Molecular analysis of blood-associated pathogens in common ravens (Corvus corax) from Germany
Aline Lamien-Meda, Josef Harl, Astrid Lieber, Maria Sophia Unterköfler, Barbara Eigner, Licha N. Wortha, Franz Müller, Mike Heddergott, Hans-Peter Fuehrer

TL;DR
This study found that a significant portion of common ravens in Germany are infected with various blood parasites, highlighting the need for further research on these infections.
Contribution
The study provides new insights into the prevalence and diversity of blood parasites in common ravens in Germany using molecular techniques.
Findings
26.2% of common ravens were PCR-positive for at least one blood parasite.
Filarioid nematodes showed the highest prevalence at 16.7%.
Four Leucocytozoon CytB lineages and one Haemoproteus lineage were identified.
Abstract
Common ravens (Corvus corax) are intelligent scavengers that adapt to diverse environments, playing a key ecological role, but their health and ecosystem contributions can be affected by parasites. This study investigates the prevalence and diversity of blood parasite infections in common ravens using molecular techniques. Blood samples (n = 42) were collected from dead common ravens in Germany and screened for filarioid nematodes, trypanosomatids, and haemosporidian parasites. The results showed that 26.2% of the common ravens were PCR-positive for at least one parasite, with some cases of mixed infections. Filarioid nematodes were found in 16.7%, trypanosomatids in 4.8%, and haemosporidian parasites in 16.7% of the common ravens. Sequencing revealed the presence of four Leucocytozoon CytB lineages and one Haemoproteus lineage. The findings suggest that common ravens in Germany are…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
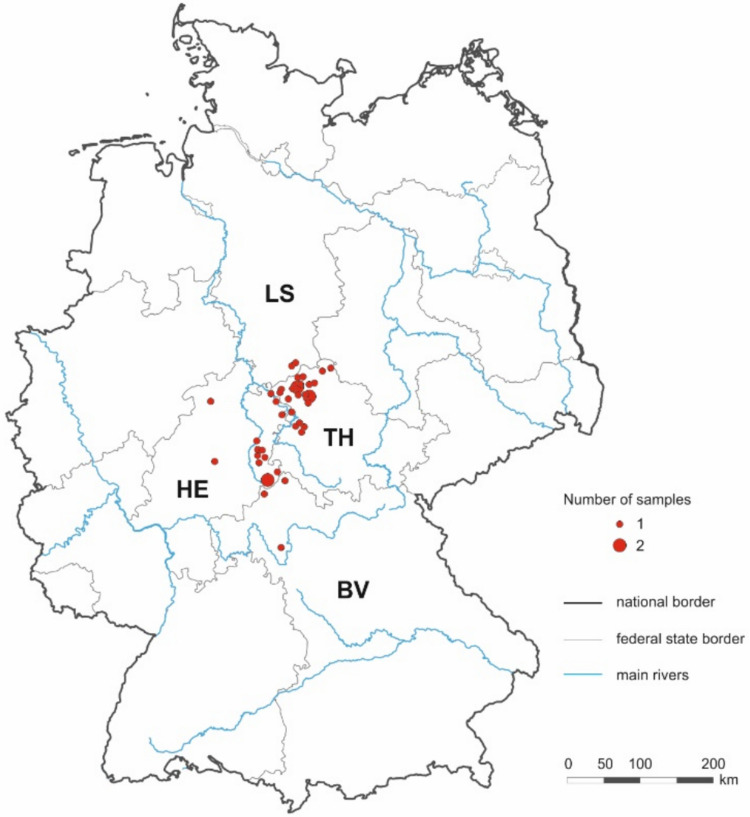 Figure 1
Figure 1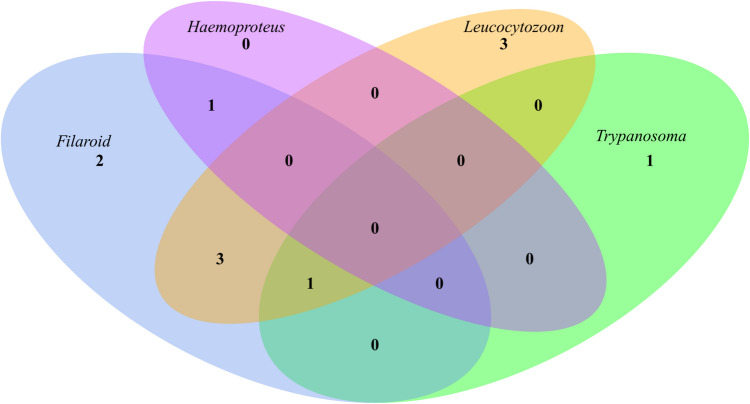 Figure 2
Figure 2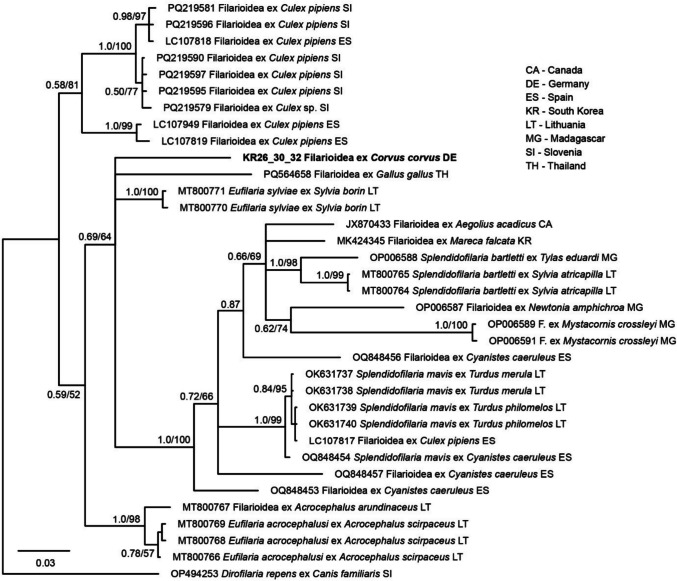 Figure 3
Figure 3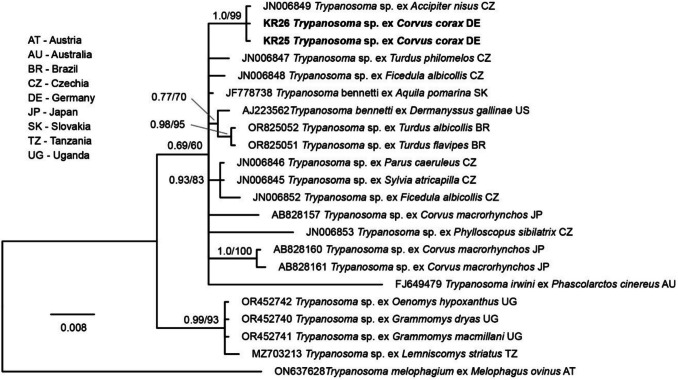 Figure 4
Figure 4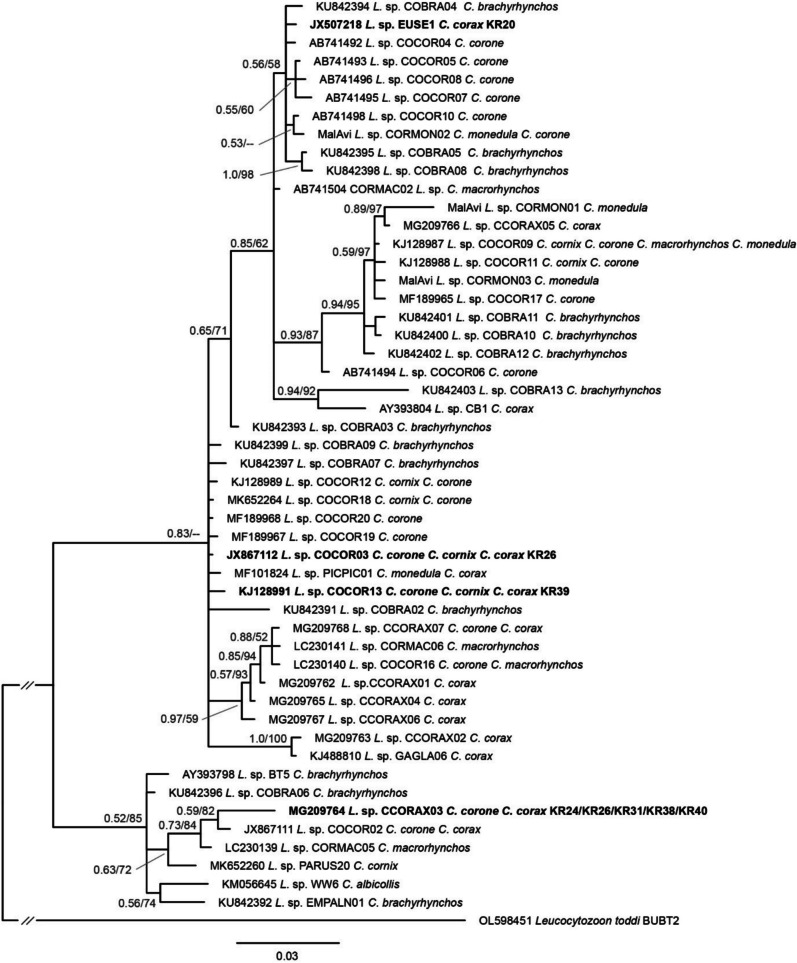 Figure 5
Figure 5- —University of Veterinary Medicine Vienna
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBird parasitology and diseases · Vector-borne infectious diseases · Parasite Biology and Host Interactions
Introduction
Blood parasites from the genera Plasmodium, Haemoproteus, and Leucocytozoon, which are transmitted by dipteran vectors such as mosquitoes, biting midges, and blackflies, are widespread among bird populations and can cause mild to severe infections and life-threatening diseases. Studying blood parasites in wild bird populations is important for understanding host-parasite co-evolution, the ecological role of parasitism, and the impact of environmental factors on transmission dynamics (Valkiunas 2004).
