Involvement of a NIMA-related kinase in cell division of the liverwort Marchantia polymorpha
Hikari Mase, Aoi Sumiura, Yoshihiro Yoshitake, Takayuki Kohchi, Taku Takahashi, Hiroyasu Motose

TL;DR
This study shows that a NIMA-related kinase in a liverwort regulates cell division and growth, similar to its role in fungi and animals.
Contribution
The novel contribution is identifying MpNEK1's role in cell division and growth suppression in Marchantia polymorpha.
Findings
Overexpression of MpNEK1 suppresses cell division and causes lethality in juvenile plants.
MpNEK1 induction reduces microtubule density and dynamics without affecting cell morphology.
Growth suppression by MpNEK1 is partially independent of its kinase activity.
Abstract
Never-in-mitosis A (NIMA)-related kinases (NEKs) regulate a series of mitotic events in fungi and animals, whereas plant NEKs have been shown to control the growth direction of cells and organs. Plant NEKs are highly expressed in the meristem, but whether they regulate meristematic activity remains unknown. The liverwort Marchantia polymorpha has a single functional MpNEK1 gene, and its knockout results in twisted rhizoid growth. For a gain-of-function approach, we generated lines for the inducible expression of MpNEK1 using an estrogen receptor-mediated system. Estradiol treatment effectively induced the accumulation of MpNEK1 mRNA and MpNEK1–Citrine fusion protein throughout the plant. MpNEK1 overexpression severely suppressed rhizoid and thallus growth, ultimately leading to the lethality of juvenile plants. This severe effect was observed even at the nanomolar level of estradiol.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
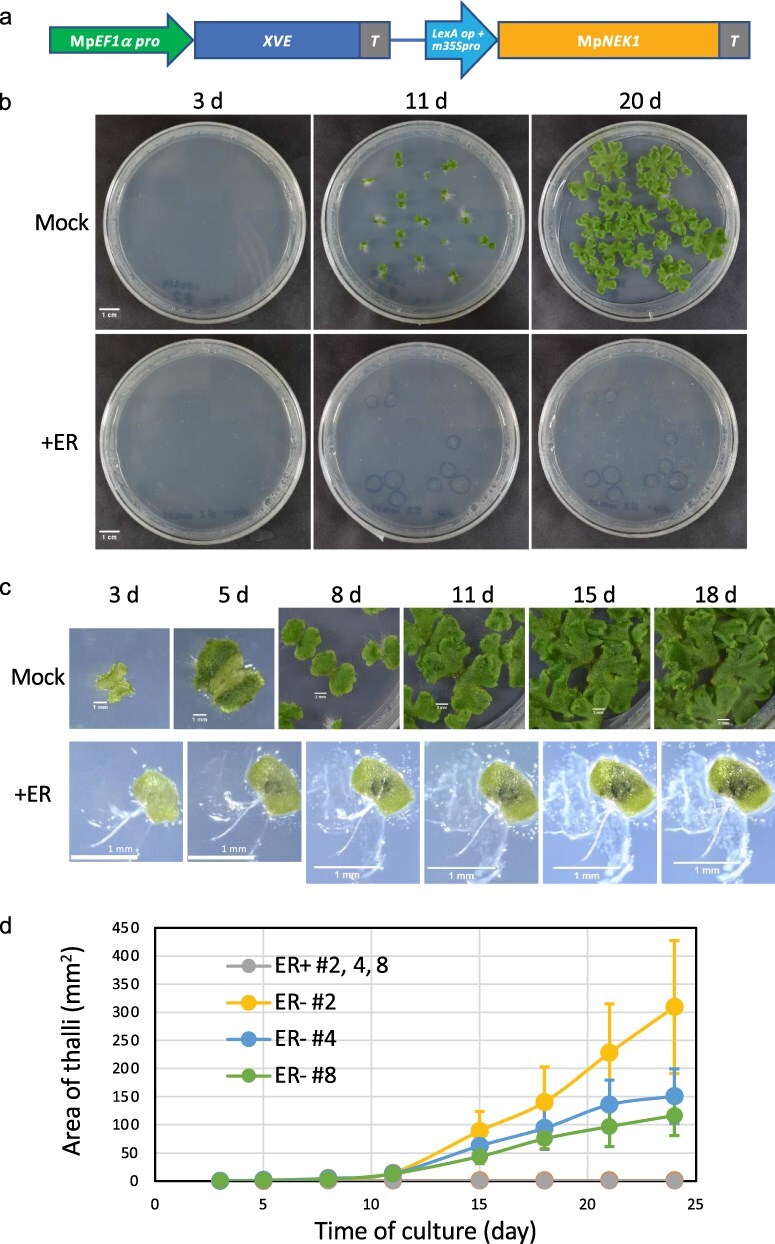 Figure 1
Figure 1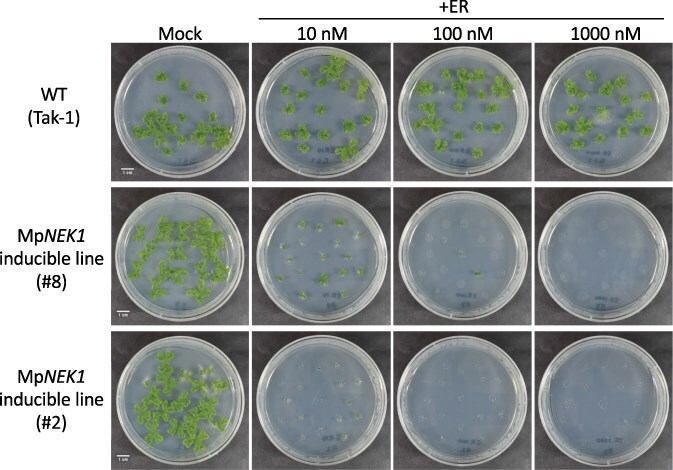 Figure 2
Figure 2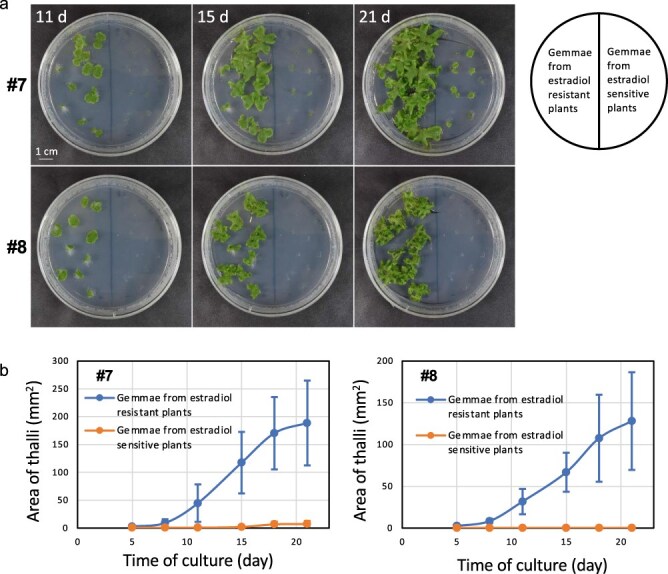 Figure 3
Figure 3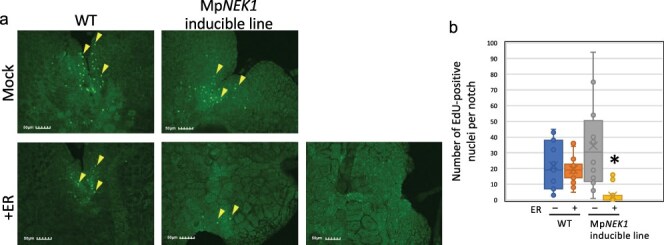 Figure 4
Figure 4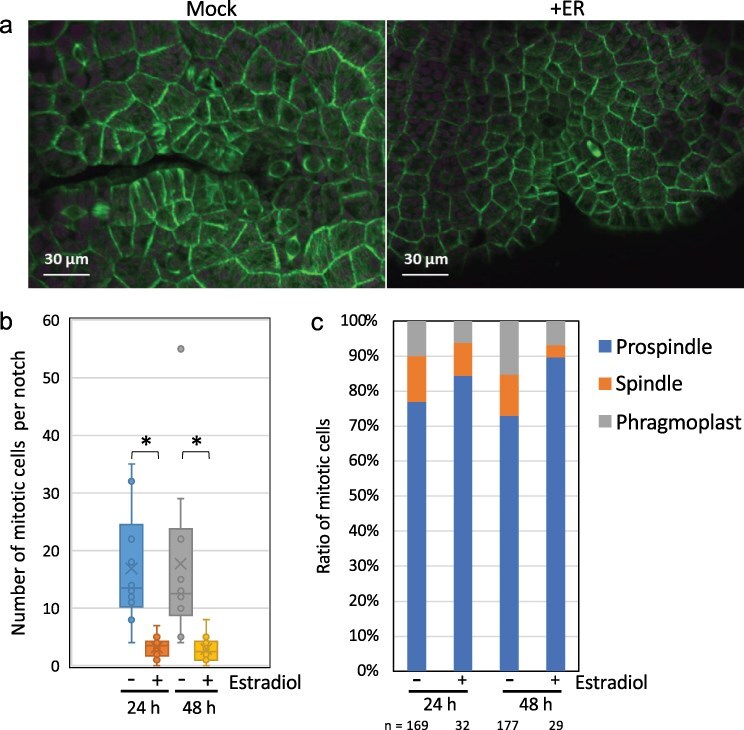 Figure 5
Figure 5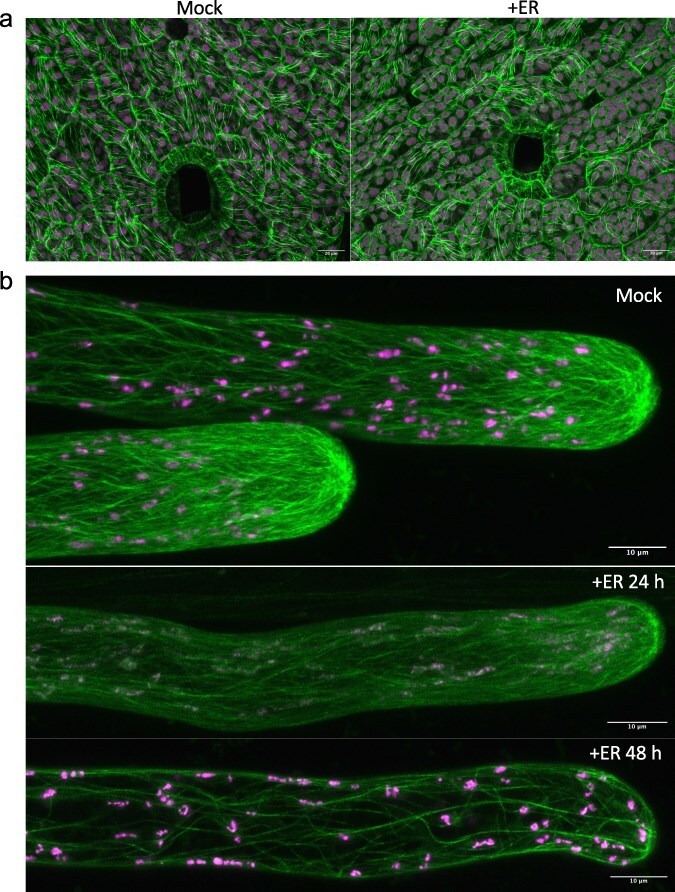 Figure 6
Figure 6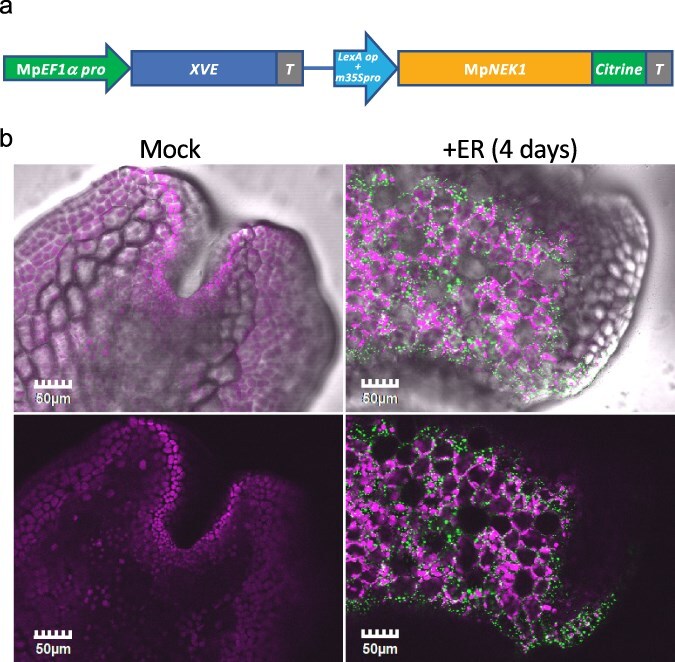 Figure 7
Figure 7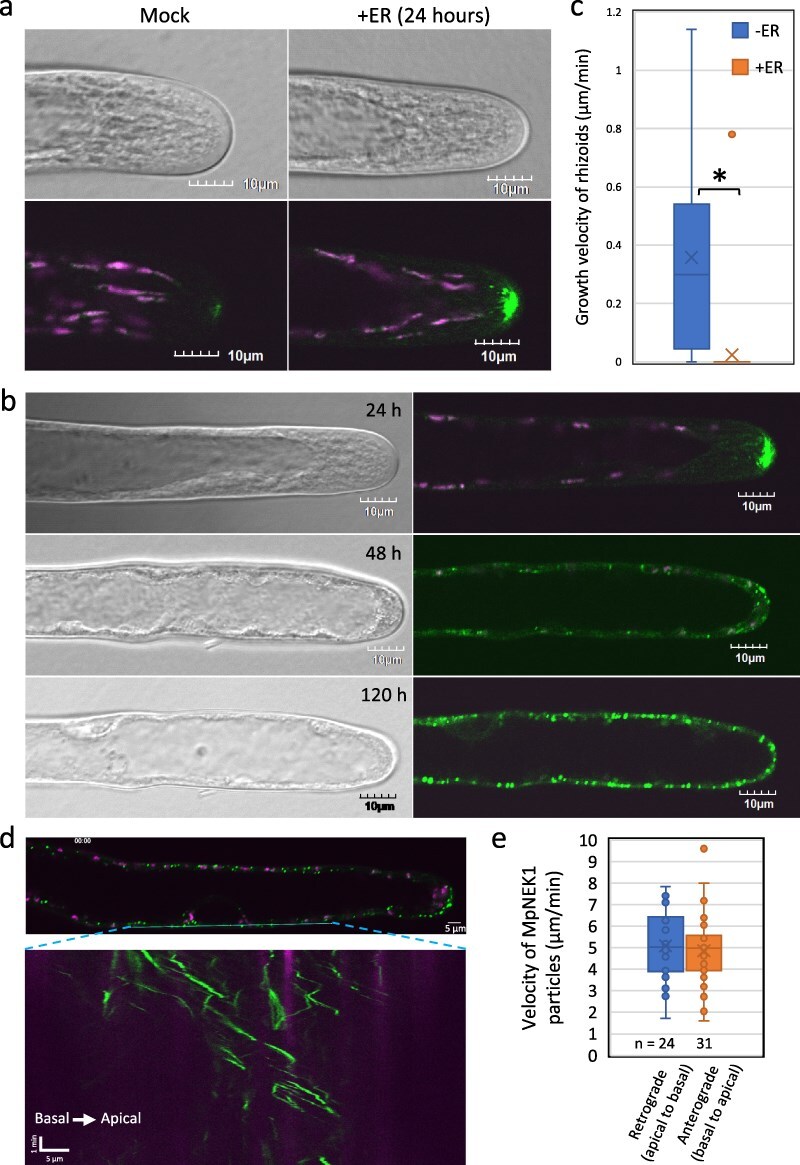 Figure 8
Figure 8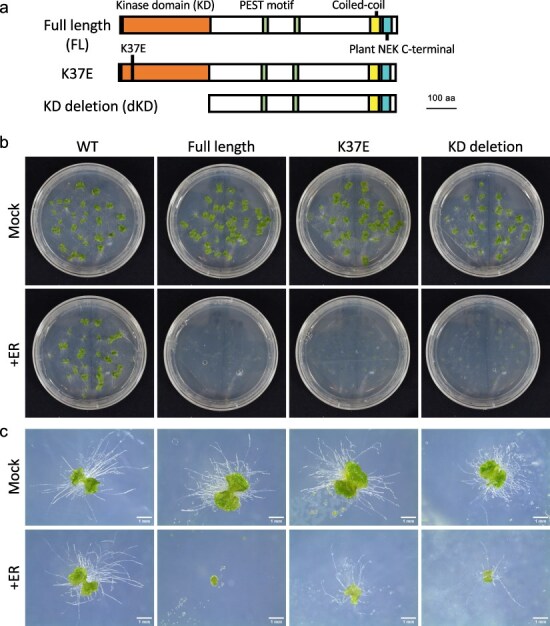 Figure 9
