Two new species of Gymnopus sect. Levipedes (Omphalotaceae, Agaricales) from Central and North China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
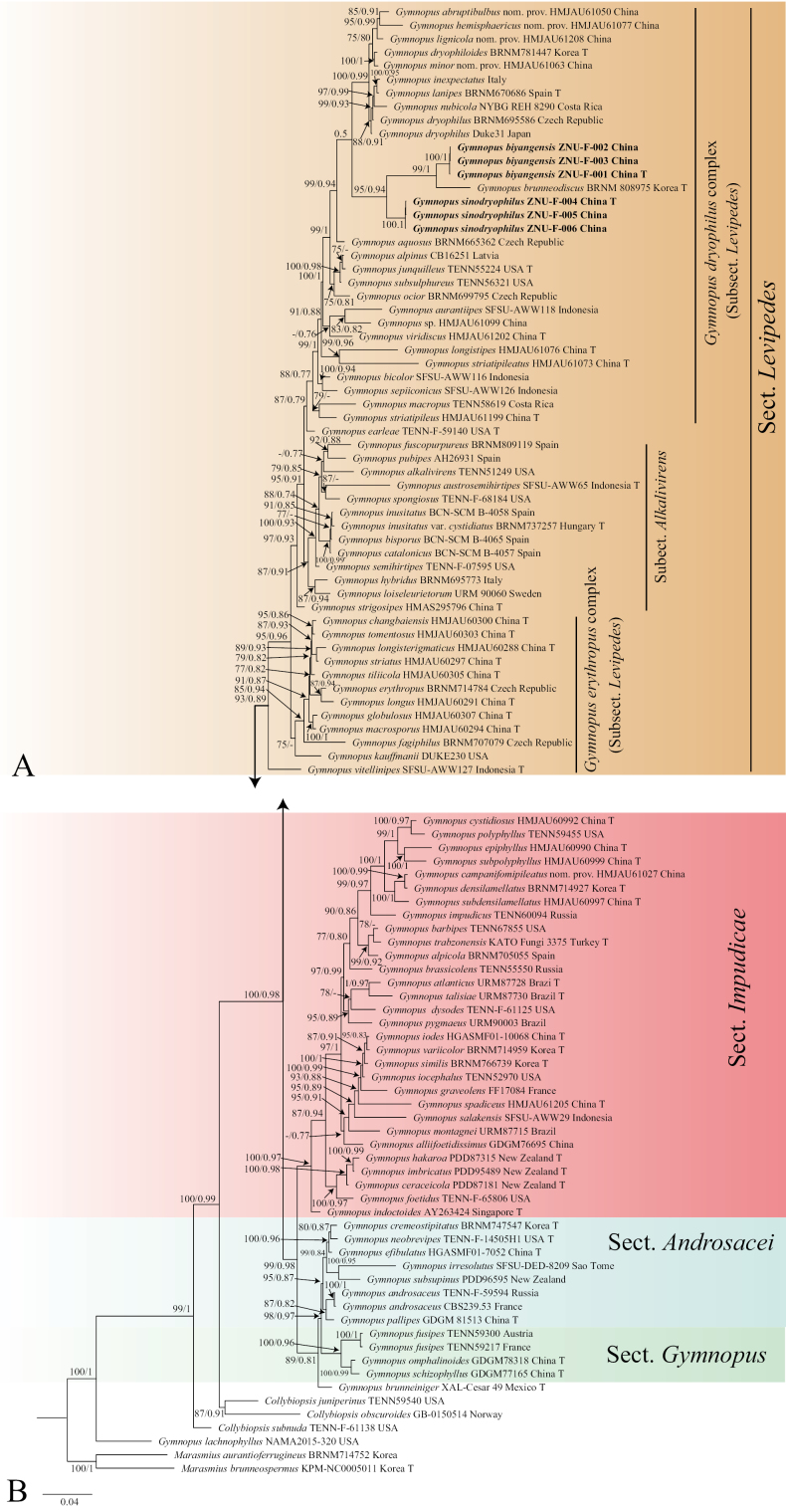 Figure 1
Figure 1 Figure 2
Figure 2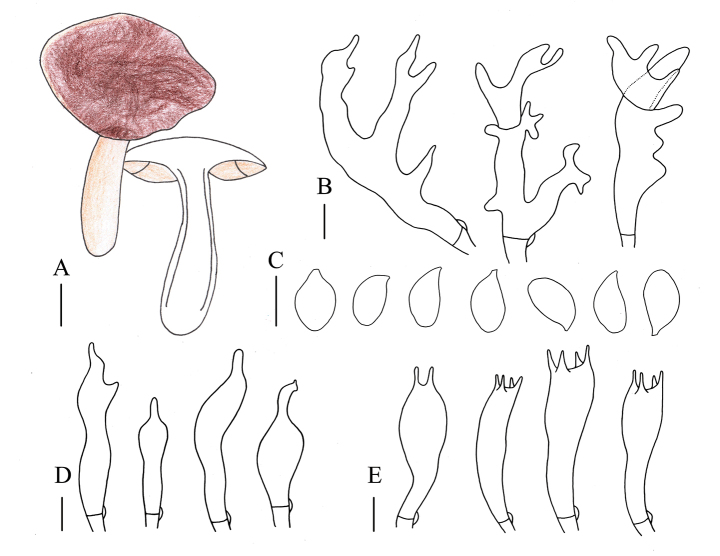 Figure 3
Figure 3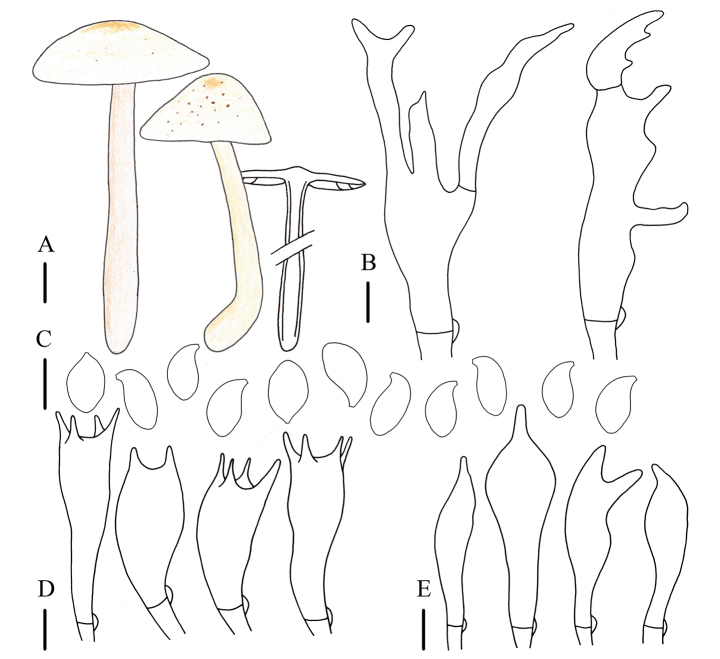 Figure 4
Figure 4| Scientific name | Country | Specimen/Voucher numbers | GenBank Accession Numbers | ||
|---|---|---|---|---|---|
|
|
|
| |||
|
| USA | TENN59540 |
|
| – |
|
| Norway | GB-0150514 |
|
| – |
|
| USA | TENN-F-61138 |
|
| – |
| China | HMJAU61050 |
| – | – | |
|
| USA | TENN51249 |
| – | – |
|
| China | GDGM76695 |
|
| – |
|
| Spain | BRNM705055 |
|
| – |
|
| Latvia | CB16251 |
| – |
|
|
| Russia | TENN-F-59594 |
|
| – |
|
| France | CBS239.53 |
|
| – |
|
| Czech Republic | BRNM665362 |
| – |
|
| Brazi |
|
| – | ||
|
| – | AWW118 |
|
| – |
| Indonesia | SFSU-AWW65 |
| – | – | |
|
| USA | TENN67855 |
|
| – |
|
| – | AWW116 |
|
| – |
|
| Spain | BCN-SCM B-4065 |
|
| – |
|
|
|
|
|
| |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| Russia | TENN55550 |
| – | – |
| Mexico | XAL-Cesar 49 |
| NG-075396 | – | |
|
| Korea | BRNM 808975 |
|
| – |
| China | HMJAU61027 |
|
| – | |
|
| Spain | BCN-SCM B-4057 |
|
| – |
| New Zealand |
| – | – | ||
| China | HMJAU60300 |
|
| – | |
| Korea | BRNM747547 |
|
| – | |
| China | HMJAU60992 |
|
| – | |
| Korea | BRNM714927 |
|
| – | |
|
| Czech Republic | BRNM695586 |
| – |
|
|
| Japan | Duke31 |
| – | – |
| South Korea | BRNM781447 |
|
| – | |
|
| USA | TENN-F-61125 |
|
| – |
|
| USA | TENN-F-59140 |
|
| – |
| China | HGASMF01-7052 |
|
| – | |
| China | HMJAU60990 |
|
| – | |
|
| Czech Republic | BRNM714784 |
| – |
|
|
| Czech Republic | BRNM707079 |
| – |
|
|
| USA | TENN-F-65806 |
|
| – |
|
| Spain | BRNM-809119 |
|
| – |
|
| Austria | TENN59300 |
| – | – |
|
| France | TENN59217 |
|
| – |
| China | HMJAU60307 |
|
| – | |
|
| France | FF17084 |
|
| – |
| New Zealand |
| – | – | ||
| China | HMJAU61077 |
|
| – | |
|
| Italy | BRNM695773 |
| – |
|
| New Zealand |
| – | – | ||
|
| Russia | TENN60094 |
| – | – |
|
| Singapore |
| – | – | |
|
| Italy | – |
|
| – |
|
| Spain | BCN-SCM B-4058 |
|
| – |
| Hungary | BRNM737257 |
|
|
| |
|
| USA | TENN52970 |
|
| – |
| China | HGASMF01-10068 |
|
| – | |
|
| Sao Tome | SFSU-DED-8209 |
| – | – |
| USA | TENN55224 |
| – | – | |
|
| USA | DUKE230 |