Common ravens (Corvus corax), known for their intelligence, adaptability, and wide distribution, are an intriguing species for studying blood parasite infections. As large, long-living scavengers, these ravens inhabit a variety of ecosystems, ranging from urban environments to remote wilderness areas, exposing them to diverse environmental pressures and parasite transmission risks. Social behavior, such as communal roosting and group foraging, may also increase the likelihood of parasite exposure and transmission (Boarman and Heinrich 1999). Despite their ecological significance, there is limited research on blood parasite prevalence and diversity in wild raven populations, with most parasite studies of birds focusing on smaller songbirds or captive populations (Shurulinkov et al. 2018).
Previous studies of avian blood parasites have shown variable infection rates, influenced by factors such as habitat, climate, and host characteristics such as age, sex, and immune status (Atkinson and Van Atkinson 1991; Hamer et al. 2013). Parasite prevalence and intensity often peak during the breeding season when birds experience increased stress and physiological demands (Marzal et al. 2005). Understanding how these factors impact raven populations is critical as parasitism could affect their fitness, reproductive success, and overall survival.
Recent studies highlight corvids as important hosts for haemosporidian parasites, particularly Leucocytozoon and Plasmodium. In Bulgaria, wild common ravens showed a 49% prevalence of Plasmodium and 31% of Leucocytozoon, with some Leucocytozoon lineages being raven-specific (Shurulinkov et al. 2018). In southwest Germany, Leucocytozoon was the most prevalent genus, infecting 85.3% of carrion crows (Corvus corone) and all sampled Eurasian magpies (Pica pica), with 65.3% of samples showing multiple infections (Schmid et al. 2017). These findings underscore the high diversity and prevalence of haemosporidian parasites in corvids. In Germany, no specific studies focusing on common ravens’ blood parasites were identified in the literature.
In this study we investigated the prevalence and diversity of blood parasite infections in common ravens from Germany. Using molecular diagnostic techniques, we evaluated the presence of filarioid nematodes, and members of the genera Trypanosoma, Plasmodium, Haemoproteus, and Leucocytozoon in 42 blood samples of common ravens collected during autopsy.
Material and methods
A total of 42 blood samples were collected from dead common ravens (15 male and 27 females) during autopsy. The birds were collected between September 2003 and August 2016 in four federal states (Thuringia, Hesse, Bavaria, and Lower Saxony) of Germany (Fig. 1). The location, date, sex, and growth stage (immature or adult) of the birds were recorded whenever possible. The age was determined using the usual throat coloration for Corvus species (Kalchreuter 1971). According to this, immature common ravens (< 2 years) have a red coloration and adult birds (> 2 years) have a black coloration. From each blood sample, DNA was extracted using the DNeasy Blood and Tissue Kit (QIAGEN, Hilden, Germany) according to the manufacturer’s instructions, and used to screen for DNA of filarioid nematodes, trypanosomatids, Plasmodium, Haemoproteus, and Leucocytozoon parasites.Fig. 1. Sampling locations in German federal states: Thuringia (TH), Hessen (HE), Bavaria (BV), and Lower Saxony (LS). The red dots 1 and 2 represent the number of samples per location; black lines represent the national border, grey lines the federal state border, and blue lines the main rivers. The graphic was created with CorelDRAW 2021 (Corel, Ottawa, Canada)
All PCRs were performed in 25 µl volumes with the GoTaq G2 Flexi DNA Polymerase kit (Promega GmbH, Walldorf, Germany). The mastermix contained 14.375 µl nuclease-free water, 5 µl 5X Green Reaction Buffer, 2 µl MgCl_2_ (25 mM), 0.5 µl dNTPs (10 mM), 0.125 µl GoTaq G2 Polymerase, each 1 µl each primer (10 mM), and 1–4 µl DNA template. The PCRs were run under the following conditions: initial denaturation for 2 min at 94 °C, followed by 35 cycles with 30 s at 94 °C, 30 s at X °C (see annealing temperatures provided for primers), 1 min at 72 °C, and a final extension for 10 min at 72 °C.
