Iron-restricted Mycobacterium tuberculosis exports pathogenicity factors packed in extracellular vesicles
Nishant Sharma, Nevadita Sharma, Ashis Biswas, Shamba Gupta, Assirbad Behura, Gloria Marcela Rodriguez

TL;DR
This study shows that tuberculosis bacteria release vesicles containing harmful proteins when iron is scarce, which could help develop new TB diagnostics.
Contribution
The novel finding is that iron limitation triggers M. tuberculosis to export pathogenicity factors via extracellular vesicles.
Findings
M. tuberculosis increases extracellular vesicle production under iron limitation.
Vesicles contain proteins that enhance bacterial survival and immune evasion.
The findings suggest vesicles could serve as TB biomarkers and diagnostic tools.
Abstract
Mycobacterium tuberculosis, the pathogen responsible for human tuberculosis, responds to iron limitation by increasing the production of extracellular vesicles. This study examined the protein composition of induced M. tuberculosis extracellular membrane vesicles using chromatography coupled with mass spectrometry. The results revealed that vesicles contain key pathogenicity factors, including proteins that enhance bacterial survival, immune evasion, and inflammation. These findings deepen our understanding of the potential role of extracellular vesicles in M. tuberculosis-host interactions. The data can also aid in identifying new biomarkers of infection and developing vesicle-based, culture-independent TB diagnostic platforms.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
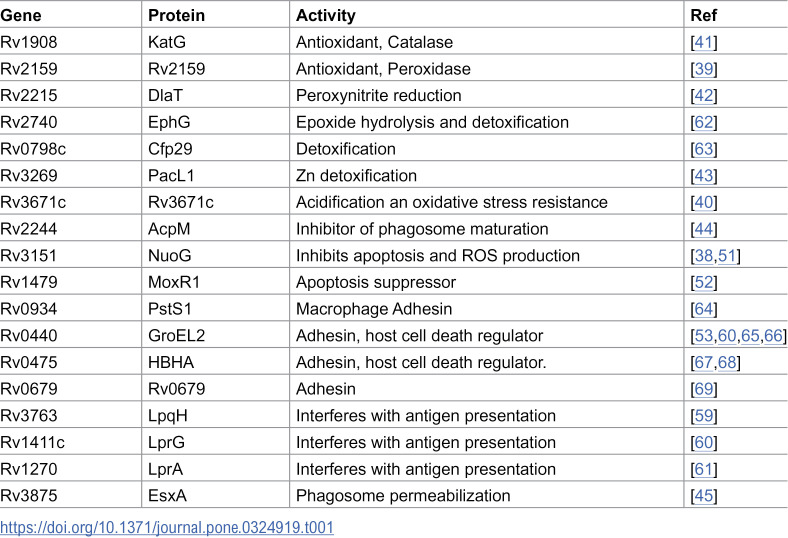 Figure 1
Figure 1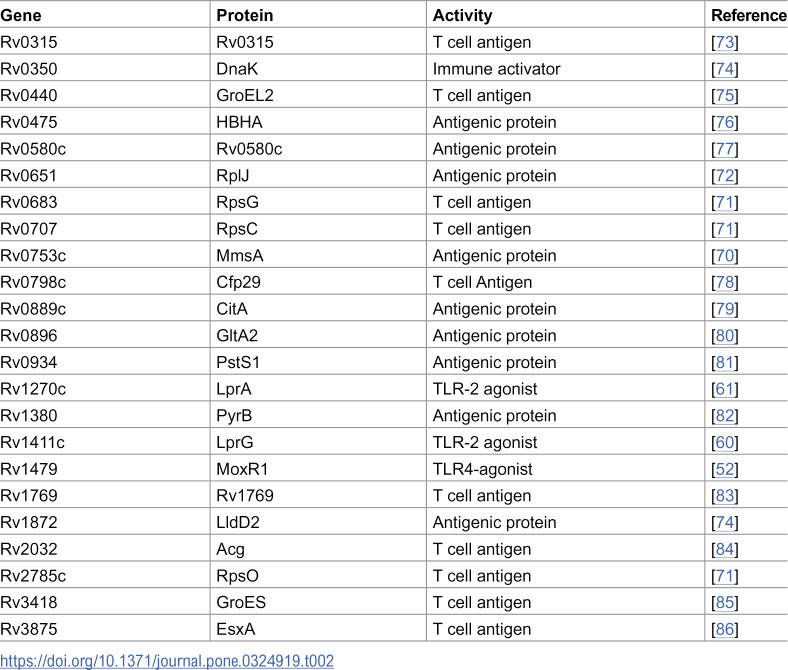 Figure 2
Figure 2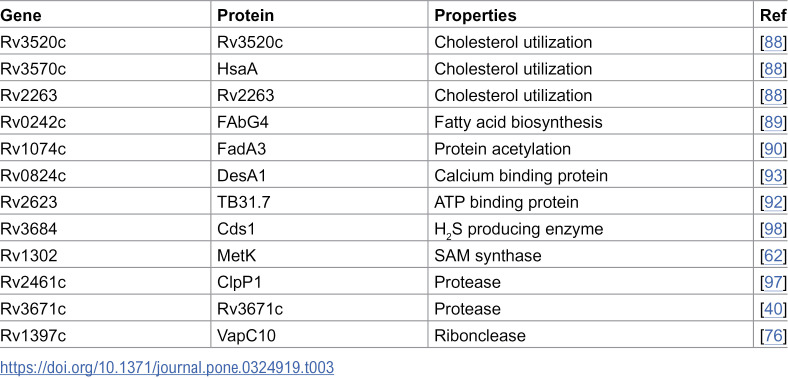 Figure 3
Figure 3- —http://dx.doi.org/10.13039/100015691Division of Microbiology and Infectious Diseases, National Institute of Allergy and Infectious Diseases
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial Infections and Vaccines · Extracellular vesicles in disease · Pneumocystis jirovecii pneumonia detection and treatment
Introduction
Mycobacterium tuberculosis (Mtb) is an ancient pathogen that continues to cause significant morbidity and mortality worldwide. Mtb is responsible for over one million deaths due to tuberculosis (TB) each year. However, our understanding of its virulence strategies remains incomplete. Although immunocompromised individuals are at greater risk of developing TB disease, most people with subclinical TB are immunocompetent, highlighting Mtb’s ability to persist despite an active immune response [1]. Understanding the interactions between the pathogen and the host that undermine effective immunity is essential for developing urgently needed vaccines, innovative therapies targeted at either the pathogen or the host, and diagnostic tools to help control the ongoing TB pandemic.