| – | – |
|
| USA | NAMA2015-320 |
| – | – |
| Spain | BRNM670686 |
| – |
| |
|
| Sweden | URM 90060 |
|
| – |
| China | HMJAU60288 |
|
| – | |
| China | HMJAU61076 |
|
|
| |
| China | HMJAU60291 |
|
| – | |
|
| Costa Rica | TENN58619 |
| – | – |
| China | HMJAU60294 |
|
| – | |
|
| Brazil |
| – | – | |
| USA | TENN-F-14505H1 |
|
| – | |
|
| Costa Rica | NYBG REH 8290 |
| – | – |
|
| Czech Republic | BRNM699795 |
| – |
|
| China | GDGM 78318 |
|
| – | |
| China | GDGM81513 |
| – | – | |
|
| USA | TENN59455 |
| – | – |
|
| Spain | AH26931 |
|
| – |
|
| Brazil |
|
| – | |
|
| Indonesia | SFSU-AWW29 |
| – | – |
| China | GDGM 77165 |
|
| – | |
|
| USA | TENN-F-07595 |
| – | – |
|
| Indonesia | SFSU-AWW126 |
| – | – |
| Korea | BRNM766739 |
|
| – | |
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
| China | HMJAU61205 |
|
|
| |
|
| USA | TENN-F-68184 |
|
| – |
| China | HMJAU61073 |
|
| – | |
| China | HMJAU60297 |
|
| – | |
| China | HMAS295796 |
|
| – | |
| China | HMJAU60997 |
|
| – | |
| China | HMJAU60999 |
|
| – | |
|
| USA | TENN56321 |
| – | – |
|
| New Zealand |
|
| – | |
| Brazil |
|
| – | ||
| China | HMJAU60305 |
|
| – | |
| China | HMJAU60303 |
|
| – | |
| Turkey | KATO |
| – | – | |
| Korea | BRNM714959 |
|
| – | |
| China | HMJAU61202 |
|
|
| |
| Indonesia | SFSU-AWW127 |
|
| – | |
|
| South Korea | BRNM714752 |
|
| – |
| South Korea | KPM-NC0005011 |
|
| – | |
| 1 | Basidiomata with red stipe |
|
| – | Basidiomata with yellow stipe |
|
| 2 | Stipe covered with dense hairs at the base |
|
| – | Stipe smooth, or covered with sparse hairs at the base |
|
| 3 | Basidia sterigmata extremely long |
|
| – | Basidia sterigmata short |
|
| 4 | Stipe smooth in upper part |
|
| – | Stipe covered with brown pruina on the upper part |
|
| 5 | Pileus pale color, stipe color uneven |
|
| – | Pileus dark color, stipe color uniform |
|
| 6 | Growing on the deciduous layer or rotten branches |
|
| – | Grows at the base of |
|
| 7 | Pileus pale color, near white |
|
| – | Pileus deep color |
|
| 8 | Stipe covered with longitudinal striate |
|
| – | Stipe without longitudinal striate |
|
| 9 | Pileipellis a cuits, typically “ |
|
| – | Pileipellis layered, hyphae inflated to spherical to prolate |
|
| 10 | Apex of cheilocystidia diverticulate |
|
| – | Apex of cheilocystidia not diverticulate |
|
| 11 | Basidiomata marasmioid |
|
| – | Basidiomata collybioid or tricholomatoid |