To detect filarioid nematodes, we used two separate primer sets targeting partial sequences of the nuclear 18S rRNA gene (18S) and the mitochondrial cytochrome c oxidase subunit I (COI). The primers ChandFO (5′-GAG ACC GTT CTC TTT GAG GCC-3′) and ChandRO (5′-GTC AAG GCG TAN NTT TAC CGC CGA-3′) (Hamer et al. 2013) target a 550 bp section of the 18S, and the primers COIintF (5′-TGA TTG GTG GTT TTG GTA A-3′) and COIintR (5′-ATA AGT ACG AGT ATC AAT ATC-3′) (Casiraghi et al. 2001; Merkel et al. 2007) target a 649 bp section of the COI. The annealing temperatures for the two primer sets were 57 °C (18S) and 51 °C (COI).
To detect Trypanosoma parasites, we used a nested PCR targeting an 870 bp section of the 18S as previously described (Peña-Espinoza et al. 2023) with the nest 1 primers Tryp_18S_F1 (5′-GTG GAC TGC CAT GGC GTT GA-3′) and Tryp_18S_R1 (5′-CAG CTT GGA TCT CGT CCG TT GA-3′) and the nest 2 primers Tryp_18S_F2 (5′-CGA TGA GGC AGC GAA AAG AAA TAG AG-3′) and Tryp_18S_R2 (5′-GAC TGT AAC CTC AAA GCT TTC GCG-3′). The annealing temperature used in the PCRs was 56 °C.
To detect haemosporidian parasites belonging to the genera Plasmodium, Haemoproteus, and Leucocytozoon, we used the common nested PCR assay (Hellgren et al. 2004), which targets a 476 bp section of the mitochondrial cytochrome b (CytB) gene. The first PCR was performed with the primers HaemNFI (5′-CAT ATA TTA AGA GAA NTA TGG AG-3′) and HaemNR3 (5′-ATA GAA AGA TAA GAA ATA CCA TTC-3′). The nested PCRs were performed with the primers HaemF (5′-ATG GTG CTT TCG ATA TAT GCA TG-3′) and HaemR2 (5′-GCA TTA TCT GGA TGT GAT AAT GGT-3) targeting Plasmodium and Haemoproteus parasites, and HaemFL (5′-ATG GTG TTT TAG ATA CTT ACA TT-3′) and HaemR2L (5′-CAT TAT CTG GAT GAG ATA ATG GNG C-3′) targeting Leucocytozoon parasites (Bensch et al. 2000; Hellgren et al. 2004). For the nested PCRs, each 1 µl of PCR product from the first PCR was used as template. The annealing temperature used for the PCRs with all three primer sets was 48 °C.
The PCR products were visualized on 1.8% agarose gels stained with Midori Green Advance (Biozym Scientific GmbH, Oldendorf, Germany) under UV light. PCR-positive samples were sent to LGC Genomics GmbH (Berlin, Germany) for purification and sequencing (using the PCR primers). The sequences were assembled and visually inspected using BioEdit v.7.7.1 (Hall 1999). For the phylogenetic analysis, additional nucleotide sequences were retrieved from NCBI GenBank using BLAST search and aligned and sorted with MAFFT v.7 (Katoh and Standley 2013) using the default options. Genetic distances were calculated with MEGA X v.10.0.5 (Kumar et al. 2018).
To infer the phylogenetic position of the Trypanosoma parasite lineage obtained in the present study, one of the 18S sequences was subjected to BLAST search on NCBI GenBank, and all sequences covering the entire 884 bp sequence section and showing a genetic similarity between 90 and 100% were downloaded. The resulting 336 hits from GenBank and the two sequences obtained in the present study were aligned and sorted with MAFFT v.7 (Katoh and Standley 2013) using the default options. The 338 sequences were aligned, and a maximum likelihood (ML) bootstrap tree (1000 replicates, model GTR + G + I) was calculated with IQ-TREE v.2.3.6 (Nguyen et al. 2015). Based on the latter tree (not shown), we identified the clade containing the Trypanosoma sequences obtained in the present study. Using the 21 sequences in the latter clade and a sequence of Trypanosoma melophagium (ON637628) as the outgroup, we calculated ML and Bayesian Inference (BI) trees. The alignment contained 903 positions, of which 32 positions contained gaps and were excluded before the analyses. The best-fit substitution model based on the corrected Akaike information criterion (cAIC) was evaluated with IQ-TREE v.2.3.6 (Nguyen et al. 2015), resulting in the model TIM3e + I + G4. Since the latter model is not implemented in MrBayes v.3.2.7 (Ronquist et al. 2012), the next similar model GTR + G + I was used for the phylogenetic analyses. The BI tree was calculated with MrBayes v.3.2.7 (Ronquist et al. 2012) by running 5 million generations and sampling every thousandth tree; the first 25% of trees were discarded as burn-in, and a 50% majority rule consensus tree was calculated from the remaining 3750 trees. An ML majority-rule consensus tree (1000 bootstrap replications) was calculated with IQ-TREE v.2.3.6 (Nguyen et al. 2015).