A central feature of Mtb’s success as a pathogen is its remarkable ability to modify its environment, creating an intracellular niche and evading and subverting the macrophage’s antimicrobial defenses and immune functions; for this, Mtb is known to employ cell envelope glycolipids and secreted proteins. Furthermore, Mtb secretes extracellular vesicles, enabling distal communication with uninfected host cells [2–4].
Extracellular vesicles (EVs) are membrane-bound nanoparticles that encapsulate and transport a variety of biomolecules, including proteins, lipids, nucleic acids, and metabolites. Bacteria ubiquitously produce EVs, and this process is genetically regulated [5–8]. The composition of bacterial EVs is specific to different species and influenced by environmental factors [9,10]. Growing evidence indicates that EVs produced by symbiotic bacteria can promote beneficial health effects for the host [11,12], contrasting with the selective export of toxins and other virulence factors via EVs observed in many pathogenic bacteria [13–16]. EV production likely aids pathogens by enabling the simultaneous export, transport, and delivery of multiple effectors into host cells in a concentrated and protected manner [17].
Mycobacterial EVs originate from the cytoplasmic membrane and range in size from 60 to 300 nm in diameter [18]. We previously demonstrated that Mtb produces abundant vesicles when faced with iron restriction, a well-known stress condition prevalent in the host environment and intensified by nutritional immunity. Mtb generates very few vesicles under conditions of iron sufficiency [19]. Iron restriction also triggers the expression of many virulence-associated genes [20]. The hydrophobic siderophore and virulence factor, mycobactin [21], produced in response to iron limitation [22,23], is packed and exported in low-iron-induced EVs (hereafter referred to as MEVs) [19], which can support the growth of a siderophore biosynthesis mutant in low-iron medium, indicating a role for MEVs in iron acquisition [19].
Recently, we demonstrated that the upregulation of mycobacterial vesicle production is linked to enhanced expression of the iron-regulated dynamin-like proteins IniA and IniC, which are required for vesicle biogenesis [24]. Utilizing an Mtb iniA deletion mutant, we showed that the intracellular packing and subsequent export of bacterial antigens from infected macrophages within EVs enable intracellular Mtb to communicate with bystander host cells [24–26].
To improve understanding of MEV functionality during infection, we characterize the protein composition of MEVs using a quantitative proteomic approach. The results offer valuable insights into the potential role of MEV cargo in TB pathogenesis and establish a foundation for new research on vesicle-associated TB biomarkers.
