In vitro screening of dihalomethanes as potential methane inhibitors in dairy cows
M. Thorsteinsson, M.O. Nielsen

TL;DR
This study tested dihalomethanes as methane inhibitors in dairy cows and found that some significantly reduced methane production but also affected rumen fermentation.
Contribution
The study identifies specific dihalomethanes with high methane-mitigating potential while highlighting their impact on rumen fermentation.
Findings
Dibromomethane, bromoiodomethane, and diiodomethane reduced methane production by over 90%.
Dichloromethane had no significant methane-mitigating effect.
Methane reduction was accompanied by reduced volatile fatty acid production.
Abstract
Summary: Halogenated compounds, particularly trihalomethanes, have demonstrated large potential as antimethanogenic feed additives. However, concerns can be raised in relation to the use of these compounds as feed additives due to the transfer of halogenated metabolites into milk. This in vitro study aimed to identify dihalomethanes with CH4-mitigating potential to potentially reduce the transfer of halogens to milk when used as feed additives. When added to corn silage, CH4 production was reduced by the dihalomethanes dibromomethane (>90%), bromoiodomethane (>90%), and chloroiodomethane (58%) compared with corn silage incubated alone in a buffered rumen inoculum, whereas dichloromethane showed no CH4-mitigating effect. However, reductions in CH4 production were accompanied by reductions in total volatile fatty acid (VFA) production; hence, future studies should focus on finding the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
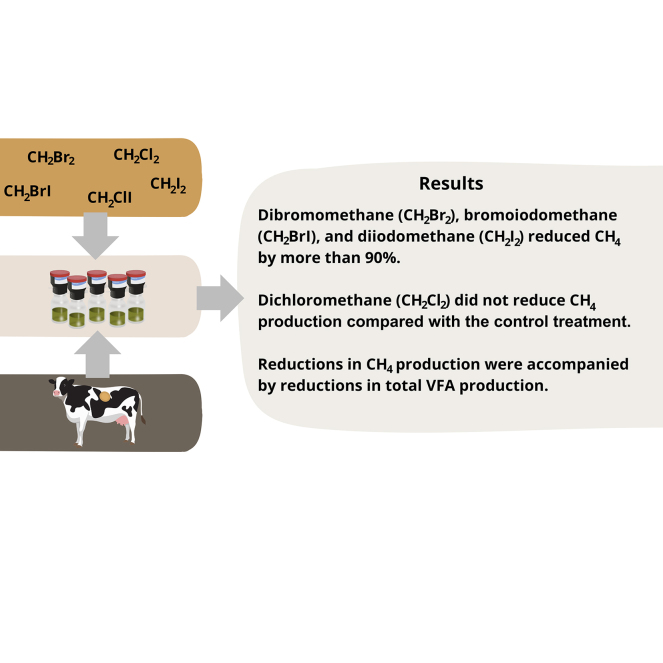 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRuminant Nutrition and Digestive Physiology · Alcohol Consumption and Health Effects · Odor and Emission Control Technologies
Research investigating the effects of volatile halogenated compounds (VHC) as ruminal CH_4_ inhibitors has been conducted for several decades (Czerkawski and Breckenridge, 1975; Thorsteinsson et al., 2023a). The primary mode of action is believed to be the competitive binding of the compounds to enzymes in the final steps of methanogenesis (Wood et al., 1968), but VHC have also been suggested to inhibit CH_4_ production by serving as competing terminal electron acceptors (Yu and Smith, 2000). The antimethanogenic activity is usually related to the number of halogens on the molecule with iodine-containing compounds being the most efficient followed by brominated and chlorinated analogs (Chalupa, 1980). Hence, 30% to 66% reductions in enteric CH_4_ yield (g/kg DMI) have been reported when bromoform (extract from the red seaweed Asparagopsis taxiformis), chloroform, and iodoform were administered to dairy cows as antimethanogenic feed additives (Martinez-Fernandez et al., 2018; Thorsteinsson et al., 2023a; Alvarez-Hess et al., 2024). However, the use of VHC as antimethanogenic feed additives is not unproblematic as bromoform and chloroform can be categorized as potential human carcinogens (EPA, 2000; ECHA, 2024). Moreover, daily levels of supplemented iodine exceeded the maximum total limit set by the US Food and Drug Administration regulations (NRC, 2021) when iodoform was used as a CH_4_ inhibitor for dairy cows (Thorsteinsson et al., 2023a). Hence, concerns can be raised regarding the transfer of halogenated metabolites into milk, which may pose a threat toward consumer health (Krizsan et al., 2023). The aforementioned VHC are all trihalomethanes, indicating that the molecule consists of 3 halogens and 1 hydrogen covalently bound to 1 carbon atom. Transfer of halogens originating from halogenated CH_4_-mitigating feed additives into milk can, at least in theory, be reduced by reducing the number of halogen atoms on the molecule.