|
| 12 | Cheilocystidia absent |
|
| – | Cheilocystidia present |
|
| 13 | Caulocystidia present |
|
| – | Caulocystidia absent |
|
| 14 | Pileus green |
|
| – | Pileus not green |
|
| 15 | Stipe cylindrical or clavate |
|
| – | Stipe enlarged at base |
|
| 16 | Stipe light red |
|
| – | Stipe light yellow to yellow |
|
| 17 | Stipe covered with tomentose |
|
| – | Stipe smooth |
|
| 18 | Stipe pale yellow, basidiospores smaller than 6 µm |
|
| – | Stipe light yellow to reddish brown, basidiospores larger than 6 µm |
|
| 19 | Basidiomata and lamellae light yellow |
|
| – | Basidiomata reddish brown, lamellae light reddish brown |
|
- —National Natural Science Foundation of China 501100001809 https://ror.org/01h0zpd94 http://doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Plant and Fungal Species Descriptions · Plant Diversity and Evolution
Introduction
Gymnopus (Pers.) Gray has a long research history dating back to 1801 (Persoon 1801) and plays a significant role in various ecosystems (Mata et al. 2006). In many countries, including China and elsewhere, certain species of Gymnopus are also treated as food resources (Hu and Zhang 2023). As with many macrofungi, Gymnopus (previously classified under the genus Collybia) was first studied in Europe, with significant contributions made by European mycologists (Persoon 1801; Fries 1821; Antonín et al. 1997; Antonín and Noordeloos 1997). Over time, this research enthusiasm spread to other regions (Cooper and Leonard 2013; Antonín et al. 2014; Coimbra et al. 2015; Deng 2016; Oliveira et al. 2019; Hu et al. 2022a, b), particularly North America, where many studies have been conducted (Halling 1996; Desjardin et al. 1999; Mata et al. 2006; Petersen and Hughes 2014).
However, compared to other genera, such as Agaricus L. and Cantharellus Adans. ex Fr., Gymnopus has not been as thoroughly studied. This discrepancy in research interest is partly due to regional differences in mycologists’ areas of focus, which, have led to significant research gaps between Gymnopus and more widely studied genera. For example, approximately 300 species of Gymnopus have been described globally (Hu et al. 2022a, b), with the majority of species documented in North America, Europe, and Asia (Sun et al. 2021b; Hu and Zhang 2023). In contrast, fewer species have been reported in Oceania and Africa (Hu and Zhang 2023).