Materials and methods
Culture conditions
Mtb H37Rv (ATCC) was recovered from a frozen stock on Middlebrook 7H10 agar supplemented with 0.2% glycerol, 0.5% bovine serum albumin fraction V, 0.2% dextrose, 0.085% NaCl, and 0.05% Tween-80. To induce MEV production, Mtb grown to logarithmic phase on 7H10 was inoculated into 1 L of minimal medium (MM) [0.1% asparagine, 0.05% KH_2_PO_4_, 0.25% Na_2_HPO_4_, 0.2% glycerol, 0.2% dextrose, and 0.085% NaCl (pH 6.8)] at an optical density (O.D_600_) of 0.1 and cultured at 37°C until reaching an O.D_600_ of 0.8. Before use, MM was treated with 5% Chelex-100 (BioRad) for 24 hours to remove traces of iron. Chelex was subsequently removed by filtration through a 0.22 μm filter (Millipore), and the medium was supplemented with 0.5 mg of ZnCl_2_ per liter, 0.1 mg of MnSO_4_ per liter, and 40 mg of MgSO_4_ per liter. This medium contains approximately 1 µM residual iron as determined by atomic absorption spectrometry. Bacterial growth was monitored based on the increase in O.D.600 measured in an aliquot removed from the primary culture and supplemented with 0.05% Tween-80 to prevent bacterial clumping.
MEV isolation
MEVs were isolated and purified as previously described [27]. Briefly, Mtb cultures were centrifuged at 3,900 × g for 15 minutes at 4°C. The culture supernatant was collected for vesicle isolation, while the bacterial cell pellet was processed to generate whole-cell protein extracts. The culture supernatant was sterilized by filtration through a 0.22 µm filter and concentrated to 25 ml using an Amicon Ultrafiltration system (Millipore) equipped with a 100-KDa exclusion membrane. The concentrate was centrifuged at 15,000 ×* *g for 15 minutes at 4°C to remove protein aggregates. MEVs were separated by ultracentrifugation at 100,000 × g for 2 hours at 4°C. MEVs were adjusted to 45% w/v iodixanol (OptiPrep, Sigma-Aldrich) by adding OptiPrep in PBS buffer to a total of 1 ml, then loaded into the bottom of a 12.5 mL ultracentrifuge tube (Beckman) and layered with decreasing percentages of OptiPrep/PBS (1 ml of a 40% concentration, 1 ml of 35%, 1 ml of 30%, 1 ml of 25%, 1 ml of 20%, and 1 ml of PBS). Samples were centrifuged (100,000 × g, 18 h, 4°C), and fractions were removed sequentially from the top of the gradient. The iodixanol was removed by dilution in PBS (25 ml) followed by centrifugation (100,000 × g, 1 hour, 4°C). Previously, we established that membrane vesicles separate in the Optiprep 30−25% fraction, as observed through visualization by electron microscopy, immunological detection of antigenic components, nanoparticle analysis, and estimation of vesicle-associated proteins and lipids [19,27]. MEVs isolated from three independent cultures were analyzed by LC-MS/MS.
Generation of whole-cell protein extracts
The cell pellet collected from MM cultures was resuspended in 20 ml of cold PBS along with a protease inhibitor cocktail (Roche). The cells were lysed using two 1-minute pulses of bead beating (FastPrep-24 instrument, MP Biomedicals, Santa Ana, CA) with ice cooling between pulses. The lysates were centrifuged at 10,000 x g at 4°C to eliminate cell debris, and the supernatant was collected and filtered through a 0.22 µm filter.
Proteomic analysis
Sample preparation.
Five micrograms of each sample were reduced and denatured by adding 10 mM DTT and heating at 60^°^C for 30 minutes. Next, the samples were alkylated with 20 mM iodoacetamide for 45 minutes at room temperature in the dark. Sequencing grade trypsin (0.2 µg, Thermo Scientific) was added, and the proteins were digested at 37^°^C overnight. Peptides were extracted twice with 5% (v/v) formic acid and 60% (v/v) acetonitrile and then dried under vacuum.
Liquid chromatography-tandem mass spectrometry (LC-MS/MS).
Samples were analyzed by LC-MS using Nano LC-MS/MS (Dionex Ultimate 3000 RLSCnano System, interfaced with Eclipse (ThermoFisher).