Therefore, the objective of the current study was to investigate the potential of 5 different dihalomethanes (DHM) as antimethanogenic feed additives in vitro. It was hypothesized that the DHM dibromomethane (CH_2_Br_2_; BM), bromoiodomethane (CH_2_BrI; BIM), dichloromethane (CH_2_Cl_2_; CM), chloroiodomethane (CH_2_ClI; CIM), and diiodomethane (CH_2_I_2_; DIIM) all would reduce CH_4_ production from a standard feed incubated in vitro in buffered rumen fluid, with the iodine-containing compounds being most efficient.
To compare the antimethanogenic potential of the 5 DHM (BM, BIM, CM, CIM, and DIIM), the compounds were evaluated on a molar basis. Hence, 2 mM solutions were prepared by dissolving the compounds in 99% ethanol. Due to the differences in molar masses of the compounds, this resulted in different wt/vol concentrations in the solutions (Table 1).Table 1. Information on chemicals used in treatmentsItemTreatment1BIMBMCIMCMDIIMCAS no.557–68–6274–95–32593–71–5275–09–2275–11–62Concentration (mg/mL)0.440.350.350.170.541BM = dibromomethane; BIM = bromoiodomethane; CM = dichloromethane; CIM = chloroiodomethane; DIIM = diiodomethane.2Purchased from Merck, Darmstadt, Germany.
In 2 separate runs, 0.1 mL of the solutions were added to Duran bottles (DWK Life Sciences, Mainz, Germany; capacity: 132.0 ± 1.1 mL) together with 0.5 ± 0.02 g of a standard feed, corn silage (ash: 3.54%; CP: 7.77%; NDF: 32.9%; starch: 35.1% on a DM basis). To quantify the CH_4_-mitigating potential of the treatments, corn silage (0.5 ± 0.02 g) with the addition of 0.1 mL of 99% ethanol was included as control samples, whereas buffered rumen fluid incubated alone with 0.1 mL of 99% ethanol served as blank samples. Each sample type was included as triplicates in both runs. To minimize possible confounding effects of bottle position and inoculation, the runs were divided into 3 blocks with one bottle per treatment in each block. The treatments were randomly assigned to a bottle within block.
