Coxiella burnetii as a model system for understanding host immune response against obligate intracellular, vacuolar pathogens
Akhila Sasikala, Chandhana Prakash, Mullai Valli Ramamoorthy, Sandhya Ganesan

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
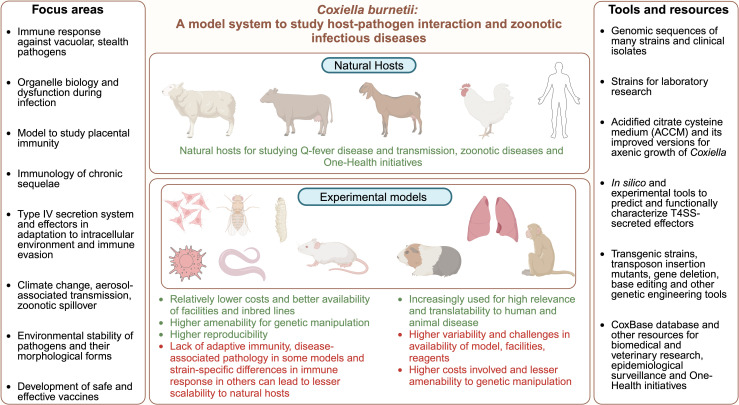 Fig 1
Fig 1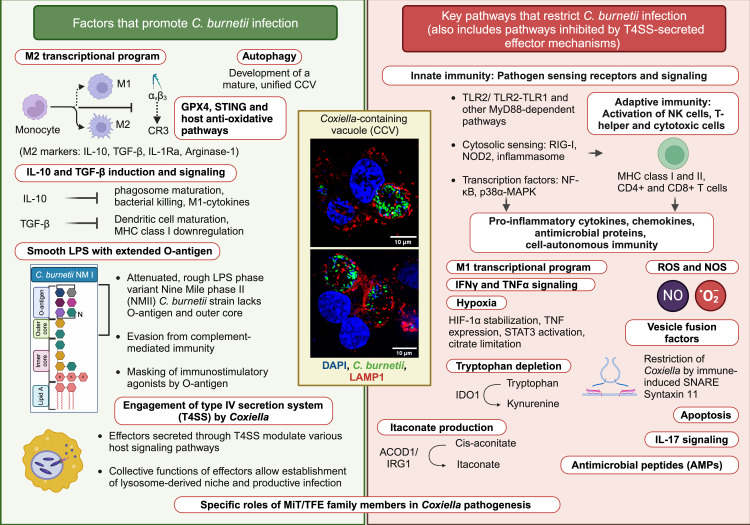 Fig 2
Fig 2- —Science and Engineering Research Board
- —http://dx.doi.org/10.13039/501100009053The Wellcome Trust DBT India Alliance
- —Department of Biotechnology
- —Indian Institute of Science Education and Research
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Viral Infections and Vectors · Leptospirosis research and findings
Introduction
Many intracellular bacterial pathogens have evolved to establish a secure vacuolar niche for replication in mammalian hosts through stealthy lifestyles and sophisticated mechanisms of virulence and adaptation. In particular, the bacterial pathogen Coxiella burnetii is increasingly gaining attention as an emerging zoonotic pathogen and an under-appreciated model to study host immune response against obligate intracellular pathogens. Here, we summarize the features that render Coxiella an excellent model to study basic, applied, and epidemiological questions in infectious disease research (Fig 1). Further, we provide a cheat sheet on how Coxiella has facilitated our understanding of innate, adaptive, and cell-autonomous immune responses to obligate intracellular pathogens (Fig 2).