A similar approach was followed to determine the phylogenetic position of the Filarioidea identified in the present study based on partial 649 bp COI sequences. The COI sequence was subjected to BLAST search and all hits covering at least 90% of the sequence were downloaded. The sequences were aligned and sorted with MAFFT v.7 (Katoh and Standley 2013) using the default options. After removing all sequences with ambiguities, the alignment contained 991 sequences. An ML bootstrap tree (1000 replicates, model GTR + G + I) was calculated with IQ-TREE v.2.3.6 (Nguyen et al. 2015). Based on the latter tree (not shown), we identified a clade with maximum support containing the Filarioidea sequences obtained in the present study. Using the 34 sequences from the latter clade and a sequence of Dirofilaria repens (OP494253) as the outgroup, we calculated BI and ML trees applying the same settings as for the Trypanosoma 18S tree, using the model GTR + G + I as suggested by IQ-TREE v.2.3.6 (Nguyen et al. 2015).
For the phylogenetic tree based on partial 476 bp CytB sequences of Leucocytozoon parasites, we followed a different approach. Since more than 1000 different CytB lineages of avian Leucocytozoon parasites have been identified, we only included Leucocytozoon lineages found in Corvus species. The alignment included 50 unique lineages from seven different Corvus species and a sequence of Leucocytozoon toddi BUBT2 (OL598451) as the outgroup. The BI and ML trees were calculated using the same settings as the Trypanosoma and Filarioidea trees, using the model GTR + G + I as suggested by IQ-TREE v.2.3.6 (Nguyen et al. 2015).
All trees were visualized with FigTree v.1.4.3 (http://tree.bio.ed.ac.uk/software/figtree/) and graphically prepared with Adobe Illustrator CC v.2015 (Adobe Inc., San Jose, CA, USA).
The 95% confidence interval (CI) for prevalence percentages in the samples was calculated using the binomial proportion method with the following formula:
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${\varvec{C}}{\varvec{I}}={\varvec{p}}\pm {\varvec{Z}}\times \surd \left[{\varvec{p}}\left(1-{\varvec{p}}\right)/{\varvec{n}}\right]$$\end{document}where p is the observed prevalence (proportion of infected individuals), Z is the Z-score corresponding to the desired confidence level (e.g., 1.96 for 95% CI), and n is the sample size.
Results
The sample included 42 common ravens (KR1 to KR42) with 38.1% adult (16/42) and 61.9% immature individuals (26/42). The adults had a higher average weight (1206.9 g, 95% confidence interval CI: 1151.3–1262.5 g) compared to immatures (1061.2 g, 95% CI: 1023.4–1099.0 g). 35.7% were males (15/42) and 64.3% were females (27/42), with males showing a slightly higher average weight (1157.3 g, 95% CI: 1094.5–1220.1 g) than females (1110.7 g, 95% CI: 1074.6–1146.8 g).
The PCR screenings and sequencing revealed that altogether 11/42 birds were infected with blood parasites. Filarioid nematodes were present in seven common ravens (16.7%, 95% CI: 5.1–28.3%), Trypanosoma sp. in two individuals (4.8%, 95% CI: 0–11.4.3%), Leucocytozoon sp. in seven individuals (16.7%, 95% CI: 5.1–28.3%) with one individual harboring a co-infection with 2 lineages (Table 2), and Haemoproteus sp. in one individual (2.4%; 95% CI: 0–7.1%) (Tables 1 and 2; Fig. 2). Mixed infections were detected in five (11.9%) individuals, one triple infection with filarioids/Trypanosoma/Leucocytozoon, one double infection with filarioids/Haemoproteus, and three double infections with filarioids/Leucocytozoon (Tables 1 and 2; Fig. 2). Table 1. Prevalence and 95% confidence interval (CI) of parasitic infections in the 42 common ravens (Corvus corax) using binomial proportion methodInfection typeNumber of birdsPrevalence (%)95% confidence interval (CI)Single infectionsFilarioid nematodes716.7[5.1%, 28.3%]Trypanosoma sp.24.8[0%, 11.4%]Leucocytozoon sp.716.7[5.1%, 28.3%]Haemoproteus sp.12.4[0%, 7.1%]Mixed infectionsDouble infections49.5[0.7%, 18.3%]Triple infections12.4[0%, 7.1%]Total infected birds1126.2[13.1%, 39.3%]Table 2. Positive samples with their GenBank accession numbers and corresponding lineages and parasitesSample no**GenBank no.^^*ParasiteLineageClosest BLAST hitKR20PV236021Leucocytozoon sp.EUSE1JX507218 (100%)KR24PV236024Leucocytozoon sp.CCORAX03MG209764 (100%)KR25PV226108Trypanosoma sp.ANI54JN006849 (100%)KR26PV243878PV236148PV236023PV236028PV226109Filarioidea sp.Filarioidea sp.Leucocytozoon sp.Leucocytozoon sp.Trypanosoma sp.MI1 GLH-2012911E_MFCOCOR03CCORAX03ANI54JQ867040 (100%)MT800770 (92.64%)JX867112 (100%)MG209764 (100%)JN006849 (100%)KR30PV243879PV236149Filarioidea sp.Filarioidea sp.MI1 GLH-2012911E_MFJQ867040 (100%)MT800770 (92.64%)KR31PV236025Leucocytozoon sp.CCORAX03MG209764 (100%)KR32PV243880PV236149PV236020Filarioidea sp.Filarioidea sp.Haemoproteus sp.MI1 GLH-2012911E_MFCXPIP27/FES-2014/H360JQ867040 (100%)MT800770 (92.64%)KJ579154/KJ128983/KJ488905 (100%)KR36PV243881Filarioidea sp.MI1 GLH-2012JQ867040 (100%)KR38PV243882PV236026Filarioidea sp.Leucocytozoon sp.MI1 GLH-2012CCORAX03JQ867040 (100%)MG209764 (100%)KR39PV243883PV236022Filarioidea sp.Leucocytozoon sp.MI1 GLH-2012COCOR13JQ867040 (100%)KJ128991 (100%)KR40PV243884PV236027Filarioidea sp.Leucocytozoon sp.MI1 GLH-2012CCORAX03JQ867040 (100%)MG209764 (100%)^*^GenBank accession numbers of sequences from the present studyFig. 2Venn diagram of parasite prevalence in the studied samples. Numbers represent counts of common ravens (Corvus corax) with respective pathogens detected (blue for filarioid, green for Trypanosoma, yellow for Leucocytozoon, and purple for Haemoproteus). The overlapping colors and representing co-infections
