiple HPV infections [29, 54, 59, 60].
The beta diversity of patients with CC was significantly different from that of healthy controls [6, 20, 21, 22, 23, 28, 31, 34, 37, 43, 44, 46, 67], similar to the differences observed in alpha diversity; only three studies reported no difference [42, 45, 66]. The diversity of microorganisms undergoes dynamic changes during the development of CC [4, 6, 8, 18, 19, 20, 21, 24, 26, 27, 35, 37, 39, 43, 44, 46]. However, some studies have pointed out that there is not necessarily a strong causal relationship between cervicovaginal microbial diversity and the severity of cervical lesions [11, 23, 24, 25, 38, 45, 68, 69]. Overall, the non‐overlapping results indicate the need for studies with more homogeneous and larger cohort sizes in order to infer a statistically significant correlation of microbial features (like diversity) with disease phenotypes.
Bacterial Groups Enriched in Diseased Condition
3.8
Table S3 shows the enrichment of different bacteria in diseased conditions, and Figure 4 shows the number of studies that reported an enrichment of a bacteria in diseased conditions at the genus level.
Number of times that different bacteria have enriched for different diseases at the genus level. CC, cervical cancer; CIN1, low‐grade cervical intraepithelial neoplasia; CIN23, high‐grade cervical intraepithelial neoplasia; HPV, human papillomavirus infection; HrHPV, high‐risk human papillomavirus infection; HSIL, high‐grade squamous intraepithelial lesion; LSIL, low‐grade squamous intraepithelial lesion; N, healthy control; PH, persistent HPV infections; TH, transient HPV infections.*
Patients with HPV and HrHPV infections are enriched with similar bacterial species (Figure 4A,B), including Gardnerella [30, 33, 56, 61, 64, 70], Sneathia [30, 49, 62, 71], Atopobium [30, 56, 61, 62, 70], Prevotella [25, 55, 62], Veillonella [18, 70], Peptostreptococcus [33, 62, 69], Megasphaera [35, 63], Acinetobacter [72], Bifidobacterium [61], Fusobacterium [51, 62, 71], Ureaplasma [51], Dialister [70], and Pseudomonas [30, 49]. Gardnerella, as reported by the highest number of studies to be enriched in HPV infection, can be investigated further regarding its role in the etiology of HPV infection and its potential as a biomarker for improved screening and diagnosis.
For patients with CC, the biomarkers in different studies did not completely overlap (Figure 4C), and most pathogenic and enriched bacteria included Porphyromonas [6, 18, 35], Fusobacterium and Prevotella [6, 17, 18, 40, 42, 66], Mycoplasma [6], Megasphaera [6], Campylobacter, Dialister [6, 32, 40, 66], Peptoniphilus, Peptostreptococcus [40], Anaerococcus [6], WAL_1855D [6], Bacillus, Sneathia [17, 35, 40], Bacteroides [18, 40], Veillonella [40, 42], Saccharimonas, Streptococcus, Ureaplasma [42], Gardnerella [32], and Atopobium [32]. Compared with the healthy controls, the proportions of Prevotella, Fusobacterium, and Dialister increased with the progression of cervical lesions [18, 73], whereas Gardnerella was less prominent in CC. Porphyromonas [6, 18, 31, 35, 71] can be used as a marker for screening CC compared with different degrees of cervical lesions (Figure 4D,E). Lactobacillus was dominant during transient HPV infection (Figure 4F) and had an unstable microbiome environment [9, 46, 48, 69]. Prevotella [10, 49, 50, 51], Dialister [48, 51], Atopobium [15, 49], Gardnerella, and Anaerococcus [48, 50] were the most common bacteria in patients with persistent HPV infection. These bacteria may play a role in disrupting the epithelial barrier [10], thus favoring persistent HPV infection or transmission. Different types of Lactobacilli in the cervicovaginal microbiota show significant differences in their correlation with HPV [18, 50, 51, 54].
Both persistent and transient infections are associated with subgroups of Lactobacillus, and the function of different types of Lactobacilli is usually considered a protective mechanism. However, subgroups of Lactobacillus have been reported to be closely associated with increased vaginal inflammation and an increased probability of transformation into bacterial vaginosis. Therefore, different types of Lactobacilli, such as Lactobacillus iners , L. crispatus , and L. gasseri , have bidirectional roles in cervicovaginal health and disease [18, 74].
Risk of Bias Assessment
3.9
The quality assessment results of these studies are presented in Table S2. Among 10 cohort studies, three scored 9 points, one scored 8 points, and the others were of moderate quality: four scored 7 points, and two scored 6 points. In some studies, the follow‐up time was short, or the number of follow‐ups was missing. Among the identified 54 case–control studies, 15 studies were of high quality: three scored 9 points, 12 studies scored 8 points, and 30 studies were of moderate quality: 12 scored 7 points, and 18 scored 6 points; the other studies scored 5 and 4 points. “Comparability of Cases and Controls on the Basis of the Design or Analysis” and “Non‐Response rate” were not identified in most studies.