Figure 9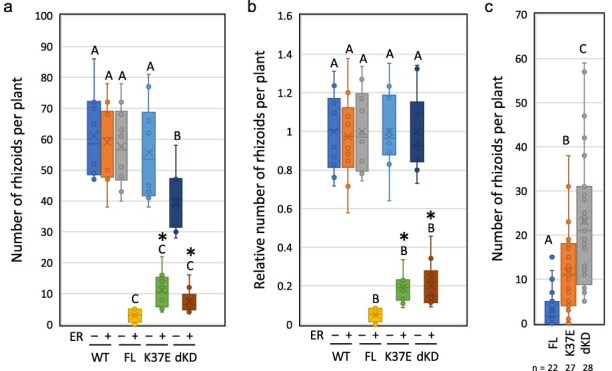 Figure 10
Figure 10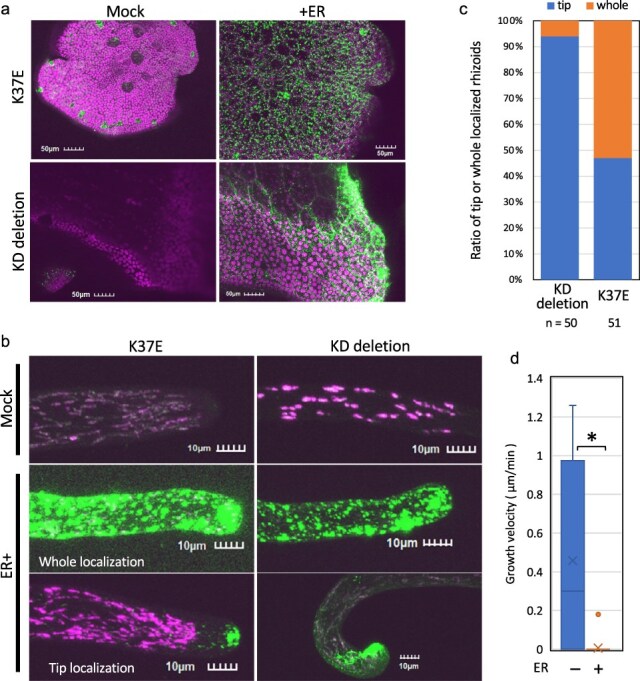 Figure 11
Figure 11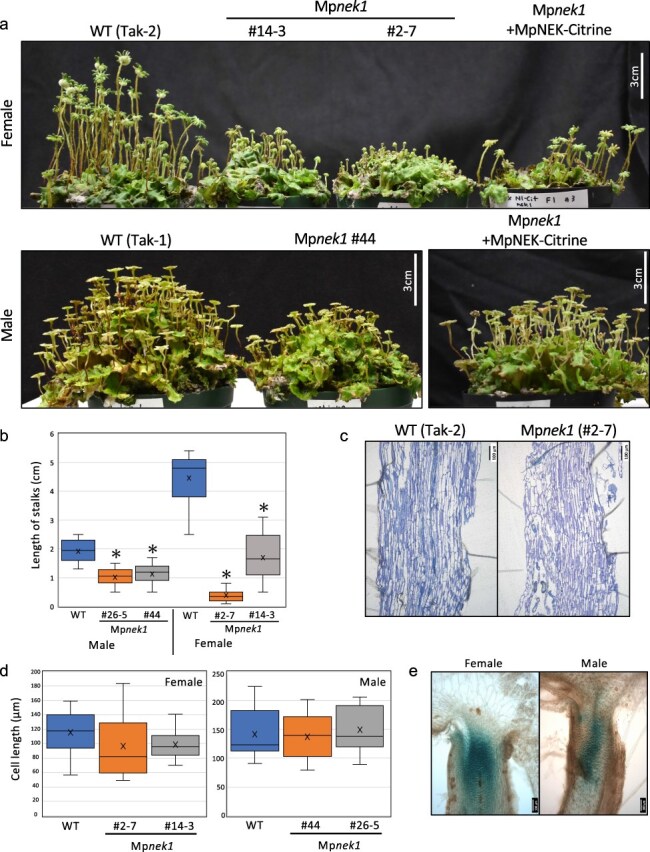 Figure 12
Figure 12| Mock ( | +Estradiol ( | |
|---|---|---|
| Microtubule | ||
| Bundling (skewness) | 1.63 ± 0.29 | 1.52 ± 0.24 |
| %Occupancy | 4.88 ± 0.72 | 3.79 ± 0.78 |
| Cell | ||
| Area (µm2) | 690.2 ± 183.6 | 694.9 ± 235.4 |
| Circularity | 0.72 ± 0.09 | 0.72 ± 0.10 |
| Aspect ratio | 2.00 ± 0.60 | 2.11 ± 0.71 |
| Roundness | 0.54 ± 0.14 | 0.52 ± 0.16 |
| Parameters | Mock ( | +Estradiol ( |
|---|---|---|
| Growth (µm/min) | 3.63 ± 1.45 | 2.35 ± 1.12 |
| Shrinkage (µm/min) | 6.96 ± 2.48 | 6.01 ± 2.96 |
| Catastrophe (events/sec) | 0.022 ± 0.026 | 0.011 ± 0.007 |
| Rescue (events/sec) | 0.032 ± 0.027 | 0.030 ± 0.052 |
| Dynamicity (µm/min) | 3.75 ± 2.19 | 2.55 ± 2.14 |
| Time spent (%) | ||
| Growth | 39.5 | 47.1 |
| Pause | 35.4 | 33.1 |
| Shrinkage | 25.1 | 19.4 |
- —Naito Foundation10.13039/100007428
- —Ryobi Teien Memory Foundation10.13039/501100013171
- —Jsps
- —Naito Foundation10.13039/100007428
- —Ryobi Teien Memory Foundation10.13039/501100013171
- —Jsps
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBryophyte Studies and Records · Plant Reproductive Biology · Mycorrhizal Fungi and Plant Interactions
Introduction
Never-in-mitosis A (NIMA)-related kinase (NEK) is a Ser/Thr protein kinase that controls a series of mitotic events in fungi and animals (O’Connell et al. 2003, Fry et al. 2012), while plant NEK members have been shown to regulate directional growth (Motose et al. 2008, Sakai et al. 2008). Arabidopsis NEK6 suppresses ectopic outgrowth of epidermal cells through microtubule organization and interaction with other NEK members (Motose et al. 2011). NEK6 localizes to the shrinking ends of microtubules, phosphorylates five amino acid residues of β-tubulin, and depolymerizes aberrant cortical microtubules to promote anisotropic growth of epidermal cells (Takatani et al. 2017). Furthermore, NEK6 has been shown to stabilize the growth direction of hypocotyls by selectively removing cortical microtubules that best align with maximal tensile stress (Takatani et al. 2020). NEK6-mediated attenuation of tensile stress response increases the noise in microtubule orientation to buffer mechanical conflicts and stabilize growth direction.