Filarioid nematodes
Filarioid nematodes were found in seven (16.7%) common ravens (KR26, KR30, KR32, KR36, KR38, KR39, KR40) using the PCR targeting the 18S (Table 2). The 18S sequences (PV243878–PV243884) were all identical in the 547 bp section analyzed and 100% matched a Filarioidea sequence (JQ867040) isolated from Quiscalus quiscula in the USA. The next-best matches were Filarioidea sequences isolated from Eurasian blue tit (Cyanistes caeruleus) in Spain (OQ859189), a dog in the USA (MH390715) (Boyd et al. 2019), and American robin (Turdus migratorius) in the USA (JQ867026), with 99.82%, 99.62%, and 99.58% similarity, respectively. The COI sequences (PV236148–PV236150, Filarioidea sp.) obtained from three individuals (KR26, KR30, KR32) were identical. The closest matches to our Filarioidea sp. were Eufilaria sylviae from garden warbler (Sylvia borin) (MT800771, MT800770) with 92.64% and 92.43% similarity, and Eufilaria acrocephalusi from common reed warbler (Acrocephalus scirpaceus) (MT800769, MT800766) with 91.50% similarity, all of which were found in Lithuania. In the phylogenetic tree (Fig. 3), the COI lineage formed a single branch in a weakly supported clade (pp = 0.69/bs = 64), which contained Filarioidea lineages from various bird species assigned either to Eufilaria or Splendidofilaria.Fig. 3. Bayesian Inference tree of closely related Filarioidea parasites based on partial COI sequences (649 positions) from the present study and NCBI GenBank. Bayesian posterior probabilities and maximum likelihood bootstrap values are indicated at most nodes. The scale bar indicates the expected mean number of substitutions per site according to the model of sequence evolution applied. The sequences obtained in the present study are marked in bold letters
Trypanosoma parasites
Trypanosoma parasites were detected in two common ravens (KR25, KR26) (Table 2). The 18S sequences (PV226108–PV226109) were identical in the 884 bp section analyzed to a Trypanosoma sequence identified in Eurasian sparrowhawk (Accipiter nisus) from Czechia (JN006849). In the phylogenetic tree (Fig. 4), the lineage clustered together in a weakly supported clade (pp = 0.69/bs = 60) with Trypanosoma sequences found in various bird species, but the relationships between the lineages in the latter clade were mostly not resolved, presenting as a polytomy.Fig. 4. Bayesian Inference tree of closely related Trypanosoma parasites based on partial 18S sequences (871 positions excluding gaps) from the present study and NCBI GenBank. Bayesian posterior probabilities and maximum likelihood bootstrap values are indicated at all nodes. The scale bar indicates the expected mean number of substitutions per site according to the model of sequence evolution applied. The sequences obtained in the present study are marked in bold letters
Haemosporidian parasites
Leucocytozoon parasites were found in seven (16.7%) common ravens (KR20, KR24, KR26, KR31, KR38, KR39, KR40), with one individual presenting a mixed infection (PV236021–PV236028). The Leucocytozoon CytB lineages EUSE1 (JX507218), COCOR03 (JX867112), and COCOR13 (KJ128991) were found in one individual each, and CCORAX03 (MG209764) was found in five individuals (Table 2; Fig. 5). Haemoproteus parasites were found in one (2.4%) common raven (KR32; PV236020); the lineage CXPIP27 was previously isolated from Corvus corone in Portugal (KJ488905) and Culex pipiens in France (KJ579154). None of the haemosporidian parasite lineages has been linked to a morphological species yet. In the phylogenetic tree, the four Leucocytozoon lineages cluster together with numerous other lineages that had been found in C. corax and other Corvus species.Fig. 5. Bayesian Inference tree based on partial (476 bp) cytb sequences of Leucocytozoon parasites found in birds of the genus Corvus. A sequence of Leucocytozoon toddi (OL598451) was included as an outgroup. Bayesian posterior probabilities and maximum likelihood bootstrap values are indicated at all nodes. The scale bar indicates the expected number of substitutions per site according to the model of sequence evolution applied. The sequences obtained in the present study are marked in bold letters
Discussion
Many studies have investigated blood parasite prevalences in birds, but there are relatively few studies on Corvidae (Rodríguez and Matta 2001; Benedikt et al. 2009; Freund et al. 2016; Scaglione et al. 2016; Schmid et al. 2017). Our study confirmed the presence of trypanosomatids, filarioid nematodes, and haemosporidian parasites in Corvus corax by PCR and sequencing.