In recent decades, Asia has emerged as a new hotspot for Gymnopus research, evidenced by the discovery of new species or new combinations (Hu and Zhang 2023). This indicates an increasing recognition of the genus’ biodiversity and ecological importance. Nevertheless, the full extent of Gymnopus species diversity remains underappreciated and warrants further exploration.
SubsectionLevipedes is characterized by a stipe that is smooth, polished, or pubescent; a pileipellis that typically forms an entangled (never radially oriented) trichoderm, consisting of inflated, often lobed elements or coralloid “dryophila-type” structures; trama and its elements are non-dextrinoid, and usually saprophytic, commonly found in coarse humus, forest litter, or on decaying wood (Halling 1996; Antonín and Noordeloos 2010; Oliveira et al. 2019). In this subsection, some species belonging to the Gymnopusdryophilus (Bull.) Murrill complex have historically been treated as food resources in Northeast and Southwest China (Hu and Zhang 2023). Despite this, recent studies have shown that the species complex, Gymnopusdryophiloides, can cause diarrhea in humans (Ma et al. 2024). Furthermore, due to the similarity in appearance, difficulties remain in identifying the species of this complex. Although Vilgalys and Miller (1983, 1987a, b) and Antonín et al. (2013) have carefully studied the North American or European G.dryophilus species complex and identified key characteristic features, all these results were based on the North American or European specimens. Additionally, there is still a lack of comprehensive study on the complex outside of North America and Europe.
Looking back at the research history, it is evident that taxonomic studies are mainly focused on Europe and North America, leaving a significant gap on other continents. As research on Gymnopus deepens, taxonomic issues and questions about the classification of the genus emerge, highlighting the need for further attention to this group. Consequently, taxonomic research on Gymnopus has begun in China. Specimens of the G.dryophilus complex collected from Henan and Jilin Provinces are studied in detail here. As a result, two new species belonging to Gymnopussect.Levipedes are described and illustrated. This study enriches our understanding of the species diversity of Gymnopus and provides valuable insights for future studies.