The liverwort Marchantia polymorpha is an emerging model to study the evolution of land plants (Kohchi et al. 2021) because of its low genetic redundancy (Bowman et al. 2017) and various genetic tools (Ishizaki et al. 2015, 2016). Indeed, Arabidopsis has seven NEK members, whereas M. polymorpha has a single NEK gene, MpNEK1 (Takatani et al. 2015, Otani et al. 2018). Thanks to these advantages, MpNEK1 has been shown to direct tip growth of rhizoids through microtubule depolymerization in the apical region (Otani et al. 2018). Rhizoids are filamentous rooting cells, mainly elongating from the ventral epidermis and are thought to be required for plant anchorage to the soil and water/nutrient uptake (Jones and Dolan 2012, Shimamura 2016, Kohchi et al. 2021, Kanda et al. 2022). Mpnek1 mutant rhizoids frequently change their growth direction, resulting in the zigzag morphology (Otani et al. 2018) and the defect of invasive growth to enter the hard substratum (Mase et al. 2022). MpNEK1 localization in the apical microtubule foci and the increased microtubule stability in Mpnek1 mutants demonstrate that MpNEK1 reorganizes and destabilizes apical microtubules to maintain the growth polarity of rhizoids (Otani et al. 2018). A similar phenotype was observed in the mutants of a microtubule-associated protein, WAVE DANPENED-LIKE (MpWDL) (Honkanen et al. 2016, Champion et al. 2021). MpWDL might stabilize longitudinal microtubules in the shank region of rhizoids. Thus, microtubule organization and dynamics are essential for the directional growth of rhizoids.
However, it is not clear how MpNEK1 regulates microtubules to stabilize growth direction. Furthermore, the functional significance of MpNEK1 expression in the meristem remains unclear. The gain-of-function approach could be helpful to elucidate these problems and identify downstream proteins that are phosphorylated by MpNEK1. In this study, we used an estradiol-inducible system to overexpress MpNEK1 in M. polymorpha and investigate its effects on plant growth and development.