Three rumen-cannulated nonlactating Danish Holstein cows (BW 755.6 ± 105.4 kg and BCS 3.75 ± 0.43) were used for collection of rumen fluid. The handling and care of the cows complied with the guidelines set out by the Danish Ministry of Environment and Food (Act No. 2028, 2020; Danish Ministry of Environment and Food, 2000) concerning animal experimentation and care of animals under experiments. The cows were fed 2 equal-sized meals of a diet consisting of grass-clover hay, barley straw, and a pelleted concentrate mixture (39.6% barley grain, 39.9% oat grain, 10% soybean meal, 3% rapeseed meal, 3% sugar beet molasses, and 4% of a commercial mineral mixture per kg fresh mixture) at maintenance level, which amounted to 7.7 kg of DM daily (with 13.7% starch, 13.9% CP, 24.1% NDF, and forage to concentrate ratio of 69:31 on a DM basis). Approximately 30 min before morning feeding, rumen liquid and particles were sampled and filtered through a nylon cloth with a pore size of 250 µm into preheated vacuum flasks. Buffer, redox indicator, reducing agent, and macro- and micromineral solutions were prepared and mixed as described by Menke and Steingass (1988). Rumen fluid was added in a 2:1 ratio (buffer solution: rumen fluid). After mixing, the solution was kept under anaerobic conditions by continuously flushing with N_2_. The final pH of buffered rumen fluid was 7.92 ± 0.10. Ninety milliliters of inoculum and 0.1 mL of additives were added to each bottle. Thereafter, a wireless fully automated ANKOM^RF^ module (ANKOM Technology, Macedon, NY) was mounted on each bottle for measurement of in vitro gas production, and the headspace was flushed with N_2_. The ANKOM^RF^ was operated as described by Thorsteinsson et al. (2023b). A gas-tight FlexFoil bag (1 L; SKC Ltd., Dorset, United Kingdom) was attached to each sensor module for the continuous collection of released gas. The bottles were incubated for 48 h at 38.5°C in a controlled incubator shaker (New Brunswick Excella E25R, Eppendorf, Hamburg, Germany) with the oscillation set at 50 rpm. After 24 h of incubation, 10 mL of gas was extracted from each gasbag using a gas-tight syringe with a twist valve (Hamilton Bonaduz AG, Bonaduz, Switzerland). Then the gas was transferred into Exetainer vials (Labco Limited, Ceredigion, United Kingdom) for later gas analyses. After 48 h of incubation, a sample of the fermented inoculum was collected for later VFA analysis by filtration of bottle contents through a F57 ANKOM bag (ANKOM Technology, Macedon, NY).
The corn silage was freeze-dried and ground on a 2-mm screen (Ultra Centrifugal Mill ZM 200, Verder Scientific, Hann, Germany). Nutrient content was analyzed using near-infrared spectroscopy at a commercial feed testing laboratory (Eurofins Agro Testing A/S, Vejen, Denmark). The collected inoculum was stabilized with 1 mL of 25% metaphosphoric acid (MPA) to reach 5% MPA in the stabilized sample. The concentrations of VFA were determined in stabilized inoculum after methanol-chloroform extraction with 2-ethylbutyrate as the internal standard, using GC (Trace 1310, Thermo Scientific, Germany) with a split/splitless injector at 225°C and a flame ionization detector at 250°C. A 30 m × 0.53 mm × 1 µm HP-FFAP column (Agilent Technologies Inc., Wilmington, DE) was used with helium as carrier gas at 34.5 kPa.
Concentrations of CH_4_ and H_2_ were determined in gas samples using GC (Trace 1310, Thermo Scientific, Germany) with a split/splitless injector at 150°C and a thermal conductivity detector at 200°C. A 30 m × 0.25 mm × 50 µm Rt-Q-BOND column (MicroQuartz, Munich, Germany) was used with argon as the carrier gas at 4 mL/min flow. The gas chromatograph was programmed with a gradually increased temperature starting at 80°C for 7 min and then from 80°C to 150°C at 20°C/min. Quantification was performed using standard curves with standard gases in concentrations ranging from 500 to 250,000 ppm for CH_4_ and from 250 to 50,000 ppm for H_2_.
Cumulative gas pressure was converted from pounds per square inch (psi) to milliliters (mL) of gas produced at standard temperature and pressure (IUPAC, 2014), assuming that the ideal gas law applies to the produced gases (predominantly CO_2_, CH_4_, and H_2_). From the cumulated total gas production (mL) and CH_4_ concentration (%) in collected gas after 24 h incubation, CH_4_ production (mL) was calculated. Total VFA production (mmol) after 48 h was calculated from concentration (mmol/L) and total inoculum volume in the bottles (L). Before the statistical analysis, total gas production parameters were blank-corrected.
The various response parameters were analyzed with the following linear mixed model in R 4.4.1 (R Core Team, 2024):
where Y_te_ is the dependent response variable, μ is the overall mean, α is the fixed effect of treatment (t = 1–5), A is the random effect of experimental run (e = 1–2), and ε_te_ is the random residual error, assumed to be independent with constant variance and normally distributed. Data are presented in tables as estimated marginal means (EMS) and SEM. Differences between EMS were evaluated using Tukey's method for comparison. Statistical significance was declared when P ≤ 0.05 and statistical tendency was declared when 0.05 < P ≤ 0.10.