Materials and methods
Specimen collection
The specimens used in this study were collected between 2020 and 2023 in Henan and Jilin Provinces, China. All specimens were photographed in situ, with an emphasis on capturing basidiomata at various stages of development. Subsequently, three or more basidiomata were collected for detailed morphological and molecular analysis. The morphological characteristics, including size, color, and odor, were documented. The color references followed Flora of British Fungi: Color Identification Chart (Royal Botanic Garden 1969). A clean tissue sample from each specimen was dried using allochroic silica gel for DNA extraction. The specimens were dried in an electric oven at approximately 45 °C.
Identification
The recognition and description of macro-characteristics were based on field notes and photographs. Dried specimens were rehydrated in 94% ethanol and subsequently mounted in 3% potassium hydroxide (KOH), 1% Congo Red, or Melzer’s Reagent for examination. Structures such as basidiospores, basidia, and cheilocystidia were observed using a Zeiss Axio Lab. A1 microscope. For each specimen, a minimum of 40 measurements were taken from at least two different basidiomata. The size of basidiospores is expressed as length × width (L × W). To account for size variation, 5% of the measurements from each end of the range were excluded, and the final measurements are given as (a) b × c (d). Q represents the ratio of L to W for each studied specimen, while Qm denotes the average Q value ± standard deviation. The examined specimens are deposited in the Herbarium of Mycology at Zhejiang Normal University (ZNU-F).
DNA extraction, PCR, and sequencing
Total DNA was extracted from dried materials using the NuClean Plant Genomic DNA Kit (Kangwei Century Biotechnology Company Limited, Beijing, China), following the manufacturer’s instructions. The internal transcribed spacer (ITS) region, nuclear large ribosomal subunits (nLSU), and translation elongation factor (tef-1α) loci were selected for phylogenetic analysis. The primer pairs ITS4-ITS5 (Gardes and Bruns 1993), LROR-LR5/LR7 (Vilgalys and Hester 1990; Cubeta et al. 1991), and 983F-1567R (Rehner and Buckley 2005) were used to amplify the ITS, nLSU, and tef-1α, respectively.
PCR reactions (25 μL) were prepared as follows: 8.5 μL of dd H_2_O, 12.5 μL of 2 × Taq PCR MasterMix, 1 μL of each primer, and 2 μL of DNA sample. The reaction conditions were based on those described by Coimbra et al. (2015) for ITS, Ryoo et al. (2020) for nLSU, and Xu et al. (2021) for tef1-α. PCR products were visualized under UV light following electrophoresis on 1% agarose gels stained with ethidium bromide. The PCR products were then sent to Hangzhou Huada-Qinglan Innovation Technology Co., Ltd. for sequencing, using the Sanger method. The new sequences were deposited in GenBank (http://www.ncbi.nlm.nih.gov/genbank), and the detailed sequence information is provided in Table 1.
Phylogenetic analysis
Based on the BLASTn results and morphological similarities, sequences related to these samples were collected (Table 1). A combined dataset of ITS, nLSU, and tef-1α fragments, consisting of 85 sequences obtained from the type species of Gymnopus and Marasmius Fr., was used for phylogenetic analysis. Species belonging to Marasmius, Marasmiusaurantioferrugineus Hongo and Marasmiusbrunneospermus Har. Takah., were selected as outgroups (Hu et al. 2024).
Each gene region in the dataset was aligned using MAFFT 7.490 (Katoh and Standley 2013) and subsequently manually inspected in BioEdit 7.0.5.3 (Hall 1999). The alignments of the ITS, nLSU, and tef-1α sequences were then combined through PhyloSuite 1.2.2 (Zhang et al. 2020). A partition homogeneity test (PHT) (Farris et al. 1994) was performed on the multi-gene dataset with PAUP 4.0b10 (Swofford 2002), employing 1000 homogeneity replicates. The best-fit evolutionary model was estimated using ModelFinder (Kalyaanamoorthy et al. 2017). Bayesian inference (BI) was applied for phylogenetic analysis, utilizing MrBayes 3.2.6 with a general time-reversible DNA substitution model and gamma distribution rate variation across the sites (Ronquist and Huelsenbeck 2003). Four Markov chains were run for two independent runs, starting from random trees, until the split frequency value fell below 0.01. Trees were sampled every 100 generations, with the first 25% discarded as burn-in. The remaining trees were used to construct a 50% majority consensus tree and calculate the Bayesian posterior probabilities (PP). Maximum likelihood (ML) analysis was performed using RaxmlGUI 2.0.10 (Edler et al. 2021) with 1000 bootstrap (BS) replicates to search for the optimal topology. The resulting trees were visualized using FigTree 1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/, accessed 25 October 2020).