Results
Establishment of MpNEK1 inducible lines
In this study, we generated MpNEK1-inducible transgenic lines using the XVE chimeric transcriptional activator (Zuo et al. 2000). This activator is composed of the DNA-binding domain of LexA, the transcriptional activation domain of VP16, and the estradiol-binding domain of the human estrogen receptor. This system has no apparent toxic physiological effects and has been effectively used in flowering plants. The XVE vector was optimized for the application to M. polymorpha in several points. Because the endogenous elongation factor MpEF1α promoter is more active in the meristem and reproductive organ than the cauliflower mosaic virus 35S promoter (Althoff et al. 2014), the MpEF1α promoter was utilized to drive XVE. XVE system has been applied in several studies in M. polymorpha (e.g. Flores-Sandoval et al. 2016, Furuya et al. 2022, Bao et al. 2024) but not fully characterized.
We isolated eight transgenic lines by transformation of wild-type (WT) sporelings with pMpGWB344-MpNEK1 (MpEF1α pro:XVE≫LexAop:MpNEK1), in which XVE is expressed under the control of MpEF1α promoter and activates transcription of MpNEK1 through the binding to the LexA operator in the presence of estradiol. Among them, seven transgenic lines (Lines #2–8) were almost identical to the WT in the growth and morphology of thalli. We examined MpNEK1 expression in these transgenic lines by quantitative real-time polymerase chain reaction (RT-qPCR) (Supplementary Fig. S1). Two-week-old thalli of the WT and transgenic lines were transferred to the medium with or without β-estradiol and incubated for 24 hours. MpNEK1 expression remarkably increased in most transgenic lines (#2, 4, 5, 7, and 8) by a factor of 4–15 in the estradiol treatment than in the estradiol-free medium. Estradiol did not induce MpNEK1 expression in the WT and Line #6. MpNEK1 induction was reproducible and reliable, especially in Lines #2, 4, and 8, which exhibited no obvious defects in growth and morphology without estradiol. Thus, these lines were used for further analyses.
MpNEK1 induction severely suppresses thallus growth
We conducted a phenotypic analysis to assess the effects of estradiol-induced MpNEK1 on growth and development. The gemmae of the WT and MpNEK1-inducible lines were grown in the presence or absence of exogenous β-estradiol for 24 days. There was no obvious effect of β-estradiol on the growth and morphology of the WT thalli (Supplementary Fig. S2). In the inducible lines, thallus growth was severely suppressed when cultured with β-estradiol (Fig. 1). Most gemmalings stopped growing and died in the presence of estradiol (Fig. 1), whereas the non-inducible line #6 did not show growth retardation (Supplementary Fig. S3). These results clearly demonstrate that the induction of MpNEK1 severely suppresses thallus growth and causes juvenile lethality.
Effect of estradiol on thallus growth of MpNEK1 inducible line. (a) Construct for MpNEK1 induction. MpEF1a promoter, terminator (T), LexA operator (LexA op), minimal CaMV35S promoter (m35Spro). The diagram is not shown to scale. (b) The gemmae of MpNEK1-inducible line (#2) were planted in the agar medium with (+ER) or without 10 µM estradiol (Mock) and grown for 3, 11, and 20 days. (c) Time course of growth of MpNEK1-inducible line (#2) with (+ER) or without 10 µM estradiol (Mock). (D) Quantification of thallus growth of MpNEK1-inducible line (#2, 4, 8) with (ER+) or without 10 µM estradiol (ER−). The mean projection area of thalli (n = 8–11 plants) was quantified by ImageJ. Circles and error bars indicate mean values and standard deviations, respectively.
We next examined whether the effect of MpNEK1 induction is reversible or not. The gemmalings of the WT and MpNEK1-inducible lines were grown in the presence of estradiol and then transferred to the estradiol-free medium at various times after estradiol treatment (Supplementary Fig. S4). After 1- and 3-day treatment with estradiol, thallus growth seemed to be recovered, whereas irreversible growth suppression was observed after 7- and 14-day treatment. In the WT, the growth inhibitory effect of estradiol was not observed. Therefore, 1-week MpNEK1 induction (estradiol treatment) confers an irreversible effect on thallus growth.
In the XVE induction system in flowering plants, 2–10 µM concentrations of estradiol have been commonly used. In the MpNEK1-inducible lines, 10 µM estradiol caused severe growth suppression and lethality. However, more mild conditions would be desirable to further investigate the MpNEK1 function. To determine the less-effective conditions and dose-dependent effect of estradiol, thalli of the WT and MpNEK1-inducible lines were incubated in the presence of estradiol at various concentrations (Fig. 2, Supplementary Fig. S5). The WT thallus showed no obvious phenotype in all concentrations tested, whereas thallus growth was suppressed in the MpNEK1-inducible lines even at 10 and 100 nM of estradiol. From this result, MpNEK1 induction has a severe effect on thallus growth.
Effect of various concentrations of estradiol on thallus growth. The gemmae of the WT and MpNEK1-inducible lines were planted in the agar medium supplemented with or without estradiol at concentrations of 10, 100, or 1000 nM and grown for 16 days.
We noticed that some plants obtained the resistance to estradiol and survived in the agar medium supplemented with estradiol (Fig. 3). The estrogen resistance was observed in the next-generation gemmae, suggesting the inheritance of gene silencing from the thallus to gemma. These estradiol-resistant plants appeared more frequently at lower concentrations and exhibited no induction of MpNEK1 (Supplementary Fig. S6). At the lower concentrations, larger numbers of thallus cells could survive and acquire estradiol resistance. Thus, lower concentrations of estradiol create an undesirable condition that can lead to estradiol resistance.
Estradiol resistance and its transfer to the next generation. (a) The gemmae of the estradiol-resistant plants or sensitive plants of MpNEK1-inducible lines were planted in the agar medium supplemented with 10 µM estradiol and grown for 11, 15, and 21 days. (b) Quantification of growth of gemmalings derived from the estradiol-resistant plants or -sensitive plants of MpNEK1-inducible lines. The mean projection area of thalli (n = 10 plants) was quantified by ImageJ. Circles and error bars indicate mean values and standard deviations, respectively.
MpNEK1 induction suppresses cell proliferation
Previous studies show that plant NEKs mainly regulate the growth direction of cells and organs. However, severe suppression of thallus growth by estradiol-induced MpNEK1 suggests some roles of plant NEK in cell division and proliferation. To determine whether estradiol-induced MpNEK1 affects cell division, we monitored cell proliferation activity by staining with EdU (5-ethynyl-2ʹ-deoxyuridine) (Fig. 4, Supplementary Fig. S7). This thymidine analog can be incorporated into DNA during the S phase. The subsequent reaction of EdU with a fluorescent azide labels newly replicated DNA strands of the S-phase cells during the incubation with EdU.
Effect of estradiol on cell proliferation. (a) EdU-labeled nuclei in the gemmalings treated with 1 µM estradiol for 4 days. The gemmae of the WT and MpNEK1-inducible line were planted in the medium supplemented with or without 1 µM estradiol, grown for 4 days, and incubated with 10 µM EdU for 1 hour in the same kind of liquid medium. EdU-labeled nuclei were visualized according to the manufacturer’s instructions as described in methods. (b) Quantification of EdU-labeled nuclei in the thalli grown as in (a). Data are shown by box plots (n = 12–16 plants). An asterisk indicates significant difference from the control (−ER) (t-test, P < .05).
In both the WT and inducible lines, EdU-positive nuclei were observed in the meristematic notch region in thalli grown in the absence of estradiol (Fig. 4, Supplementary Fig. S7). The number of EdU-positive nuclei was not affected in the WT by estradiol, whereas it was significantly reduced in the estradiol-treated inducible lines. Hence, estradiol-induced MpNEK1 severely suppressed cell proliferation.
MpNEK1 induction suppresses mitotic cell division
Because Arabidopsis NEK6 depolymerizes specific microtubules, which detached from the plasma membrane or aligned to the maximal tensile stress, to regulate cell expansion and organ growth (Takatani et al. 2017, 2020), we monitored the effect of MpNEK1 overexpression on microtubule organization and mitotic apparatus. MpNEK1-inducible construct was introduced into the transgenic lines expressing a microtubule marker Citrine–MpTUB2 to generate the inducible lines expressing Citrine–MpTUB2. The gemmae of these transgenic plants were grown for 3 days in the estradiol-free medium and then applied to the liquid medium with or without 10 µM estradiol. After 24 and 48 hours of treatment, thallus cells were observed with confocal microscopy. In the estradiol-treated thalli, the number of mitotic cells was significantly decreased compared with the mock-treated thalli (Fig. 5a and b, Supplementary Fig. S8). We classified mitotic cells according to mitotic structures, prospindle, spindle, and phragmoplast since these structures are clearly distinguished under confocal microscopy (Buschmann et al. 2016). We observed a slight increase of cells with prospindle and a slight decrease of cells with spindle and phragmoplast (Fig. 5c), suggesting that MpNEK1 induction has a mild effect on mitotic progression (e.g. transition from prospindle to spindle formation). In summary, overexpression of MpNEK1 significantly suppressed mitotic cell division.