We found 16.7% of the common ravens to be infected with filarioid nematodes. This prevalence is slightly higher than the prevalence obtained by Hamer et al. (2013) with American robins and house sparrows (1.4–11.1%). Our study is the first to report filarioid nematodes in common ravens. Indeed, the few studies about Corvidae parasites did not investigate the prevalence of filarioid nematodes in these birds (Yoshimura et al. 2014; Freund et al. 2016; Scaglione et al. 2016; Schmid et al. 2017; Shurulinkov et al. 2018). The filarioid 18S rRNA sequences were identical to Onchocercidae sp. (JQ867040) from common grackles (Quiscalus quiscula) in the USA (Hamer et al. 2013), and the COI sequences matched closely with Eufilaria or Splendidofilaria from various bird species (Binkienė et al. 2021). The detection of the identical 18S rRNA sequence in 16.7% of the samples (2 adult and 5 immature birds) suggests that the detected filarioid circulates in the area studied and might have an impact on the common raven population. As mentioned by Hamer et al. (2013), lice, flies, and biting midges are likely vectors of the filarioid nematodes. Investigating the parasitemia of vectors collected directly on the host could bring more details regarding the source of infection.
The second parasite group with a significant prevalence of 16.7% was Leucocytozoon. This prevalence was lower compared to the 31.4% prevalence observed in Bulgarian ravens (Shurulinkov et al. 2018), and much lower compared to prevalences in other Corvus species from Germany, Italy, Japan, and the USA (Yoshimura et al. 2014; Freund et al. 2016; Scaglione et al. 2016; Schmid et al. 2017). The Leucocytozoon lineage CCORAX03 was found in five individuals; it was previously found in C. corax from Bulgaria (Shurulinkov et al. 2018) and C. corone in Germany (Strehmann et al. 2023). The lineage COCOR03 (identical to CORMAC03), present in one individual, was previously found in C. corone in Germany (Schmid et al. 2017) and hooded crow (Corvus cornix) in Italy (Scaglione et al. 2016). The lineage COCOR13, also present in one individual, was found in C. corone and C. cornix from Austria (Himmel et al. 2019; Harl et al. 2019), C. corone from Germany (Schmid et al. 2017), and C. cornix from Italy (Scaglione et al. 2016). The fourth lineage, EUSE1, was previously isolated from Eusimulium securiforme in Czechia (Synek et al. 2013) and Eurasian jay (Garrulus glandarius) from Spain (Illera et al. 2017).
Despite its broad presence, Leucocytozoon remains understudied, with significant gaps in understanding its taxonomy, transmission dynamics, and pathogenicity (Valkiūnas 2023). A relatively low prevalence of Leucocytozoon in common ravens were observed with a high genetic diversity. These findings highlight a relatively low prevalence of Leucocytozoon in common ravens with a genetic diversity of Leucocytozoon, emphasizing the need for further molecular studies to understand infection patterns and host-parasite coevolution.
The prevalence of Haemoproteus was much lower in the present study (2.7%) than in other studies of corvids (Murata 2002; Scaglione et al. 2016; Schmid et al. 2017). The lineage CXPIP27, found in one individual, was previously detected in C. cornix in Italy (Scaglione et al. 2016), C. corone in Portugal (Drovetski et al. 2014), and Culex pipiens in France (Zélé et al. 2014). This lineage’s presence in 3 species of the genus Corvus (C. Corax, C. Cornix, and C. Corone) and in vectors underscores a host adaptation and the importance of vector-host interactions in Haemoproteus transmission.