Results
Phylogenetic analysis
A total of 18 new sequences (six per locus) were obtained from six samples in this study. In the combined dataset, 189 sequences derived from three gene loci (ITS, nLSU, and tef-1α) from 102 samples were used for phylogenetic analysis. The best-fitting model for BI was GTR+F+I+G4, while the GTRGAMMA model was applied for ML (Vizzini et al. 2015). The Bayesian analysis was run for four million generations, resulting in an average standard deviation of split frequencies of 0.004528. The same dataset and alignment were also analyzed using the ML method. Both phylogenetic analyses yielded a similar topology, which is depicted in Fig. 1.
Maximum likelihood analysis generated from the combined ITS, nLSU, and tef-1α dataset of genus Gymnopus. Bootstrap values (BS) ≥ 75% from ML analysis and Bayesian posterior probabilities (PP) ≥ 0.80 are shown on the branches. Newly sequenced collections are indicated in bold, and the type specimens are denoted by (T).
Our phylogenetic analysis revealed that species of the G.dryophilus complex form a distinct clade, which is sister to the Gymnopuserythropus (Pers.) Antonín, Halling & Noordel. complex. Two newly proposed species are independently positioned within the genus Gymnopus, with strong phylogenetic evidence.
Taxonomy
Gymnopus
biyangensis
Taxon classificationFungiAgaricalesOmphalotaceae
J.J. Hu, B. Zhang, X. Li & Y. Li sp. nov.
87617FF1-925E-5083-8D1A-F43F68E49494
Fungal Names: FN 572234
Etymology.
Refers to the location of type material.
Diagnosis.
[English] This species is characterized by the basidiomata that appear in summer and originate from broad-leaved forests, dark reddish-brown pileus, cylindrical to clavate stipe, clavate to cylindrical cheilocystidia with a narrowly protruding apex.
The habit of Gymnopus spp. described in this study: AGymnopusbiyangensisBGymnopuschangchunensis. Scale bars: 1 cm.
Type.
China. Henan Province • Zhumadian City, Biyang County, Mingzhuang Village, 10 July 2021, Jia-Jun Hu, Bo Zhang, and Xiao Li, ZNU-F-001 (Collection No.: Hu 769), holotype.
Morphological characteristics of Gymnopusbiyangensis (ZNU-F-001) A basidiomata B pileipellis elements C basidiospores D cheilocystidia E basidia. Scale bars: 1 cm (A); 5 µm (B–E).
Description.
Basidiomata medium-sized. Pileus 3.3–4.4 cm in diameter, applanate-convex, reddish brown to brown, smooth, glabrous; margin entire, wavy to upturned, dark reddish-brown to light brown. Context thin, yellow to light brown, freshy, odorless. Stipe 2.0–5.0 cm long and 0.5–1.3 cm wide, central, cylindrical to clavate, smooth, light yellow, occasionally with reddish brown tones or slight spots at the base. Lamellae adnate to adnexed, close, yellowish brown to light brown, unequal. Occurrence in leaf litter.
Basidiospores 5.0–6.0 × 3.0–4.0 µm, Q = (1.25)1.30–1.93, Qm = 1.64 ± 0.19, elliptic, hyaline, smooth, inamyloid, thin-walled. Basidia (12)16–23 × 3–6 µm, clavate to cylindrical, 2- or 4-spored, hyaline, smooth, thin-walled. Cheilocystidia (16)17–28(29) × 3–6 µm, cylindrical to clavate with mamiform, often longer apical projections, less commonly weakly coralloid, hyaline, smooth, thin-walled. Pleurocystidia and caulocystidia not observed. Pileipellis a “dryophila-type” cutis, 8–14 µm wide, hyaline, smooth, thin-walled. Clamp connections present in all tissues.
Habit, habitat, and distribution.
Scattered to gregarious. Saprotrophic, with humicolous habitat, found in broad-leaved forests. So far, it is only known from Henan Province, China.
Other specimens examined.
China. Henan Province • Zhumadian City, Biyang County, Mingzhuang Village, 10 July 2021, Jia-Jun Hu, Bo Zhang, and Xiao Li, ZNU-F-002 (Collection No.: Hu 770) • Zhumadian City, Biyang County, Mingzhuang Village, 10 July 2022, Jia-Jun Hu, Bo Zhang, and Xiao Li, ZNU-F-003 (Collection No.: Hu 775).
Note.
This species is characterized by the basidiomata occurring in the summer, a dark reddish-brown pileus, a cylindrical to clavate stipe, an apex of cheilocystidia that is not diverticulate or lobate.