Effect of MpNEK1 induction on mitosis. (a) Mitotic cells in the apical notches. The gemmae of MpNEK1-inducible line with a microtubule marker CaMV35S:Citrine–MpTUB2 were grown in the estradiol-free medium for 3 days and then supplemented with the liquid medium with (+ER) or without 10 µM estradiol (Mock). Thalli were observed under a confocal microscope after 24 hours of treatment. Green: Microtubule, magenta: plastid autofluorescence. (b) Number of mitotic cells per notch in the MpNEK1 inducible line with CaMV35S:Citrine–MpTUB2 after 24 and 48 hours of treatment with or without estradiol. Data are shown in the box plot (n = 10 notches, N > 6 plants). Asterisks indicate significant difference (t-test, P < .01). (c) The classification of mitotic cells with prospindle, spindle, or phragmoplast (n indicates the number of mitotic cells used for the classification).
MpNEK1 induction reduces microtubule density and dynamics
We next analyzed the effects of MpNEK1 induction on cell growth and cortical microtubule organization (Fig. 6). Estradiol treatment did not obviously affect the morphology of thallus epidermal cells but seemed to disorganize cortical microtubules (Fig. 6a). These were confirmed by the quantification of parameters of epidermal cells and microtubules (Table 1). Cell area and morphological parameters were not severely affected by estradiol treatment but both bundling and density of cortical microtubules were decreased. The quantification of the dynamic parameters of cortical microtubules showed that MpNEK1 induction decreased growth rate, catastrophe events, and dynamicity of microtubule plus ends (Table 2). These results show that MpNEK1 induction decreases microtubule density and dynamics in the thallus.
Effect of MpNEK1 induction on microtubules. (a) Cortical microtubules in thallus epidermal cells. The gemmae of MpNEK1 inducible line with CaMV35S:Citrine–MpTUB2 were grown in the estradiol-free medium for 6 days and then supplemented with the liquid medium with (+ER) or without 10 µM estradiol (Mock). Thalli were observed under a confocal microscope after 24 hours of treatment. Green: Microtubule, magenta: plastid autofluorescence. (b) Microtubules in rhizoids. The gemmae of MpNEK1-inducible line with CaMV35S:Citrine–MpTUB2 were grown as in (a). Rhizoids were observed under a confocal microscope after 24 and 48 hours of treatment. Green: microtubule, magenta: plastid autofluorescence.
We also analyzed the effects of MpNEK1 induction on microtubule organization in rhizoids (Fig. 6b). In the control condition, most microtubules in the shank region aligned along the longitudinal direction and exhibited cable-like bundles, whereas microtubule foci were observed at the apical dome of rhizoids (n = 16/16, 16 rhizoids with microtubule foci/all 16 rhizoids). After 24 hours of estradiol treatment, microtubule signal was reduced but microtubule foci still existed in ∼70% of rhizoids (n = 12/17). After 48 hours of estradiol treatment, microtubules were further reduced and microtubule foci were absent in 80% of rhizoids (n = 3/15, 3 rhizoids with microtubule foci/all 15 rhizoids). The cable-like bundles still existed and extended toward the apical region. Concomitant with the lack of microtubule foci, rhizoids grow more wavy after 24–48 hours of estradiol treatment. Thus, MpNEK1 induction suppressed microtubule foci at the apical dome of rhizoids.
The overaccumulation of MpNEK1–Citrine suppresses thallus growth
We generated inducible lines of MpNEK1–Citrine fusion protein (Fig. 7), in which protein induction and localization could be easily detected by confocal microscopy, facilitating functional analysis of MpNEK1. Eight transgenic lines were isolated by the transformation of WT sporelings with pMpGWB144–MpNEK1–Citrine. Among them, four transgenic lines (Lines #1, 5, 7, and 8) were almost identical to the WT in the absence of estradiol. When grown in the agar medium supplemented with estradiol, these transgenic lines exhibited severe growth retardation and juvenile lethality (Supplementary Fig. S9).
Localization of estradiol-induced MpNEK1–Citrine in the thallus. (a) Construct for MpNEK1 induction. MpEF1a promoter, terminator (T), LexA operator (LexA op), and minimal CaMV35S promoter (m35Spro). The diagram is not shown to scale. (b) The gemmae of the MpNEK1-inducible line were planted in the medium with (+ER) or without 1 µM estradiol (Mock), grown for 4 days, and observed under a confocal microscope. Upper panels are light field images merged with confocal images shown in the lower panels. Green: MpNEK1–Citrine, magenta: plastid autofluorescence.
We next analyzed the expression and localization of the MpNEK1–Citrine fusion protein (Fig. 7b, Supplementary Fig. S10A). The gemmae of MpNEK1–Citrine-inducible lines were grown for 4 days in the medium with or without 1 µM β-estradiol and observed under a confocal microscope (Fig. 7b). MpNEK1–Citrine was not detected in the absence of estradiol, whereas it was accumulated throughout estradiol-treated thalli (Fig. 7b, Video S1). MpNEK1–Citrine localized in the cytoplasmic particles. In the 24-hour estradiol treatment, similar expression and localization pattern were observed (Supplementary Fig. S10A). From these results, MpNEK1–Citrine was induced throughout the thallus within 24 hours after estradiol treatment and severely suppressed thallus growth.
We noticed an accumulation of MpNEK1–Citrine particles in the meristem in some transgenic plants even in the absence of estradiol (Supplementary Fig. S10B). This leaky expression level varied in each individual and transgenic line. In this study, we used plants and transgenic lines (Lines #1, 5, 7, and 8) that exhibited reduced leaky expression of MpNEK1–Citrine.
The overaccumulation of MpNEK1–Citrine suppresses rhizoid growth
Taking account that MpNEK1 localizes in the apical microtubule foci to direct tip growth of rhizoids (Otani et al. 2018), we analyzed the localization of estradiol-induced MpNEK1–Citrine in tip-growing rhizoids (Fig. 8). Because rhizoids were not formed when gemmae were grown with estradiol at the start of culture, we added estradiol to 3-day-old gemmalings that had been grown without estradiol to investigate the localization and effects of MpNEK1–Citrine over time. After 24-hour treatment, MpNEK1–Citrine accumulated in the apical region of the rhizoids (Fig. 8a, Video S2). MpNEK1–Citrine expression varied in each rhizoid. A few rhizoids showed highly accumulated MpNEK1–Citrine, which formed large droplets, moved to the apical direction, and fused with each other (Video S3). To determine when MpNEK1–Citrine starts to accumulate, we observed rhizoids at the early periods of estradiol treatment (Supplementary Fig. S10C). MpNEK1–Citrine began to accumulate at ∼1 hour of treatment and gradually increased in the rhizoid apex. After 48 and 120 hours, MpNEK1–Citrine accumulated throughout the entire region of rhizoids in the particle and filamentous patterns (Fig. 8b, Videos S4–S7). Tip growth ceased or severely reduced after 48 and 120 hours of estradiol treatment (Fig. 8c, Videos S4–S6). After 48 hours, movement of MpNEK1–Citrine particles was obvious (Fig. 8d, Videos S4 and S5). MpNEK1–Citrine particles moved toward the apex (anterograde) and basal region (retrograde) at the same velocity (Fig. 8e). These results suggest that overexpression of MpNEK1 causes its mislocalization throughout the entire length of rhizoids to suppress rhizoid growth.
Localization of estradiol-induced MpNEK1–Citrine in rhizoids. (a) The gemmae of the MpNEK1-inducible line were planted in the medium without estradiol, grown for 3 days, and then supplemented with (+ER) or without 1 µM estradiol (Mock). Rhizoids were observed under a confocal microscope. Green: MpNEK1–Citrine, magenta: plastid autofluorescence. (b) The gemmae of the MpNEK1-inducible line were grown and treated with estradiol as in (a) and observed at the time indicated in each panel. The same rhizoid was observed under a confocal microscope over time. Green: MpNEK1–Citrine, magenta: plastid autofluorescence. (c) Growth velocity of rhizoids in the MpNEK1–Citrine-inducible line incubated for 48 hours with (+ER) or without 1 µM estradiol (−ER). Data are shown by box plots (n = 32 rhizoids). An asterisk indicates significant difference (t-test, P < .01). (d) Movement of MpNEK1–Citrine particles in rhizoids. The gemmae of the MpNEK1-inducible line were grown and treated as in (b). Rhizoids were observed under a confocal microscope after 48 hours. The lower panel is a kymograph of MpNEK1–Citrine particles. Green: MpNEK1–Citrine, magenta: plastid autofluorescence. (e) Velocity of MpNEK1–Citrine particles. Data are shown by box plots (n = 24 or 31 particles). There was no significant difference (t-test, P > .05).