Chalupa (1980) claimed that chlorinated VHC were less effective as CH_4_ inhibitors compared with iodinated and brominated VHC. This hypothesis seems to be supported by the results from the current study, at least when evaluated on a molar basis, as all DHM, except CM, significantly (P < 0.001) reduced CH_4_ per gram of incubated DM and OM compared with the control (Table 2). Moreover, BIM and DIIM resulted in significantly stronger suppression of CH_4_ production compared with CIM. These reductions were caused by a combination of both reduced total gas production and substantially lower CH_4_-percentage. However, it should be noted that wt/vol concentrations of the DHM in the solutions varied due to differences in molecular weights, causing the wt/vol concentration of DIIM to be 3 times as high as CM. In addition to the type of halogen on the molecules and concentration, the volatility of the compounds might also have affected the antimethanogenic potential of the DHM. The CM has a low boiling point (40°C), close to that of the incubation temperature, whereas the remaining compounds had boiling points ranging from 67 (DIIM) to 138°C (BIM; Merck, 2024). Hence, CM might have evaporated from the inoculum into the headspace relatively quickly after the initiation of the experiment and released from the bottles during the venting of the system. This hypothesis is supported by the similar CH_4_ between control and CM.Table 2. Methane and hydrogen production after 24 h of incubationItemTreatment1SEMP-valueBIMBMCIMCMDIIMControlTotal gas production after 24 h mL/g DM in corn silage74.9b70.8b76.3ab90.3ab71.8b98.1a6.88<0.01 mL/g OM in corn silage77.1b72.9b78.6ab93.0ab73.9b101.0a7.09<0.01Methane after 24 h mL/g DM in corn silage0.150c0.929bc4.63b9.88a0.463c11.1a0.973<0.001 mL/g OM in corn silage0.154c0.957bc4.76b10.2a0.446c11.4a1.00<0.001 Concentration in total gas (%)0.200c1.34c6.05b10.9a0.586c11.1a0.517<0.001Hydrogen after 24 h mL/g DM in corn silage3.79a2.92a0.102b0.025b3.35a0.022b0.620<0.001 mL/g OM in corn silage3.90a3.01a0.105b0.025b3.45a0.023b0.638<0.001 Concentration in total gas (%)5.08a4.68a3.99a0.127b4.68a0.022b0.806<0.001a–cValues within the same row with different superscripts differ (P < 0.05).1BM = dibromomethane; BIM = bromoiodomethane; CM = dichloromethane; CIM = chloroiodomethane; DIIM = diiodomethane.
The H_2_ production followed the opposite pattern than CH_4_ with the highest production on BIM, BM, and DIIM. This change in gas composition was reflected in the VFA profile as the proportion of propionate (P < 0.001), formed in a [H]-consuming pathway, in total VFA increased, whereas the proportion of acetate (P < 0.001), formed in a net [H]-releasing pathway, in total VFA decreased causing the acetate:propionate ratio to decrease with increasing inhibition of CH_4_ production (P < 0.001; Table 3; Guyader et al., 2017). Valerate and caproate are normally considered to be [H]-sinks (Ungerfeld, 2015). Interestingly, while the proportion of propionate and butyrate appeared to follow the expected pattern toward a lower net [H]-release during reduced methanogenesis (Ungerfeld, 2020), the control treatment and CM had the highest proportion of caproate and valerate (P < 0.001 for both parameters). An increased ruminal H_2_ pressure has been speculated to change the ruminal thermodynamics and thereby negatively affect fermentation (Janssen, 2010; van Lingen et al., 2016). In the current study, the in vitro system was continuously vented during the incubation at a lower partial pressure than observed in