Gymnopusbiyangensis is similar to species in the G.erythropus complex due to the dark reddish-brown pileus. However, this species differs from G.erythropus by the light yellow and cylindrical to clavate stipe, smaller basidiospores, and a non-encrusted pileipellis. Gymnopusbiyangensis can be distinguished from G.fagiphilus by the smooth and light-yellow stipe and smaller basidiospores.
Gymnopus
sinodryophilus
Taxon classificationFungiAgaricalesOmphalotaceae
J.J. Hu, B. Zhang & Y. Li sp. nov.
C279EFAE-8B3E-550D-B62C-3D08B389C61E
Fungal Names: FN 572235
Etymology.
Refers to the species similar to G.dryophilus.
Diagnosis.
[English] This species is characterized by the yellowish-white to light-brown basidiomata, arisen from coniferous and broad-leaved mixed forest, coralloid but inflated pileipellis elements, and a non-coralloid apex of cheilocystidia.
Morphological characteristics of Gymnopussinodryophilus (ZNU-F-003) A basidiomata B pileipellis elements C basidiospores D basidia E cheilocystidia. Scale bars: 1 cm (A); 5 µm (B–E).
Type.
China. Jilin Province • Changchun City, Jingyue District, Mt. Lianhua, 09 August 2021, Jia-Jun Hu and Bo Zhang, ZNU-F-005 (Collection No.: Hu 809), holotype.
Description.
Basidiomata small to medium. Pileus 1.0–4.4 cm in diameter, applanate-hemispheric to convex, light yellow to light brown, darker at center, brown, occasionally with brown spots, with an umbo sometimes, smooth, glabrous; margin entire, involute, yellowish white to light yellow. Context thin, fresh, white to light yellow, odorless. Stipe 1.3–8.8 cm long and 0.3–0.9 cm wide, central, clavate, light brown to brown, paler downwards, becoming light yellow to yellowish white, striped, smooth, glabrous, fistulose, fibrous. Lamellae adnexed, close to crowded, yellow to light brown, unequal. Occurrence in leaf litter in mixed forest.
Basidiospores (4.0)5.0–6.0(6.2) × 3.0–3.8 µm, Q = (1.33)1.39–2.00(2.07), Qm = 1.67 ± 0.18, elliptic, hyaline, smooth, inamyloid, thin-walled. Basidia (12)13–23 × 4–8 µm, clavate to cylindrical, 2- or 4-spored, hyaline, smooth, thin-walled. Cheilocystidia (11)13–27(31) × 3–8 µm, cylindrical to clavate, umbonate-mamiform with a short apical projection, occasionally forked, hyaline, smooth, thin-walled. Pleurocystidia and caulocystidia not observed. Pileipellis a “dryophila-type” cutis, (5)7–10(12) µm wide, hyaline, smooth, thin-walled. Clamp connections present in all tissues.
Habit, habitat, and distribution.
Scattered to gregarious. Saprotrophic, with humicolous habitat, found in mixed forests. So far, only known from Jilin Province, China.
Other specimens examined.
China. Jilin Province • Yanbian Korean Autonomous Prefecture, Antu County, Erdaobaihe Town, 26 June 2021, Jia-Jun Hu and Bo Zhang, ZNU-F-006 (Collection No.: Hu 743) • Changchun City, Jingyue District, Jingyuetan National Forest Park, 08 August 2021, Jia-Jun Hu, Bo Zhang, ZNU-F-004 (Collection No.: Hu 807) • Changchun City, Jingyue District, Jingyuetan National Forest Park, 08 August 2021, Jia-Jun Hu, Bo Zhang, ZNU-F-007 (Collection No.: Hu 811) • Changchun City, Jingyue District, Jingyuetan National Forest Park, 08 August 2021, Zheng-Hao Zhang, Jia-Jun Hu, Bo Zhang, ZNU-F-008 (Collection No.: Hu 892).
Note.
This species is characterized by the yellowish-white to light-brown basidiomata, which arise in summer; coralloid but inflated pileipellis elements; and a non-diverticulate apex of cheilocystidia.
This species is extremely similar to G.dryophilus due to the analogous morphology. However, this species differs from G.dryophilus by appearance in summer, smaller basidiospores, and the apex of cheilocystidia not being diverticulate.
Key to the reported species of Gymnopussubsect.Levipedes in China
**: **
Discussion
In this study, two new species within the G.dryophilus complex are proposed. Gymnopusbiyangensis is characterized by summer-fruiting basidiomata found in broad-leaved forests, a dark reddish-brown pileus, a cylindrical to clavate stipe, and clavate to cylindrical cheilocystidia with a long apical projection. In contrast, Gymnopussinodryophilus is characterized by yellowish-white to light-brown basidiomata that arise from coniferous and broad-leaved mixed forests, coralloid but inflated pileipellis elements, and cheilocystidia with a short apical projection.