Three microliters of a 12.5 µl peptide digest per sample were loaded onto a fused silica trap column (Acclaim PepMap 100, 75 µm x 2 cm, ThermoFisher). After washing for 5 minutes at a flow rate of 5 µl/min with 0.1% TFA, the trap column was connected in line with an analytical column (Nanoease MZ peptide BEH C18, 130A, 1.7 µm, 75 µm x 250 mm, Waters) for LC-MS/MS. Peptides were fractionated at 300 nL/min using a segmented linear gradient of 4–15% B over 30 minutes (where A is 0.2% formic acid, and B is 0.16% formic acid, 80% acetonitrile), followed by 15–25% B over 40 minutes, 25–50% B over 44 minutes, and then 50–90% B over 11 minutes. Solution B returns to 4% for 5 minutes for the next run.
Data collection and analysis.
The eluted peptides were analyzed using the DIA (Data Independent Acquisition) workflow. The MS scan range was set from 400 to 1200, the resolution was 12,000, AGC was configured to 3E6, and the ion time was set to auto. An 8 m/z window was used to sequentially isolate (AGC 4E5 and ion time set to auto) and fragment the ions in a C-trap with a relative collision energy of 30. The MS/MS data were recorded at a resolution of 30,000.
Two samples (EV3 and CP3) were also analyzed using DDA (data-dependent acquisition) to create specialized libraries for MaxQuant analysis of DIA data. The DDA scan sequence commenced with an MS1 spectrum (Orbitrap analysis, resolution 120,000, scan range from M/Z 375–1500, automatic gain control (AGC) target 1E6, maximum injection time 100 ms). The top S (3 sec) duty cycle scheme was employed to determine the number of MS/MS events performed during each cycle. Parent ions with charges between 2 and 7 were selected for MS/MS, and a dynamic exclusion of 60 seconds was applied to prevent repeat sampling. Parent masses were isolated in the quadrupole with an isolation window of 1.2 m/z, AGC target of 1E5, and fragmented using higher-energy collisional dissociation with a normalized collision energy of 30%. The fragments were scanned in Orbitrap at a resolution of 15,000. The MS/MS scan ranges were established based on the charge state of the parent ion, with the lower limit set at 110 amu.
The DDA raw mass spectrometry data were processed with MaxQuant (version 2.4.10.0) [28] and searched against the Mycobacterium tuberculosis H37Rv reference proteome downloaded from UniProt using the standard label-free workflow. The search parameters were as follows: carbamidomethyl (C) (fixed), acetyl (N-term), and oxidation (M) (variable) modifications were applied. For the full-scan MS spectra (MS1), the mass error tolerance was set to 20 ppm, and for the MS/MS spectra (MS2) to 20 ppm. Trypsin was selected as a protease with a maximum of two missed cleavages. For protein identification, a minimum of one unique peptide with a peptide length of at least seven amino acids and a false discovery rate below 0.01 was required on the peptide and protein levels. The DIA data were analyzed with MaxQuant again using the MaxDIA workflow [29] with the DDA evidence.txt as a library. The match between runs function was enabled, and a time window of one minute was set. Label-free quantification was selected using iBAQ (calculated as the sum of the intensities of the identified peptides and divided by the number of observable peptides of a protein) [30].
Statistical analysis.
The average iBAQ value of both MEVs and cell pellets (CP) was used to calculate the fold enrichment as the ratio of iBAQ in MEVs to iBAQ in CPs; n = 3. For the fold comparison, an unpaired Student’s t-test was calculated using GraphPad Prism 9.0 software. A t-test value of 0.01 was considered statistically significant.
Results
Proteomic analysis of MEVs
To investigate MEV composition, we employed mass spectrometry and identified proteins selectively packed into vesicles by Mtb H37Rv under iron-limited conditions. The cell pellets from three independent low-iron cultures in the logarithmic phase were lysed to generate whole-cell protein extracts, while the culture supernatants were processed for MEV isolation. MEVs were isolated, purified, and validated as previously [19,24,27] (S1 Fig). The whole-cell protein extracts and MEV preparations underwent proteomic analysis using tryptic digestion followed by LC-MS/MS. Proteins with at least five identified peptides in MEVs (totaling 2,192) are detailed in the S1 Data Set. The relative abundance of each protein in each preparation was determined based on the quantitative MS intensities (Intensity Absolute Quantification or iBAQ), calculated as the sum of the intensities of the identified peptides divided by the number of observable peptides. The most abundant proteins (iBAQ ≥ 0.1%) (S2 Data Set) were selected for further analysis and annotation (S2 Fig). As expected for membrane EVs, this group of proteins predominantly consists of proteins previously identified in proteomic analyses of Mtb membrane fractions (97.5%) (S3 Fig) [31–33], and a substantial proportion of them (39.5%) had been detected in Mtb culture filtrates previously [32,34,35]. A total of 85 proteins showed higher abundance (>1.0-fold) in MEVs compared to cell pellets; for 32 of these, the difference reached statistical significance (S1 Table), indicating selective packaging into MEVs. A brief review revealed no apparent commonalities among the proteins enriched in MEVs. They display a wide range of molecular weights (6.8–176.9 KDa), isoelectric points (4.7–11.5), and lengths (61–456 amino acids), belonging to diverse protein families. Notably, lipoproteins comprised this group’s largest single protein family (18.7%) (S1 Table). We questioned whether the observed preference for incorporating these proteins into MEVs correlated with increased gene expression under iron limitation. To investigate this question, we analyzed published data on the dynamics of iron-regulated gene expression in Mtb [23,36]. Among the proteins enriched in MEVs, only PfkA is upregulated under iron limitation, indicating that selective packaging into MEVs is not solely dictated by gene expression.