The overexpression of kinase-deficient MpNEK1 suppresses growth of thalli and rhizoids
To determine whether the kinase activity of MpNEK1 is required for growth suppression, we generated the inducible lines of MpNEK1 without its kinase activity (Fig. 9). For this purpose, lysine-34 essential for the kinase activity was substituted with glutamic acid (K37E, MpNEK1^K37E^) or the N-terminal kinase domain was deleted (KD deletion, MpNEK1^dKD^) (Fig. 9a). MpNEK1^K37E^ and MpNEK1^dKD^ were fused with Citrine at the C-termini and constructed into the XVE system as shown in Fig. 9a.
Effect of kinase-deficient MpNEK1 induction on the growth of thalli and rhizoids. (a) Full-length and kinase-deficient MpNEK1 used for estradiol induction. (b) The gemmae of the WT and inducible lines of full-length MpNEK1, MpNEK1K37E, or MpNEK1dKD (KD deletion) were planted in the medium with (+ER) or without 1 µM estradiol (Mock) and grown for 14 days. (c) The gemmae of the WT and inducible lines were grown as in (b) for 7 days.
We compared the effects of MpNEK1^K37E^ and MpNEK1^dKD^ induction to that of the full-length MpNEK1 induction ([Figs 9 and 10](#F9 F10), Supplementary Fig. S11). The gemmae of the WT and inducible lines of full-length MpNEK1, MpNEK1^K37E^, and MpNEK1^dKD^ were grown in the medium with (+ER) or without 1 µM estradiol (Mock). In the presence of estradiol, thallus growth was suppressed in all inducible lines (Fig. 9b). This finding indicates that the kinase activity of MpNEK1 is not required for the suppression of thallus growth. However, close-up observation showed that the thalli of MpNEK1^K37E^- and MpNEK1^dKD^-inducible lines were larger than that of full-length MpNEK1-inducible line (Fig. 9c). Furthermore, rhizoids were scarcely formed in the full-length inducible lines, whereas MpNEK1^K37E^- and MpNEK1^dKD^-inducible lines developed several rhizoids, which elongated as those of the WT (Fig. 9c).
Effect of MpNEK1 induction on the number of rhizoids. (a) The number of rhizoids in the gemmae grown for 7 days in the medium with (+ER) or without 1 µM estradiol (Mock). The gemmae of the WT and inducible lines of full-length MpNEK1–Citrine (FL), MpNEK1K37E–Citrine (K37E), and MpNEK1dKD–Citrine (dKD) were used. Data are shown by box plots (n = 10 plants). The different letters indicate significant differences by Tukey’s Honest Significant Difference (HSD) test (P < .03). Asterisks indicate significant differences from the value of FL with estradiol (+ER) (t-test, P < .01). (b) The relative number of rhizoids in (a). The average number of rhizoids in estradiol-free media (−ER) in each genotype was used for the normalization. Data are shown by box plots (n = 10 plants). The different letters indicate significant differences by Tukey’s HSD test (P < .03). Asterisks indicate significant differences from the value of FL with estradiol (+ER) (t-test, P < .01). (c) The number of rhizoids in the gemmae grown for 14 days in the medium supplemented with 1 µM estradiol. The gemmae of the inducible lines of full-length MpNEK1–Citrine (FL), MpNEK1K37E–Citrine (K37E), and MpNEK1dKD–Citrine (dKD) were used. Data are shown by box plots (n indicates the number of plants). The different letters indicate significant differences by Tukey’s HSD test (P < .03).
We quantified the number of rhizoids, which is a good indicator of the difference among the inducible lines (Fig. 10). The gemmae of the WT and inducible lines of full-length MpNEK1, MpNEK1^K37E^, and MpNEK1^dKD^ were grown in the medium with (+ER) or without 1 µM estradiol (Mock) for 7 days (Fig. 10a). The number of rhizoids in the WT was not affected by estradiol. Although the number of rhizoids was significantly decreased in all of the inducible lines, the inducible lines of MpNEK1^K37E^ and MpNEK1^dKD^ developed more rhizoids than the full-length inducible lines. This difference was confirmed in the normalized data by using the average number of rhizoids in the estradiol-free medium as a standard (Fig. 10b). The difference between kinase-deficient and full-length inducible lines was more clear in the quantification of rhizoids in 14-day treatment (Fig. 10c). Thus, the induction of kinase-deficient MpNEK1 is slightly milder than the full-length induction. The kinase activity of MpNEK1 may have a little contribution to growth suppression.
To investigate the accumulation and subcellular localization of kinase-deficient MpNEK1, inducible lines were grown in the medium with or without 1 µM estradiol and observed under confocal microscopy (Fig. 11). Because the inducible lines of kinase-deficient MpNEK1 develop larger thalli and more rhizoids than the full-length inducible lines, they provide good opportunity to analyze localization of MpNEK1 in the growing organs and more viable cells. Estradiol treatment induced accumulation of MpNEK1^K37E^ and MpNEK1^dKD^ in the granules throughout the thallus as the full-length MpNEK1 (Fig. 11a).
Localization of kinase-deficient MpNEK1–Citrine. (a) The gemmae of the inducible lines of MpNEK1K37E–Citrine (K37E) or MpNEK1dKD–Citrine (KD deletion) were planted in the medium with (+ER) or without 1 µM estradiol (Mock), grown for 4 days, and observed under a confocal microscope. Green: MpNEK1–Citrine, magenta: plastid autofluorescence. (b) The gemmae of the inducible lines of MpNEK1K37E–Citrine (K37E) or MpNEK1dKD–Citrine (KD deletion) were grown as in (a) and rhizoids were observed under a confocal microscope. Green: MpNEK1–Citrine, magenta: plastid autofluorescence. (c) Ratio of rhizoids with tip-localized or whole localized MpNEK1dKD–Citrine (KD deletion) or MpNEK1K37E–Citrine (K37E) after 120 hours of estradiol treatment. (d) Growth velocity of rhizoids in the MpNEK1K37E–Citrine-inducible lines incubated for 48 hours with (+ER) or without 1 µM estradiol (−ER). Data are shown by box plots (n = 30 rhizoids). An asterisk indicates significant difference (t-test, P < .01).
We observed localization of MpNEK1^K37E^ and MpNEK1^dKD^ in rhizoids (Fig. 11b). Some rhizoids showed granular accumulation throughout rhizoids (whole localization), while others showed preferential localization at the apex of rhizoids (tip localization). We quantified the number of rhizoids with the whole localization or tip localization (Fig. 11c). Tip-localized rhizoids were ∼47% in MpNEK1^K37E^ while 94% in MpNEK1^dKD^. Thus, MpNEK1^dKD^ exhibited relatively normal localization compared with MpNEK1^K37E^ in the inducible condition.
Growth suppression of gametangiophores in the Mpnek1 mutants. (a) Overall morphology of archegoniophores (upper panel) and antheridiophores (lower panels) of 2-month-old plants. (b) Quantification of the length of stalks of gametangiophores. Asterisks indicate significant differences from WT (t-test, P < 0.05, n = 20). (c) Longitudinal sections of stalks of archegoniophores of the WT and Mpnek1 mutant. (d) Quantification of the length of stalk cells of gametangiophores (n = 20 in female, 10 in male). (e) Promoter activity of MpNEK1 in the stalks of gametangiophores.
Next, we measured the velocity of rhizoid growth in the MpNEK1^K37E^-inducible lines (Fig. 11d). Rhizoid growth was almost stopped after 48 hours of estradiol treatment. This is comparable to the effect of the full-length inducible lines. Therefore, the excess accumulation of kinase-dead MpNEK1 also suppressed rhizoid elongation.
Mpnek1 mutants exhibit growth suppression of reproductive organs
Our gain-of-function approach suggests the involvement of MpNEK1 in cell division and proliferation. To verify this, we analyzed the phenotype of Mpnek1 mutants (Fig. 12). Because Mpnek1 mutant thalli showed a relatively mild phenotype, wavy morphology, and the same size as WT thalli (Otani et al. 2018), it is difficult to estimate the function of MpNEK1 in cell division and meristematic activity in thalli. Hence, we monitored the growth of reproductive organs, gametangiophores. Mpnek1 mutants showed obvious growth suppression of gametangiophores in both female and male plants (Fig. 12a and b). To determine whether this growth suppression is due to reduced cell elongation, we analyzed cell length in stalks (Fig. 12c and d). The length of stalk cells in Mpnek1 was almost identical to that in the WT. These results suggest that growth suppression of gametangiophores is attributed to the reduced cell division. This was supported by the preferential activity of MpNEK1 promoter in the upper region of the stalks (Fig. 12e), which is the meristematic zone of stalks.