Coxiella burnetii: A model system to study host–pathogen interaction and zoonotic infectious diseases.Technological advances in the field of C. burnetii research, including the development of axenic media, availability of virulent and attenuated strains for laboratory research, in silico and experimental tools to predict and functionally characterize T4SS-secreted effectors (SecReT4, DeepSecE, T4SEpp) [92–94], enhanced genetic toolkit [21–24,88,89,95,96] and other resources (example: CoxBase) [97], as listed on the right panel, have facilitated better understanding of interaction between host and C. burnetii at a molecular, cellular, and organism level. Bottom panel depicts some of the experimental host systems employed, including, epithelial cells/ fibroblasts, macrophages of different animal origins, Drosophila melanogaster, Caenorhabditis elegans, Galleria mellonella, wildtype and SCID mice, guinea pigs, human ex vivo lung model, and non-human primates. These models are characterized by a spectrum of advantages and limitations, as highlighted by green and red text, respectively. These developments are expected to promote modeling of zoonotic disease biology in natural hosts (top panel) and research efforts in several key focus areas including dysregulation of lysosome biology by pathogens, placental immunity [98], effector functions, molecular mechanisms underlying chronic sequelae, environmental stability of Coxiella, impact of rising climate change, and vaccine development (left panel). Created with Biorender.com.
Factors promoting or restricting Coxiella burnetii infection.Development of bacterial replication-permissive, lysosome-derived compartment and intracellular bacterial proliferation (shown in the central boxed panel) is supported by the Coxiella T4SS and the translocation of numerous effectors into the host cell, M2 transcriptional program including upregulation of immunosuppressive cytokines TGF-β and IL-10, autophagy, and host anti-oxidative mechanisms. Further, the smooth LPS variants produced by the C. burnetii NMI strain mask the bacterial surface lipoproteins and prevent TLR2 activation, thus inhibiting pathogen sensing in the host and allowing the establishment of disease. At the same time, the host restricts Coxiella through activation of innate, adaptive, and cell-autonomous immune mechanisms including multiple pathogen recognition receptor-based pathways, activation of T and NK cells, secretion of pro-inflammatory cytokines, especially IFNγ, TNF, and IL-17, production of ROS and RNS, activation of the M1 program in macrophages, tryptophan depletion, production of itaconate, expression of STX11, antimicrobial peptide production, hypoxic conditions, and apoptosis. MiT/TFE family of transcription factors regulate Coxiella vacuole biogenesis and intracellular bacterial replication and the precise contribution of each, in promoting or restricting different stages of infection, requires further investigation. Central Boxed panel: Immunofluorescent images of TPA-differentiated THP1 cells infected with C. burnetii NMII for 3 days and fixed with 4% PFA. LAMP1, lysosome-associated membrane protein-1; TPA: 12-O-tetradecanoylphorbol-13-acetate; THP1: human monocyte cell line. Figure created with Biorender.com. Credit for immunofluorescent images in the central boxed panel: Ms. Amrita Bhattacharya, IISER Thiruvananthapuram.
Coxiella burnetii: An emerging zoonotic pathogen and a great model system for biomedical research
C. burnetii was named in honor of Herald Rea Cox and Macfarlane Burnet, who initially identified and characterized this infectious agent along with Gordon Davis and Marvis Freeman [1,2]. C. burnetii belongs to the class Gammaproteobacteria, order Legionellales, and family Coxiellaceae. C. burnetii is a CDC select agent and NIAID priority pathogen due to its low infectious dose, high aerosol transmissibility, environmental stability, history of multiple outbreaks, and potential as an agent of bioterrorism [3,4]. The first and largest outbreak occurred in the Netherlands between 2007 and 2010, with >4000 human cases, presenting an unprecedented challenge to veterinary and public health [5]. Reservoirs of Coxiella include, but are not limited to, domestic animals (cattle, sheep, goats), wild birds, and rodents. Coxiella typically causes an acute febrile illness called “query- or Q-fever” and adversely affects animal reproductive fitness and productivity. Coxiella is transmitted from infected animals to humans primarily through aerosols, handling of infected tissues or birthing products, occupational exposure, and consumption of contaminated animal products*.* In humans, acute Q-fever is treatable with doxycycline. However, severe complications may also arise, including endocarditis, pneumonia, or a chronic fatigued condition called Q-fever fatigue syndrome [6]. While Q-fever is reportable in some countries (USA), it is not yet notifiable in many countries. The vaccine Q-vax (formalin-inactivated virulent strain) is licensed only in Australia and requires pre-screening due to the hypersensitivity of patients previously exposed to Coxiella. Coxevac, a veterinary vaccine, is available only in certain parts of Europe. Thus, there is a compelling need for a globally approved, safe vaccine.