According to the broad functional categorization in the mycobacterial database, Mycobrowser (https://mycobrowser.epfl.ch/genes), MEVs contain proteins from various categories: conserved hypothetical proteins (14.2%), proteins predicted to be involved in intermediate metabolism and respiration (41.4%), lipid metabolism (14.8%), cell wall and cell processes (11.7%), information pathways (11.1%), and virulence, detoxification, and adaptation (5.6%) (S4 Fig). However, a comprehensive review of recent literature revealed that MEVs pack numerous housekeeping proteins that moonlight as adaptation and pathogenicity effectors; based on these findings, we categorized and discussed the most abundant proteins in MEVs according to their potential functionality at the pathogen-host interface.
MEV-associated proteins can promote bacterial survival
Mtb survives in the host by subverting lung macrophage antimicrobial defenses, including acidification, iron restriction, intoxication with heavy metals, antimicrobial peptides, reactive oxygen species (ROS), reactive nitrogen species (RNS), and lysosomal degradation [37]. Our proteomic analysis identified proteins in MEVs that can protect Mtb against ROS (NuoG, Rv2159c, Rv3671c, and KatG) [38–41], RNS (DlaT) [42], and toxic zinc accumulation (PacL) [43]. Additionally, MEVs contain AcpM, which inhibits phagosome maturation [44], and EsxA, a virulence effector that promotes phagosome membrane permeabilization and bacterial escape into the cytoplasm [45] (Table 1).
Table 1: MEV-associated proteins facilitate immune evasion.
Mtb-infected macrophages initiate programmed cell death (apoptosis) as a last resort to eliminate intracellular infection [46]. In apoptosis, degraded cellular contents remain within membrane blebs. Subsequent efferocytosis of apoptotic cells eliminates Mtb and protects the host [47]. Virulent Mtb inhibits apoptosis and promotes necrotic host cell death, leading to tissue pathology and facilitating bacterial dissemination and transmission [48–50]. MEVs carry multiple proteins that regulate host cell death, including the apoptosis inhibitors NuoG [51], MoxR1 [52], and GroEL2 [53]. By inhibiting apoptosis in infected macrophages and neutrophils, these proteins delay antigen acquisition by dendritic cells and trafficking to the lymph nodes, thus hindering effective communication between the innate and adaptive immune systems. Apoptogenic proteins such as LpqH [54,55], EsxA [56], and PstS-1 [57], along with anti-autophagy factors like MoxR1, KatG, and the heparin-binding hemagglutinin protein (HBHA), were also identified in MEVs [47,58] (Table 1).
Some MEV-associated proteins help Mtb evade immune surveillance by directly interfering with antigen-presenting cell function and T-cell activation. For instance, the chaperone GroEL2 inhibits dendritic cell maturation, and the lipoproteins LpqH, LprG, and LprA have been demonstrated to inhibit macrophage IFN-induced MHC-II expression and antigen presentation to CD4^+^ T cells [59–61] (Table 1).
MEV-associated proteins provoke the immune system
Evidence from various sources links mycobacterial EVs to inflammation. A mutant of Mtb (virR::Tn), which exhibits enhanced vesicle release, induced significantly more inflammation in infected mice than the parental strain [8]. Isolated vesicles derived from Mtb cultures injected intratracheally into mice triggered robust inflammation and exacerbated a subsequent infection with virulent Mtb [18]. Furthermore, EVs released by macrophages infected with an iniA mutant deficient in vesicle production exhibited a reduced capacity to induce proinflammatory cytokine release by uninfected macrophages compared to EVs released by wild-type Mtb-infected macrophages [21]. Consistent with the pro-inflammatory properties of MEVs, the data revealed that MEVs transport numerous immunostimulatory proteins, including multiple Toll-Like Receptor-2 (TLR2) agonists (LpqH, LprG, LprA, EsxA) and recognized immunodominant T cell antigens (MmsA, GroEL2, DnaK, Cfp29, RpIJ, RpsO, RpsG, and RpsC) [70–72] (Table 2).