Discussion
Here, we established and characterized the induction system of MpNEK1 as a new tool to analyze plant NEK function. Estradiol-based XVE induction system has been widely used in angiosperms, but its application and characterization in M. polymorpha still remain limited. The efficient induction of MpNEK1 transcripts and overaccumulation of MpNEK1–Citrine fusion protein throughout the plant indicate that this system is very helpful for the functional analysis of MpNEK1 and potentially for those of other proteins in M. polymorpha.
We noticed several unexpected phenomena in which caution is required. MpNEK1–Citrine showed leaky expression in the meristem without estradiol. This might be due to the basal expression of artificial transcriptional activator XVE under the control of MpEF1α promoter, which is highly active in meristematic tissues (Althoff et al. 2014). Nonetheless, this leaky expression did not affect thallus growth, suggesting that the effective threshold of NEK may exist. Since the leaky expression varied in plants and transgenic lines, estrogen-free XVE-mediated transactivation might be stochastic and/or responsive to the fluctuations of physiological states.
Second, estrogen-resistant silenced plants emerged as survivors in the prolonged treatment, especially at the lower concentrations of estradiol. This resistant phenotype could be transferred to the next-generation gemmae (e.g. DNA methylation, siRNA movement). Thus, it would be useful for the analysis of gene silencing in M. polymorpha, while invoking concerns about treatment time and estradiol concentration to minimize the contamination of silenced plants.
In summary, our results show that the short-term treatment (∼24 to 48 hours) with relatively higher concentrations of estradiol (1–10 µM) is suitable to avoid the silencing effect. Furthermore, leaky expression does not affect plant growth in our experimental condition. Nevertheless, an alternative inducible system with dexamethasone or heat treatment would be more beneficial to avoid these issues. Dexamethasone-inducible translocation system has been successfully utilized for transcriptional regulators (Yamaoka et al. 2018, Yasui et al. 2019), while a transactivation system is required for the analysis of non-nuclear proteins. Dexamethasone-inducible transactivation system (e.g. Gal4/GVG, pOp6/LhGR) is not applied in M. polymorpha and has been reported to suppress plant growth in several species (e.g. Kang et al. 1999). The heat-inducible system is also useful for gene induction and conditional knockout (Nishihama et al. 2016), while its use for gene/protein induction may not be suitable for thermolabile proteins and heat-sensitive phenotypes (e.g. Furumoto et al. 2024). Although further improvement will be required by referring to the optimization of the XVE system in a moss Physcomitrium patens (Kubo et al. 2013) and other systems, the estradiol-inducible system is very informative for functional analysis because of its less toxic effect and high induction efficiency.
MpNEK1 induction severely suppressed thallus growth probably through cell proliferation. There are several possible mechanisms of this growth-suppressing effect. First, MpNEK1 may depolymerize microtubule-based mitotic apparatus including spindle and phragmoplast or affect cortical microtubule organization, which is required for directional growth prior to cell division. However, our microtubule imaging showed that overexpression of MpNEK1 did not severely affect the mitotic apparatus. During the interphase, MpNEK1 reduced the density and dynamics of cortical microtubules but did not severely affect cell growth and morphology. MpNEK1 may depolymerize cortical microtubules but its effect is under the threshold and/or masked by other regulators. Second, overexpression of MpNEK1 may activate cell cycle checkpoints to prevent cell cycle progression. According to the experiment of EdU labeling, the cells through the S phase were reduced under the MpNEK1 overexpression. Furthermore, MpNEK1 induction reduced mitotic cells in the M phase. Thus, MpNEK1 induction may stop the cell cycle in the G1 phase or G1/S transition to eventually reduce the number of dividing cells. The nimA mutants of a filamentous fungus Aspergillus nidulans exhibit cell cycle arrest just before the M phase, and conversely, overexpression of NimA causes premature chromosome condensation and aberrant spindle formation, resulting in cell cycle arrest (Oakley and Morris 1983, Osmani et al. 1988). Taking into account of remarkable promoter activity of MpNEK1 in the meristem (Otani et al. 2018), it would be likely that MpNEK1 controls cell cycle progression and checkpoints.
Overexpression of MpNEK1 suppressed tip growth in rhizoids. Furthermore, overexpression altered localization of MpNEK1–Citrine from the rhizoid apex to the entire region as the time of estradiol treatment increased. This may reflect the overflow of MpNEK1 from the transport and localization machineries. As a consequence, tip growth was stopped in rhizoids with this dispersed localization pattern. Over-accumulated MpNEK1 may affect microtubule organization and/or microtubule-dependent processes during rhizoid growth. Because the longitudinal microtubules promote rhizoid elongation through organelle transport (Kanda et al. 2022), it is plausible that MpNEK1 accumulation throughout rhizoids suppresses the transport system (see the last paragraph).
Unexpectedly, we found growth suppression of thalli and rhizoids by kinase-deficient MpNEK1. This result demonstrates that growth suppression is mainly independent of protein phosphorylation by MpNEK1. Therefore, the induction system is not suitable for the identification of target proteins phosphorylated by MpNEK1. Nevertheless, kinase-dead MpNEK1 is slightly milder than the WT MpNEK1 in the estradiol induction system. The kinase activity of MpNEK1 is not essential for growth suppression but may have a minor contribution. The most plausible scenario is that excessively accumulated MpNEK1 might trap interacting proteins and substrates in the granules to suppress thallus growth and rhizoid elongation in a kinase-independent manner. This is supported by the findings that the C-terminal regulatory regions of plant NEKs interact with various proteins (Pnueli et al. 2001, Sakai et al. 2008, Motose et al. 2011). Animal NEK7 licenses the assembly and activation of NLRP3 inflammasome, a large protein complex for the activation of inflammatory caspases, independently of its kinase activity (He et al. 2016, Sharif et al. 2019). Furthermore, NEK7 is required for the oligomerization of NLRP3 and its adaptor protein ASC, which forms the granular structure in the cytosol (He et al. 2016, Sharif et al. 2019). Plant NEK may also function as the assembly factor of protein supercomplex and cytosolic granular structures.
In summary, the MpNEK1 induction system is useful for the analyses of its function in cell division and proliferation. Furthermore, it would be helpful to visualize directional protein transport in rhizoids because we observed MpNEK1 particles moved to the acropetal and basipetal direction (Videos S3–S5). The velocity of MpNEK1 particles is quite similar to that of armadillo-repeat kinesin (Kanda et al. 2022), about half of velocity of growing microtubule plus ends, suggesting this motility is not due to plus-end tracking but rather to the kinesin-dependent transport along microtubules. MpNEK1 overexpression has the potent growth effect than that of Arabidopsis NEK6, whose overexpression caused a slight reduction of root growth (Takatani et al. 2017). Compared with MpNEK1, NEK6–Citrine fusion protein is not highly accumulated in the XVE system (H.Mo., unpublished result), implying the differential degradation efficiency of NEK proteins and the potential advantage of protein induction and analyses in M. polymorpha.
Materials and Methods
Plant materials and growth conditions
M. polymorpha accessions Takaragaike-1 (Tak-1, male) and Takaragaike-2 (Tak-2, female) were used as the WT strains. Mpnek1 mutants and MpNEK1pro:GUS lines were reported in Otani et al. (2018). Plants were grown on the half-strength Gamborg’s B5 medium solidified with 1% agar at 22°C under the light cycles with 16 hours of light and 8 hours of darkness.
Plasmid construction and transformation
Plasmids were mainly constructed based on the Gateway Technology (Life technologies). Primers used in the plasmid construction are listed in Table S1. To generate a plasmid of MpNEK1 induction by estradiol, the full-length MpNEK1 coding sequence in the pENTR/D-TOPO entry vector (Otani et al. 2018) was transferred by LR reaction (LR clonase II enzyme mix, Life Technologies) into the Gateway binary vector pMpGWB344 (MpEF1α pro:XVE≫LexA operator:Gateway cassette).
To generate a plasmid of estradiol-inducible MpNEK1-mCitrine, mCitrine coding sequence was amplified by KOD One (Toyobo) and a primer set (Table S1) and was cloned into the AscI site of the pENTR/D-TOPO entry vector containing full-length MpNEK1 CDS (Otani et al. 2018) by In-fusion system (Takara). The resulting vector pENTR/D–TOPO–MpNEK1–mCitrine was subjected to LR reaction to transfer MpNEK1–Citrine fusion into the Gateway binary vector pMpGWB144 (MpEF1α pro:XVE≫LexA operator:Gateway cassette).