The process of transition from a free-living/tick-associated ancestral lifestyle to an obligate intracellular environment is accompanied by a reduction in genome size, and loss/pseudogenization of many genes [7]. For instance, C. burnetii, containing a 2 Mb genome, has retained genes for heme, biotin, and LPS biosynthesis but is auxotrophic for almost 11 amino acids [7,8]. The most widely used strains for laboratory research are the Nine Mile Phase I (virulent and requires BSL-3 containment) and the attenuated phase variant, Nine Mile Phase II that was generated through continuous passage of NMI in yolk sacs and is suitable for use in BSL-2 laboratories [9]. NMI and NMII differ in their O-antigen and outer polysaccharide core but are indistinguishable in their lipid A and exhibit similar intracellular lifecycles [10–14] (Fig 2). In addition to phase variation, C. burnetii has two morphological forms, the spore-like small-cell variant (SCV) and the replicative large-cell variant (LCV) [15]. LCVs are metabolically more active and larger in size, whereas the SCVs exhibit greater stability to environmental stresses owing to a thick and remodeled peptidoglycan (PGN) layer [16,17].
At a single organism level, Coxiella is tropic to alveolar macrophages, phagocytic cells, placental, and heart tissues, but can infect multiple other cell types. Entry into host cells is followed by extensive fusion of the nascent vacuole with endolysosomal, autophagic, and secretory vesicles to generate a mature, acidic, spacious vacuole called Coxiella-containing vacuole (CCV) [18] (Fig 2). CCV biogenesis is mediated by bacterial proteins (called effectors) secreted through the Dot/Icm type IVB secretion system (T4SS) [19–24].
Thus, Coxiella (i) is evolutionarily adapted to and replicates in lysosomal-like environments, (ii) exhibits prolonged and persistent intracellular survival, and (iii) employs an extensive repertoire of effectors to intercept host vesicle traffic and manipulate host signaling. Here, we review the key determinants of protective host response against stealthy mammalian-adapted pathogens as learned from host and Coxiella-centric studies.
Innate immunity
The two main virulence determinants of Coxiella are its LPS and the T4SS (Fig 2). Acute response is typically initiated by the detection of Coxiella by pattern-recognition receptors (PRRs) of the host cells, the production of pro-inflammatory cytokines (TNF, IL-1, and IL-18), the recruitment of monocytes to infection sites, and the formation of granulomas [25,26]. Despite being a Gram-negative pathogen, Coxiella LPS is tetra-acylated and is a weak activator of TLR4 [13,27]. TLR2, NOD2, and TLR1/TLR2 heterodimer also play an important role in cytokine production [13,27–31]. However, the precise mechanisms that allow cytosolic detection of the PGN derivatives of this vacuole-residing pathogen are unclear. The smooth LPS variant of NMI masks bacterial surface lipoproteins and evades detection by TLR2 [32] and complement-mediated killing [33]. It engages with ⍺V_β_3 integrin but inhibits CR3 activation, allowing higher survival inside monocytes compared to strains with rough LPS [34]. Other strains with (i) intermediate LPS length (NM Crazy), (ii) smooth LPS but deficient in Dot/Icm function (NMI ∆dot/icm), (iii) natural NMII revertants with intermediate LPS (NMII-E), and (iv) smooth LPS but attenuated in animal models (Dugway) have also emerged as key resources for dissecting the distinct roles of LPS, its length, and the T4SS in driving innate immune responses [35–38]. T4SS-secreted effectors inhibit the activation of several innate immune signaling pathways including inflammasome, NF-κB, p38α-MAPK, IL-17, and RIG-I, to dampen cytokine response and promote bacterial replication [39–46]. Coxiella effectors also inhibit type I IFN production, which appears to play a dual role in tissue-specific models of infection [46,47].