Trypanosoma parasites were detected in two common ravens (4.8%), a much lower prevalence compared to 17–18% in another study (Hamer et al. 2013). The Bayesian Inference tree of closely related Trypanosoma parasites based on partial 18S rRNA gene sequences revealed a Trypanosoma lineage which had previously been detected in Accipiter nisus from Czechia (Zídková et al. 2012). The study of Votypka et al. (2004) on the phylogenetic relationship of Trypanosoma corvi with other avian trypanosomes has revealed two well-supported monophyletic clades (the “Trypanosoma avium” clade and the “Trypanosoma corvi” clade) with no significant differences in their 18S rRNA sequences. Our study provides additional molecular data on Trypanosoma in common ravens, but it must be noted that Trypanosoma parasites are considerably less host-specific with respect to their bird hosts (Zídková et al. 2012).
The present study confirms the presence of filarioid nematodes, Leucocytozoon, Haemoproteus, and Trypanosoma parasites in common ravens (Corvus corax) and their circulation within the study area. Molecular evidence of co-infection with blood parasites in common ravens was identified, and we reported, for the first time, the presence of filarioid nemathodes in this specific Corvidae species. Our findings align with previous phylogenetic data, which suggest a variety of Leucocytozoon lineages in corvids, distributed across two distinct clades (Freund et al. 2016). Additionally, the results also highlight the genetic diversity and adaptability of Trypanosoma, which appears capable of infecting a wide range of avian hosts.
Conclusion
Overall, our results show that common ravens in Germany are infected with filarioid nematodes, Trypanosoma, Haemoproteus, and Leucocytozoon parasites. Three out of the four co-infected birds harbored filarioids and Leucocytozoon sp., and one bird was co-infected with filarioids and Haemoproteus sp. However, our study only found a small number of infected ravens within a geographically restricted area and, therefore, cannot provide evidence for the circulation of the parasites we found within the German raven population as a whole. An additional investigation on a larger population of common ravens in the studied areas is required to confirm and morphologically characterize the filarioid nematode species found in the present study.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Binkienė R, Chagas CRF, Bernotienė R, Valkiūnas G (2021) Molecular and morphological characterization of three new species of avian Onchocercidae (Nematoda) with emphasis on circulating microfilariae. Parasit Vectors 14. 10.1186/s 13071-021-04614-810.1186/s 13071-021-04614-8PMC 793443633673865 · doi ↗ · pubmed ↗
- 2Drovetski S V., Aghayan SA, Mata VA, et al (2014) Does the niche breadth or trade-off hypothesis explain the abundance-occupancy relationship in avian Haemosporidia? Mol Ecol 23. 10.1111/mec.1274410.1111/mec.1274424689968 · doi ↗ · pubmed ↗
- 3Freund D, Wheeler SS, Townsend AK, et al (2016) Genetic sequence data reveals widespread sharing of Leucocytozoon lineages in corvids. Parasitol Res 115. 10.1007/s 00436-016-5121-310.1007/s 00436-016-5121-327189064 · doi ↗ · pubmed ↗
- 4Hamer GL, Anderson TK, Berry GE, et al (2013) Prevalence of filarioid nematodes and trypanosomes in American robins and house sparrows, Chicago USA. Int J Parasitol Parasites Wildl 2. 10.1016/j.ijppaw.2012.11.00510.1016/j.ijppaw.2012.11.005PMC 386251224533314 · doi ↗ · pubmed ↗
- 5Harl J, Himmel T, Valkiunas G, Weissenböck H (2019) The nuclear 18S ribosomal DN As of avian haemosporidian parasites. Malar J 18. 10.1186/s 12936-019-2940-610.1186/s 12936-019-2940-6PMC 672429531481072 · doi ↗ · pubmed ↗
- 6Hellgren O, Waldenström J, Bensch S (2004) A new PCR assay for simultaneous studies of Leucocytozoon, Plasmodium, and Haemoproteus from avian blood. Journal of Parasitology 90. 10.1645/GE-184R 110.1645/GE-184R 115357072 · doi ↗ · pubmed ↗
- 7Himmel T, Harl J, Kübber-Heiss A, et al (2019) Molecular probes for the identification of avian Haemoproteus and Leucocytozoon parasites in tissue sections by chromogenic in situ hybridization. Parasit Vectors 12. 10.1186/s 13071-019-3536-210.1186/s 13071-019-3536-2PMC 654760931159851 · doi ↗ · pubmed ↗
- 8Illera JC, López G, García-Padilla L, Moreno Á (2017) Factors governing the prevalence and richness of avian haemosporidian communities within and between temperate mountains. P Lo S One 12. 10.1371/journal.pone.018458710.1371/journal.pone.0184587 PMC 558924128880919 · doi ↗ · pubmed ↗