The sect. Levipedes is divided into two subsections: subsect. Alkalivirentes Antonín & Noordel. and subsect. Levipedes Antonín & Noordel., based on whether the mycelium turns green in potassium hydroxide (KOH) and ammonium hydroxide (NH_4_OH) (Halling 1981; Antonín and Noordeloos 1997).
Species from the G.erythropus, G.fagiphilus (Velen.) Antonín, Halling & Noordel., and G.dryophilus complexes primarily comprise subsect./ Levipedes. Our previous work demonstrated that the G.dryophilus complex species clearly differs from the G.erythropus complex species, particularly in terms of seasonal occurrence and the shape of cheilocystidia (Hu et al. 2022b, Hu 2023). The species of subsect.Levipedes are characterized by smooth, polished, or pubescent stipes; pileipellis is typically an entangled trichoderm (never radially oriented), composed of inflated, often lobed elements or coralloid, “dryophila-type” structures; trama and its elements are non-dextrinoid (Halling 1996; Antonín and Noordeloos 2010; Oliveira et al. 2019). However, in our study, G.sinodryophilus exhibited branched but not broadened pileipellis elements, which contrasts with the established boundaries. Similar deviations have been observed in Gymnopusglobulosus J.J. Hu, Y.L. Tuo, B. Zhang & Yu Li, Gymnopusearleae Murrill, and Gymnopuskauffmanii (Halling) Halling, etc. These findings suggest that the current limits and taxonomic framework of subsect.Levipedes require further examination.
Although the taxonomic history of Gymnopus (formerly Collybia) spans over two centuries, research on this genus remains considerably behind other groups, such as Amanitaceae (Cui et al. 2018), boletes (Wu et al. 2016), and Ganodermataceae (Sun et al. 2022). Although some species are edible or possess potential applications, such as in wastewater treatment (Sun et al. 2021a) and biological control (Wu 2016), they have not gained the same recognition as culinary fungi, for example, Morchella Dill. ex Pers. or Tuber P. Micheli ex F.H. Wigg. This lack of prominence may be a key reason why this genus has been overlooked in research. To truly understand this group, greater attention must be devoted to its study.
Supplementary Material
XML Treatment for Gymnopus biyangensis
XML Treatment for Gymnopus sinodryophilus
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Antonín V Noordeloos ME (1997) A monograph of Marasmius, Collybia and related genera in Europe. Part 2: Collybia, Gymnopus, Rhodocollybia, Crinipellis, Chaetocalathus, and additions to Marasmiellus. Libri Botanici 17: 1–256.
- 2Antonín V Noordeloos ME (2010) A monograph of marasmioid and collybioid fungi in Europe. IHW Verl, Eching, Germany.
- 3Antonín V Halling R Noordeloos M (1997) Generic concepts within the groups of Marasmius and Collybia sensu lato.Mycotaxon 63: 359–368.
- 4Antonín V Sedlák P TomšovskýM (2013) Taxonomy and phylogeny of European Gymnopussubsectionlevipedes (Basidiomycota, Omphalotaceae).Persoonia 31: 179–187. 10.3767/003158513 X 67404324761042 PMC 3904049 · doi ↗ · pubmed ↗
- 5Antonín V Ryoo R Ka KH (2014) Marasmioid and gymnopoid fungi of the Republic of Korea. 7. Gymnopussect.Androsacei. Mycological Progress 13: 703–718. 10.1007/s 11557-013-0953-z · doi ↗
- 6Coimbra VR Pinheiro FG Wartchow F Gibertoni TB (2015) Studies on Gymnopussect.Impudicae (Omphalotaceae, Agaricales) from Northern Brazil: Two new species and notes on G.montagnei. Mycological Progress 14: 1–9. 10.1007/s 11557-015-1131-2 · doi ↗
- 7Cooper J Leonard P (2013) Three new species of foetid Gymnopus in New Zealand.Myco Keys 7: 31–44. 10.3897/mycokeys.7.4710 · doi ↗
- 8Cubeta M Echandi E Abernethy T Vilgalys R (1991) Characterization of anastomosis groups of binucleate Rhizoctonia species using restriction analysis of an amplified ribosomal RNA gene.Phytopathology 81: 1395–1400. 10.1094/Phyto-81-1395 · doi ↗