Table 2: MEV-associated immunostimulatory proteins.
MEV-associated proteins could intersect host metabolism and signaling pathways
MEVs contain various enzymes that can target host substrates. For instance, it is well known that Mtb utilizes host lipids to persist in infected macrophages, and it can suppress host immunity by modulating host fatty acid metabolism [87]. In this regard, intriguingly, enzymes involved in cholesterol catabolism (HsaA, Rv3520c, and Rv2263) [88] are included in MEVs alongside FabG4, a protein shown to mediate host fatty acid synthesis when expressed in eukaryotic cells [89] and FadA3, which mediates differential acetylation of host proteins in Mtb infected macrophages in association with increased bacterial survival in mice [90].
MEVs also contain SerC, an enzyme that enables the production of D-serine, a metabolite that interacts with WDR24 and inhibits mTORC1 activation in CD8^+^ T cells. This action indirectly affects macrophages’ ability to restrict Mtb replication [91]. In addition, the ATP-binding protein encoded by Rv2623 [92] and the calcium-binding proteins DesA1 [93] and SecE2 [94] can alter host cell signaling pathways. Importantly, calcium-dependent signaling in immune cells affects phagosome maturation, apoptosis, and cytokine release [95,96].
The proteases ClpP1 [97] and Rv3671c [40] may alter host cell proteostasis. At the same time, Cds1, a cysteine desulfurase, can generate H_2_S, a potent gasotransmitter linked to enhanced Mtb pathogenesis, suppression of host immunity, and exacerbation of TB in mice [98,99]. Further, the ribonuclease VapC10 may target host cell RNA and alter gene expression (Table 3) [100].
Table 3: MEV proteins that can intersect host metabolism and signaling.
MEVs transport host protein-interacting partners
Some proteins identified in MEVs have been shown to interact directly with host cell proteins. These include the adhesins GroEL2 and HBHA, which bind to CD43 on macrophages [65,66], and Syndecan 4 in epithelial cells, respectively. The interaction between HBHA and epithelial cells is crucial for the extrapulmonary dissemination of Mtb [67,68]. LpqE, SahH, and Gap bind to ubiquitin 1, IL-8, and transferrin, respectively [101–103]. LpqE, LpqH, LpqN, Rv2314c, and Rv0954 were identified in a study of the host-pathogen interactome that focused on Mtb-secreted proteins [104]. The data from this study indicated that these proteins could interact with multiple host proteins. The interaction between LpqN and C3 ubiquitin ligase was validated and found essential for Mtb virulence in macrophages [104].
The potential of MEVs as a source of novel TB biomarkers
As nutritional immunity restricts iron from invading pathogens, we anticipated that the cargo of MEVs would reflect that of vesicles released during infection. We reviewed the data reported by Kruh-Garcia et al. to investigate this hypothesis [105]. These authors employed targeted MRM-MS to detect mycobacterial proteins in EVs isolated from the serum of individuals with active TB. They successfully identified 17 mycobacterial proteins with high confidence. MEVs contain all of these proteins (S3 Data Set), reinforcing the close relationship between the protein cargo of MEVs and that of EVs produced during infection.
Discussion
Iron is an essential metal for Mtb survival. However, its bioavailability within the host is usually tightly regulated and further limited by nutritional immunity [106]. Our previous studies established that Mtb upregulates EV biogenesis in response to iron restriction and that very few vesicles are released during logarithmic growth in a high-iron medium [19]. In this study, we examined induced EVs (MEVs) protein components. Overall, the proteome of MEVs can be functionally linked to the distinct types of Mtb interactions with the immune system: evasion of the innate immune response, immune subversion, and stimulation of a robust inflammatory response and tissue pathology needed for transmission [2]. Thus, based on the protein cargo of MEVs, we postulate that EV production may aid Mtb in evading immune-mediated killing while exploiting the host inflammatory response to continue its life cycle. Importantly, MEVs contained Mtb proteins associated with EVs released in humans infected with Mtb. This supports the significance of MEVs as a model for EVs produced in vivo and, therefore, a potential source of EV-associated protein signatures of TB infection.