the rumen (van Lingen et al., 2017). Hence, H_2_ accumulation cannot explain significantly lower total VFA production on all DHM treatments except CM. Therefore, it is possible that too high doses of DHM may have a negative impact on other ruminal microbes than the methanogenic archaea. A similar negative effect on ruminal digestibility was observed in vivo by Thorsteinsson et al. (2023a) in dairy cows administered increasing doses of iodoform intraruminally. Moreover, although microbial protein synthesis was not determined in the current in vitro study, the significantly (P < 0.001) lower proportion of the iso-fatty acid isovalerate could indicate a reduced efficiency of microbial protein synthesis.Table 3. Volatile fatty acid production after 48 h of incubationItemTreatment1SEMP-valueBIMBMCIMCMDIIMControlTotal VFA production after 48 h (mmol)6.91c7.24b7.23b7.74a6.92c7.97a0.111<0.001% of total VFA Acetate56.1d58.9c61.8b68.0a56.4d68.3a0.809<0.001 Propionate22.4a20.6b18.2c12.9d22.1ab12.5d0.430<0.001 Butyrate16.7a15.8ab14.6b10.1c16.7a9.56c0.477<0.001 Valerate1.55b1.43b1.47b1.96a1.54b2.02a0.0680<0.001 Caproate0.232b0.266b0.490b3.35a0.249b3.67a0.212<0.001 Isobutyrate1.06a1.09b1.09b1.09b1.10b1.08ab0.0243<0.01 Isovalerate1.91b1.93b2.30ab2.64a1.94b2.80a0.215<0.001Acetate:propionate ratio2.51d2.87c3.41b5.29a2.56cd5.45a0.121<0.001a–dValues within the same row with different superscripts differ (P < 0.05).1BM = dibromomethane; BIM = bromoiodomethane; CM = dichloromethane; CIM = chloroiodomethane; DIIM = diiodomethane.
In conclusion, all DHM reduced CH_4_ production in vitro, except CM, with BIM, BM, and DIIM resulting in the largest reductions (>90%). The opposite pattern was observed for H_2_ production. Depression of CH_4_ production was associated with a negative effect on total VFA production. Hence, future studies should focus on finding the optimal dose of the DHM resulting in the largest CH_4_ reduction without negative effects on rumen fermentation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alvarez-Hess P.S.Jacobs J.L.Kinley R.D.Roque B.M.Neachtain A.S.O.Chandra S.Russo V.M.Williams S.R.O.Effects of a range of effective inclusion levels of Asparagopsis armata steeped in oil on enteric methane emissions of dairy cows Anim. Feed Sci. Technol.3102024115932 10.1016/j.anifeedsci.2024.115932 · doi ↗
- 2Chalupa W.Chemical control of rumen microbial metabolism Ruckebusch Y.Thivend P.Digestive Physiology and Metabolism in Ruminants: Proceedings of the 5th International Symposium on Ruminant Physiology 1980 Springer Dordrecht, the Netherlands 32534710.1007/978-94-011-8067-2_16
- 3Czerkawski J.W.Breckenridge G.New inhibitors of methane production by rumen micro-organisms. Development and testing of inhibitors in vitro.Br. J. Nutr.34197542944610.1017/S 000711457500049912012671201267 · doi ↗ · pubmed ↗
- 4Danish Ministry of Environment and Food Bekendtg⊘relse om dyrefors⊘g Act no. 2028)https://dyreforsoegstilsynet.dk/om/lovgivning 2000
- 5ECHA Information on Chemicalshttps://echa.europa.eu/substance-information/-/substanceinfo/100.000.7772024
- 6EPA Bromoform; CASRN 75–25–2https://www.epa.gov/sites/default/files/2016-09/documents/bromoform.pdf 2000
- 7Guyader J.Ungerfeld E.M.Beauchemin K.A.Redirection of metabolic hydrogen by inhibiting methanogenesis in the rumen simulation technique (RUSITEC)Front. Microbiol.8201739310.3389/fmicb.2017.003932835225628352256 PMC 5349286 · doi ↗ · pubmed ↗
- 8IUPAC Compendium of Chemical Terminology. 2nd ed. Version 2.3.3https://goldbook.iupac.org/files/pdf/goldbook.pdf 2014