Discussion
4
This systematic review and meta‐analysis evaluated studies on microbiota changes in patients infected with HPV and with CC using high‐throughput sequencing. The normal cervicovaginal microbiota structure is simple, less diverse, and dominated by Lactobacillus. Microbial dysbiosis and increased diversity are more likely to allow HPV invasion, which is a risk factor for CC and is related to the severity of cervical lesions. The differences in country/population, age, microbial sequencing technology, primer pair, and sample collection site (vaginal and cervical secretions) significantly impacted the composition of microorganisms.
The composition of the cervicovaginal microbiota is dynamic; it varies depending on the female reproductive cycle, hormonal changes, oral contraceptives, sexual activity, lactation, and smoking [7] and exhibits highly variable associations with race, geography, and individual genetic factors [13, 65]. Studies on cervicovaginal microecology have focused only on the disease and neglected the impact of external factors. However, environmental and physiological changes can greatly affect patients infected with HPV. While this study describes changes in the vaginal microbiota following HPV infection in different ethnic groups, there is limited research comparing the microbiota of patients from different backgrounds within the same study to allow for direct comparison and control of confounding factors. Future studies should compare racial and ethnic groups to better characterize the influence of race/ethnicity on the cervicovaginal microbiota.
HPV infection causes extreme changes in the composition and abundance of bacteria within a short interval [53], constantly shifting and evolving bacterial communities over time and possibly eventually returning to their original state [75]. However, the duration of the process remains unclear. A positive association was found between vaginal pH and HPV positivity in women (18–34 years), but the differences were not statistically significant in women aged > 65 years [76]. Changes in vaginal pH are associated with the vaginal microenvironment; therefore, age is also a factor that cannot be ignored.
Subgroup analysis revealed that the sampling location was an important contributing factor. Although the anatomical definition, the low female reproductive tract (vagina and cervix) is closely linked [77], most studies prefer to refer to samples as “cervicovaginal” rather than discuss “cervical” and “vaginal” samples. However, different parts of the female genital tract have versatile microbiota [47]. The sampling location and sampling method are important in determining the individual vaginal microbial community composition [78]. Subsequent studies should be described separately as much as possible, or standardized sampling locations and methods should ensure the homogeneity of the research results. Further studies are needed to determine which site plays a greater role in the disease.
Differences in existing sequencing methods are also an influencing factor, as the selection of primer pairs (variable regions) is more important than the sequencing platform. When the primer pair V3–V4 (341F–785R) is selected, the bacteria coverage is the highest. A study has also shown that the primer pair V3–V4 slightly outperformed the other combinations, regardless of the reference database, and is a justified choice for human gut samples [12]. With the development of metagenomic and V1–V9 full‐length sequencing, microbiome information can be mined in more detail at very high sequencing depths. However, owing to the high economic cost of such sequencing methods, the number of studies is limited, and their advantages are not clear.
Only seven studies were included in the quantitative analysis; the statistical power of our meta‐analysis was also reduced by the large amount of missing data for published studies. Six studies were from China, which had great heterogeneity and could not represent global data.
The biomarkers at different stages of cervical lesions also differed, which may be related to the different reference controls. The relative abundance of bacterial genera identified in the same group under different primer pairs was different, leading to significant changes in biomarkers identified by the LefSe statistical tool [79]. Gardnerella was more likely to appear during HPV infection [37, 56, 61, 70], and Prevotella, Dialister, and Porphyromonas were closely related to advanced cancer. Biogenic Amines [80] and metabolites of Prevotella, Dialister, and Peptostreptococcus can protect bacteria from acidic conditions while also reducing the growth rate of Lactobacillus and promoting the establishment and maintenance of the Community State Type IV (CST IV, Low‐Lactobacillus) microbiota, which is not found in Gardnerella. In addition, these bacteria mediate carcinogenesis, including generating pro‐inflammatory conditions, immune suppression, and suppression of apoptosis [6].
Limitations and Future Suggestions
4.1
The meta‐analysis performed in the current review was limited by data heterogeneity, which included patients with HPV infection and CC from different countries and diverse data generation and analysis approaches. The number of studies included in this meta‐analysis was limited. In the future, with an increase in the study population and the deepening of the research direction, more high‐quality studies with large sample sizes should be included to draw more reliable and convincing conclusions. All of the included primary studies were observational studies.
Despite the limitations, the HPV vaccine is the best way to prevent CC. One study has shown that the diversity and dominant organisms of the vaginal microbiome do not change after women receive three doses of the HPV‐16/18 AS04‐adjuvanted vaccine (Cervarix) [81]. However, due to the small sample size of this study, it cannot be used as sufficient evidence to prove the impact of vaccines on microorganisms. Current approaches to regulating the microbiome for potentially positive health outcomes include therapeutic HPV vaccines based on probiotic engineering [82]. Mucosal vaccines can directly target the mucosa in the reproductive tract to deliver antigens and induce a potent local protective immune response [83]. It has been shown that mucosal vaccines based on Lactobacillus lactis transfected with HPV E6/E7 hold promise in preventing HPV infection and halting the progression of CC [82, 84]. Decreases in vaginal microbiota by antibiotics, replenishment of the microbiota by vaginal microbiota transplantation (VMT), and supplementation of the microbiota with prebiotics or probiotics may impact response to cancer treatment [77].