Consistent with their cytoplasmic membrane origin, MEVs contain multiple membrane-associated proteins and various proteins found in Mtb culture filtrates. Although our study is limited to three independent cultures, some proteins appeared significantly enriched in MEVs compared to whole cells, suggesting a mechanism for vesicle cargo selection. The packing of selected proteins into bacterial EVs has been demonstrated, and molecular markers for cargo inclusion have been proposed in outer membrane vesicles (OMVs) produced by Gram-negative bacteria [9]. However, in Gram-positive bacteria, the mechanisms of vesicle cargo selection remain largely unexplored. We found no correlation between the low iron-induced gene expression and the preferential inclusion of proteins into MEVs. There were no common physical properties among the enriched proteins except that many are lipoproteins. Additional studies are needed to understand the basis for selective protein packing into EVs in mycobacteria.
Our proteomic analysis indicates that Mtb utilizes MEVs to export proteins that promote its survival and virulence by preserving a favorable host environment and evading host cell death-related bactericidal effects [56,107]. Uncovering export via MEVs enhances our understanding of how particular pathogenicity effectors may function. For instance, until now, NuoG and GroEL, two recognized anti-apoptotic factors, were thought to affect macrophage biology indirectly, as they were believed to be restricted to the bacterial surface. The export within MEVs suggests they could directly target key phagosome-lysosome fusion and apoptosis regulators. Moreover, our findings indicate that bacterial proteins, previously thought to operate endogenously, might interact with the host upon export in MEVs. A notable example is the ATP-binding protein encoded by Rv2623. This protein regulates Mtb growth in mice. A mutant of Rv2623 fails to establish a chronic infection and is hypervirulent in mice and guinea pigs [92]. However, how this protein controls Mtb growth in vivo remains unclear. Our data suggest that the export within MEVs may enable Rv2623 to interface with host pathways that restrict bacterial growth.
Although seemingly paradoxical, our data indicate that MEVs carry proteins that can stimulate the immune system to fight the infection alongside proteins involved in immune evasion and bacterial survival. These include several validated TLR2 agonists and other potent immunostimulators. Activation of TLRs typically promotes immunity. However, prolonged TLR2 activation by agonists in MEVs, such as LpqH, LprG, and LprA, has been shown to induce immunosuppressive cytokines (e.g., IL-10) and inhibit MHC-II antigen presentation [108]. Therefore, prolonged TLR2 activation by MEVs may suppress immune responses, thereby facilitating immune evasion.
Additionally, MEVs may serve as decoys, triggering immune responses that do not aid in pathogen control. Research has shown that exporting antigens from infected macrophages within EVs diverts bacterial proteins from the canonical antigen presentation pathway and limits T cell control of Mtb infection [109,110]. Moreover, exporting immunologically active proteins in MEVs may benefit the pathogen by fostering an environment conducive to transmission. Mtb transmission necessitates cavity development, which requires a tissue-damaging immune response directed against viable bacilli or Mtb antigens. Hence, it is conceivable that Mtb utilizes MEVs to alter the balance between undermining and eliciting host inflammation to facilitate transmission.
The data highlights the diversity and complexity of the MEV proteome, including proteins that can aid the pathogen in surviving in the host and proteins that might place the pathogen at risk of elimination by the immune system. The advantages and disadvantages of bacterial vesicle production are evident from the fast-growing literature describing EVs’ activities and composition, recently reviewed by McMillan and Kuehn [17]. For instance, vesicle production enables membrane remodeling in response to environmental conditions; vesicles are often involved in biofilm formation and maintenance, facilitate nutrient acquisition, promote communication and quorum sensing, and can help disseminate genetic material and protect virulence factors. However, vesicle release requires considerable energy, can activate innate and adaptive immune responses, interfere with colonization by competing with bacteria for attachment sites, and in polymicrobial environments, benefit non-producing cheaters. Nonetheless, the evolutionary conservation of EV production among bacterial pathogens would suggest that the selective export of bacterial products in EVs should ultimately benefit the pathogen.
Our results provide a foundation for further characterization of MEV functionality and bear significant practical implications. For instance, the data identified several well-known adhesins in MEVs that may facilitate interactions between MEVs and host cells. If this is the case, it might be possible to inhibit MEVs by blocking MEV-associated adhesins. Additionally, when used as immunogens, mycobacterial EVs conferred similar protection against Mtb infection as BCG [111], and specific antibodies targeting MEV components identified here, such as LpqH and PstS1, have been shown to protect mice against Mtb infection [112]. Consequently, the immunological characterization of MEV components may assist in engineering EVs enriched with antigens that elicit a protective immune response, thereby contributing to effective vaccine development. Lastly, the overlap between the composition of MEVs and EVs present in the serum of TB patients suggests that incorporating multiple MEV-associated proteins into vesicle-based biomarker platforms could advance research in sputum culture-independent TB diagnosis. One abundant protein in MEVs, LprG, has already demonstrated great promise as a biomarker of TB infection in the pediatric population, a group of patients historically challenging to diagnose with sputum-based assays [113].