To generate a plasmid of estradiol-inducible kinase-deficient MpNEK1, pENTR/D–TOPO–MpNEK1–mCitrine was subjected to the inverse PCR-based mutagenesis using KOD mutagenesis kit (Toyobo) and primer sets (Table S1). The resulting vectors, pENTR/D–TOPO–MpNEK1^K37E^–mCitrine and pENTR/D–TOPO–MpNEK1^dKD^–mCitrine were subjected to LR reaction to transfer mutated MpNEK1–Citrine fusion into the Gateway binary vector pMpGWB144 (MpEF1α pro:XVE≫LexA operator:Gateway cassette).
The constructs, pMpGWB344–MpNEK1 (MpEF1α pro:XVE≫LexA operator:MpNEK1), pMpGWB144–MpNEK1–Citrine (MpEF1α pro:XVE≫LexA operator:MpNEK1–Citrine), pMpGWB144–MpNEK1^K37E^–Citrine (MpEF1α pro:XVE≫LexA operator:MpNEK1^K37E^–Citrine), and pMpGWB144–MpNEK1^dKD^–Citrine (MpEF1α pro:XVE≫LexA operator:MpNEK1^dKD^–Citrine) were transformed into F1 sporelings derived from sexual crosses between Tak-2 and Tak-1 by Agrobacterium-mediated method (Ishizaki et al. 2008). The pMpGWB344–MpNEK1 construct was transformed into the regenerating thalli of CaMV35Spro:Citrine–MpTUB2 plants harboring pMpGWB105–MpTUB2 construct (Otani et al. 2018) according to the method of Kubota et al. (2013). Chlorsulfuron and hygromycin were used for the selection of transformants in pMpGWB344 and pMpGWB144, respectively. In pMpGWB344 and pMpGWB144, the artificial transcriptional activator XVE is expressed under the control of MpEF1α promoter. XVE translocates to the nucleus in the presence of estradiol and binds to the LexA operator to induce the expression of MpNEK1– and MpNEK1–Citrine, respectively.
Quantitative real-time PCR
Total RNA was isolated from the thalli of the WT and transgenic plants. For each sample, 0.5 µg of total RNA was reverse transcribed to cDNA using ReverTra Ace reverse transcriptase (Toyobo) according to the manufacturer’s protocol. Real-time PCR was performed on a thermal cycler Dice Real Time System (Takara) using KOD SYBR qPCR Kit (Toyobo) according to the manufacturer’s protocol. Transcript levels of MpEF1α and MpACT were used as a reference for normalization (Saint-Marcoux et al. 2015). Primers used in RT-qPCR are listed in Table S1. RT-qPCR experiments were performed using four biological replicates.
Microscopy
The morphology of gemmalings and rhizoids was observed under a stereo microscope S8APO0 (Leica Microsystems) or a light microscope DM5000B (Leica) equipped with a Charge Coupled Device (CCD) camera DFC500 (Leica). The morphology of gametangiophores was photographed by a single-lens reflex camera D5600 (Nikon, Tokyo Japan). Beta-glucuronidase (GUS) staining was conducted according to Otani et al. (2018) and observed under DM5000B equipped with DFC500. To analyze the localization of MpNEK1–Citrine, plants were grown on half-strength B5 medium for 3 and 5 days under cycles with 16 hours of light and 8 hours of darkness. For live imaging, half-strength B5 agar medium was solidified in the glass bottom dish (35 mm in diameter × 10 mm in thickness, Thickness No. 1, Matsunami) and then the central region of agar medium on the bottom glass strip was removed by tweezers. This region was poured with ∼200 µl of melted half-strength B5 agar medium to generate a thin solidified medium. The gemmae were planted and grown in the central thin medium for 2–7 days. The details of this method will be described elsewhere by H.M. Gemmalings were observed under a FV1200 confocal laser-scanning microscope (Olympus) equipped with a high-sensitivity GaAsP detector and silicone oil objective lenses (30×, 60× Olympus) or FV3000 (Olympus) equipped with high-sensitivity GaAsP detectors and oil immersion objective lenses (40×, NA = 1.4; 60×, NA = 1.42, Olympus) or water immersion objective lenses (60×, NA = 1.2, Olympus). Silicone oil (SIL300CS, Olympus), immersion oil (F30CC, Olympus), or deionized water was used as immersion media for these objective lenses. The samples were excited at 473 and 559 nm (laser diode) and the emission was separated using a FV12-MHSY SDM560 filter (490–540 and 575–675 nm, Olympus) in FV1200. The samples were excited at 488 and 561 nm (laser diode) and the emission was separated using the multichannel TruSpectral detection (500–540 and 600–700 nm, Olympus) in FV3000. The images were analyzed using ImageJ (National Institutes of Health, USA) and LPIXEL ImageJ Plugins (https://lpixel.net/products/lpixel-imagej-plugins/). Microtubule bundling (skewness) and density (occupancy) were determined according to Higaki et al. (2010). Microtubule dynamics was quantified according to Abe et al. (2005). For estradiol treatment, an aliquot (200–300 µl) of 1/2 B5 liquid medium supplemented with estradiol at a concentration of 1 µM or 10 µM was added to the gemmalings for the induction of MpNEK1. As a control, the same amount of 1/2 B5 liquid medium supplemented with the same concentration of solvent (dimethyl sulfoxide, <0.1%) was applied.
EdU staining
S-phase cells were visualized using Click-iT EdU Imaging Kits (Life Technologies) according to the manufacturer’s instructions. Gemmae were incubated on the half-strength Gamborg’s B5 agar medium supplemented with or without 1 µM β-estradiol for 4 days. Alternatively, for 1-day treatment, gemmae were incubated on the agar medium without 1 µM β-estradiol for 3 days and then transferred to and incubated in the agar medium with 1 µM β-estradiol for 1 day. Gemmalings were then transferred to a solution containing 10 µM EdU and incubated for 1 hour. Plants were fixed with 3.7% formaldehyde solution in phosphate-buffered saline for 20 minutes. EdU incorporated into DNA was labeled with Alexa Fluor 488-azide-containing Click-iT reaction cocktail in the dark for 30 minutes. EdU-labeled cells were observed using a confocal laser scanning microscope, FV1200 (Olympus). The maximum Z-projection images were created using the ImageJ software.
Supplementary Material
pcaf021_Supp
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abe T. and Hashimoto T. (2005) Altered microtubule dynamics by expression of modified alpha-tubulin protein causes right-handed helical growth in transgenic Arabidopsis plants. Plant J. 43: 191–204.15998306 10.1111/j.1365-313X.2005.02442.x · doi ↗ · pubmed ↗
- 2Althoff F., Kopischke S., Zobell O., Ide K., Ishizaki K., Kohchi T., et al. (2014) Comparison of the Mp EF 1α and Ca MV 35 promoters for application in Marchantia polymorpha overexpression studies. Transgenic Res. 23: 235–244.24036909 10.1007/s 11248-013-9746-z · doi ↗ · pubmed ↗
- 3Bao H., Sun R., Iwano M., Yoshitake Y., Aki S.S., Umeda M., et al. (2024) Conserved CKI 1-mediated signaling is required for female germline specification in Marchantia polymorpha. Curr. Biol. 34: 1324–1332.38295795 10.1016/j.cub.2024.01.013 · doi ↗ · pubmed ↗
- 4Bowman J.L., Kohchi T., Yamato K.T., Jenkins J., Shu S., Ishizaki K., et al. (2017) Insights into land plant evolution garnered from the Marchantia polymorpha genome. Cell 171: 287–304.28985561 10.1016/j.cell.2017.09.030 · doi ↗ · pubmed ↗
- 5Buschmann H., Holtmannspötter M., Borchers A., O’Donoghue M.T. and Zachgo S. (2016) Microtubule dynamics of the centrosome-like polar organizers from the basal land plant Marchantia polymorpha. New Phytol. 209: 999–1013.26467050 10.1111/nph.13691 · doi ↗ · pubmed ↗
- 6Champion C., Lamers J., Jones V.A.S., Morieri G., Honkanen S. and Dolan L. (2021) Microtubule associated protein WAVE DAMPENED 2-LIKE (WDL) controls microtubule bundling and the stability of the site of tip-growth in Marchantia polymorpha rhizoids. P Lo S Genet. 17: e 1009533.10.1371/journal.pgen.1009533 PMC 817753434086675 · doi ↗ · pubmed ↗
- 7Flores-Sandoval E., Dierschke T., Fisher T.J. and Bowman J.L. (2016) Efficient and inducible use of artificial micro RN As in Marchantia polymorpha. Plant Cell Physiol. 57: 281–290.25971256 10.1093/pcp/pcv 068 · doi ↗ · pubmed ↗
- 8Fry A.M., O’Regan L., Sabir S.R. and Bayliss R. (2012) Cell cycle regulation by the NEK family of protein kinases. J. Cell Sci. 125: 4423–4433.23132929 10.1242/jcs.111195 PMC 3500863 · doi ↗ · pubmed ↗