Conclusions
5
The microbial diversity in patients with HPV infection is highly unstable, and the impact of confounding factors on their study results must be considered. The microecological environment of the cervix and vagina changes continuously after cervical lesions occur. As long as cervical epithelial lesions develop, the microbial diversity increases, and the complexity of the microecological environment intensifies with the progression of cervical lesions. Distinct bacterial markers were identified at different stages of the cervical lesions. Gardnerella was prevalent in the stage of HPV infection; however, its proportion decreased significantly after the occurrence of CC. The proportions of Prevotella, Bacillus, and Porphyromonas were also found to be increased. The lack of high‐quality randomized controlled trial data makes it difficult to provide strong evidence of a causal relationship between microbial HPV infection and cervical lesions, and its mechanism has not been deeply explored. Therefore, additional randomized controlled trials are needed to confirm the relationship between the cervical vaginal microenvironment and the progression of cervical lesions.
Author Contributions
Conceptualization, methodology, formal analysis, investigation, data curation, writing‐original draft preparation, and project administration: W.Z. Methodology, investigation, writing‐review and editing: Y.G. Formal analysis, writing‐original draft preparation: L.Y. Investigation, writing‐review and editing: Q.Y. Methodology, investigation, writing‐review and editing: J.W. Conceptualization, investigation, writing‐review and editing, and supervision: Y.Y. Conceptualization, methodology, investigation and supervision: B.L. All authors have read and agreed to the published version of the manuscript.
Ethics Statement
The authors have nothing to report.
Consent
The authors have nothing to report.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Data S1. Checklist S1: PRISMA 2020 Checklist.
Figure S1. Description of the selection of the included studies following a PRISMA flow diagram.
Table S1. Search strategies.
Table S2. Quality Assessment.
Table S3. Bacterial groups enriched in diseased condition.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1M. Arbyn , E. Weiderpass , L. Bruni , et al., “Estimates of Incidence and Mortality of Cervical Cancer in 2018: A Worldwide Analysis,” Lancet Global Health 8, no. 2 (2020): e 191–e 203, 10.1016/S 2214-109X(19)30482-6.31812369 PMC 7025157 · doi ↗ · pubmed ↗
- 2Y. Tian , S. Zhang , and F. Ni , “Targeting Glucose Metabolism for HPV‐Associated Cervical Cancer: A Sweet Poison,” Biomedicine & Pharmacotherapy 180 (2024): 117519, 10.1016/j.biopha.2024.117519.39378679 · doi ↗ · pubmed ↗
- 3M. Berggrund , I. Gustavsson , R. Aarnio , et al., “Temporal Changes in the Vaginal Microbiota in Self‐Samples and Its Association With Persistent HPV 16 Infection and CIN 2,” Virology Journal 17, no. 1 (2020): 147, 10.1186/s 12985-020-01420-z.33028395 PMC 7541248 · doi ↗ · pubmed ↗
- 4X. Huang , C. Li , F. Li , J. Zhao , X. Wan , and K. Wang , “Cervicovaginal Microbiota Composition Correlates With the Acquisition of High‐Risk Human Papillomavirus Types,” International Journal of Cancer 143, no. 3 (2018): 621–634, 10.1002/ijc.31342.29479697 · doi ↗ · pubmed ↗
- 5K. S. Vikramdeo , S. Anand , J. Y. Pierce , A. P. Singh , S. Singh , and S. Dasgupta , “Distribution of Microbiota in Cervical Preneoplasia of Racially Disparate Populations,” BMC Cancer 22, no. 1 (2022): 1074, 10.1186/s 12885-022-10112-6.36258167 PMC 9578267 · doi ↗ · pubmed ↗
- 6S. Wu , X. Ding , Y. Kong , et al., “The Feature of Cervical Microbiota Associated With the Progression of Cervical Cancer Among Reproductive Females,” Gynecologic Oncology 163, no. 2 (2021): 348–357, 10.1016/j.ygyno.2021.08.016.34503848 · doi ↗ · pubmed ↗
- 7G. Curty , P. S. de Carvalho , and M. A. Soares , “The Role of the Cervicovaginal Microbiome on the Genesis and as a Biomarker of Premalignant Cervical Intraepithelial Neoplasia and Invasive Cervical Cancer,” International Journal of Molecular Sciences 21, no. 1 (2019): 222, 10.3390/ijms 21010222.31905652 PMC 6981542 · doi ↗ · pubmed ↗
- 8M. E. Nieves‐Ramirez , O. Partida‐Rodriguez , P. Moran , et al., “Cervical Squamous Intraepithelial Lesions Are Associated With Differences in the Vaginal Microbiota of Mexican Women,” Microbiology Spectrum 9, no. 2 (2021): e 0014321, 10.1128/Spectrum.00143-21.34643408 PMC 8515943 · doi ↗ · pubmed ↗