In conclusion, although our study is limited to vesicles produced by iron-limited bacteria in culture, it highlighted the diversity and complexity of the MEV proteome, the selective nature of protein incorporation into vesicles, and the potential of MEVs in a plethora of Mtb-host cell interactions. The data indicate that the production of MEVs could benefit the pathogen by contributing to stress relief, nutrient acquisition, virulence, and immune evasion. However, MEVs can also trigger host protective immune activation. This suggests that balancing the beneficial and detrimental activities of MEVs may influence the outcome of TB infection. Lastly, the observed overlap between proteins in MEVs and those detected in EVs in the serum of TB patients suggests that the proteome of MEVs can be a good resource for investigating novel vesicle-associated TB biomarkers and developing culture-independent diagnostic tools.
Supporting information
S1 DataData Set: Proteins with five or more identified peptides in the three replicates.The number of peptides is indicated.(XLSX)
S2 DataData Set: Most abundant proteins selected based on an iBAQ value of 0.1% or higher.(XLSX)
S3 DataData Set. Proteins in MEVs that were found in EVs released during TB infection.(XLSX)
S1 FigCharacterization of MEVs.Purified MEVs were analyzed by electron microscopy, and size distribution was validated by nanoparticle tracking analysis (NTA) conducted using ZetaView (Particle Metrix).(TIF)
S2 FigProteomic data analysis and annotation Process.(PDF)
S3 FigCellular localization of the most abundant proteins in MEVs.(TIFF)
S4 FigFunctional categorization of MEV proteins according to Mycobrowser.(TIFF)
S1 TableProteins enriched in MEVs compared to whole cells.(DOCX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ernst JD. The immunological life cycle of tuberculosis. Nat Rev Immunol. 2012;12(8):581–91. doi: 10.1038/nri 3259 22790178 · doi ↗ · pubmed ↗
- 2Chandra P, Grigsby S, Philips J. Immune evasion and provocation by mycobacterium tuberculosis. Nat Rev Microbiol. 2022;20(12):750–66.35879556 10.1038/s 41579-022-00763-4PMC 9310001 · doi ↗ · pubmed ↗
- 3Gupta S, Rodriguez GM. Mycobacterial extracellular vesicles and host pathogen interactions. Pathog Dis. 2018;76(4):fty 031. doi: 10.1093/femspd/fty 031 29722822 PMC 5930244 · doi ↗ · pubmed ↗
- 4Palacios A, Gupta S, Rodriguez GM, Prados-Rosales R. Extracellular vesicles in the context of Mycobacterium tuberculosis infection. Mol Immunol. 2021;133:175–81. doi: 10.1016/j.molimm.2021.02.010 33743266 PMC 9909588 · doi ↗ · pubmed ↗
- 5Kitagawa R, Takaya A, Ohya M, Mizunoe Y, Takade A, Yoshida S, et al. Biogenesis of Salmonella enterica serovar typhimurium membrane vesicles provoked by induction of Pag C. J Bacteriol. 2010;192(21):5645–56. doi: 10.1128/JB.00590-10 20802043 PMC 2953678 · doi ↗ · pubmed ↗
- 6Kulp AJ, Sun B, Ai T, Manning AJ, Orench-Rivera N, Schmid AK, et al. Genome-Wide Assessment of Outer Membrane Vesicle Production in Escherichia coli. P Lo S One. 2015;10(9):e 0139200. doi: 10.1371/journal.pone.0139200 26406465 PMC 4583269 · doi ↗ · pubmed ↗
- 7Nevermann J, Silva A, Otero C, Oyarzun D, Barrera B, Gil F. Identification of genes involved in biogenesis of outer membrane vesicles (OM Vs) in Salmonella enterica serovar Typhi. Front Microbiol. 2019;10:104.30778340 10.3389/fmicb.2019.00104 PMC 6369716 · doi ↗ · pubmed ↗
- 8Rath P, Huang C, Wang T, Wang T, Li H, Prados-Rosales R, et al. Genetic regulation of vesiculogenesis and immunomodulation in Mycobacterium tuberculosis. Proc Natl Acad Sci U S A. 2013;110(49):E 4790-7. doi: 10.1073/pnas.1320118110 24248369 PMC 3856836 · doi ↗ · pubmed ↗