Further, Coxiella induces the M2 transcriptional program and immunosuppressive cytokines, rendering monocytes and macrophages permissive to bacterial replication [48]. While TGF-β downregulates MHC-I and impairs dendritic cell activation, IL-10 signaling impairs phagosome maturation and bacterial killing, downregulates M1-cytokines including TNF and contributes to the development of chronic Q-fever [49–51].
Adaptive immunity
Replication in a lysosomal-like vacuole allows the infected cell to present antigenic peptides and elicit T-helper cell immunity. Interestingly, both MHC-I and MHC-II are required for protective immunity, with one compensating in the other’s absence [52] (Fig 2). Consistent with this, mice lacking both CD4+ and CD8 + T cells succumb to Coxiella infection but are rescued by reconstitution of either of the T cell types [53,54]. MHC-I and CD8+ T cell-mediated immunity limits the spread of the bacterium, likely in part, through effector cytokines IFNγ and TNF [52]. However, the mechanisms that mediate MHC-I presentation of Coxiella antigens are not well understood. Interestingly, chronic Q fever is distinguished by high circulating levels of IFNγ and a high IFNγ/IL-2 ratio [55]. B cells limit tissue damage and disease severity but are not sufficient to control Coxiella [53,56]. However, antibody response is a useful serological diagnostic, since acute and chronic Q-fever are typically associated with predominantly high titer of antibodies against NM phase II and phase I, respectively [6].
Cell-autonomous defense
Activation of the M1-program in infected cells inhibits intracellular Coxiella replication and maturation of the Coxiella vacuole (Fig 2). IFNγ, TNF, IL-17, MyD88-dependent and miRNA pathways restrict Coxiella, in part, by iNOS, NADPH, ACOD1, and IDO1 that mediate production of Nitric Oxide (NO), Hydrogen peroxide (H_2_O_2_), itaconate, tryptophan catabolism, apoptosis, and other unidentified mechanisms [27,45,57–66]. In hypoxic conditions, decreased availability of citrate/TCA metabolites also suppress intracellular Coxiella replication [67].
Microphthalmia/transcription factor E (MiT/TFE) family of proteins, in particular, TFEB and TFE3 appear to play a complex role in Coxiella infection. On one hand, overexpression of TFEB was observed to restrict Coxiella replication [68], and consequently, Coxiella inhibiting nuclear translocation of TFEB through T4SS-secreted effectors [69]. On the other hand, TFEB as well as TFE3 have been observed to be activated during infection [70,71] and double-deficiency was consistently observed to yield smaller CCVs [69–71]. However, TFEB/TFE3-double deficiency, depending on the genetic and cell type model, (KO mouse macrophages, siRNA/CRISPR-based KO HeLa) yielded either a concomitant decrease, increase or minimal impact on intracellular Coxiella replication [69–71]. It is possible that these seemingly divergent observations are likely because some TFEB/TFE3-regulated processes such as autophagy, lysosomal biogenesis are subverted by Coxiella for CCV expansion and fusogenicity, while others such as excessive CCV acidification and Cathepsin activity negatively influence bacterial replication inside the vacuole [68]. The specific contribution of TFEB, TFE3, MITF, and their temporal regulation during infection requires further investigation.
In addition to direct killing mechanisms, our work demonstrated that the expression of an immune-inducible SNARE, Syntaxin 11, which mediates vesicle fusion, also decreases bacterial replication and CCV expansion [72].
Conversely, host antioxidative pathways that reduce ROS levels, such as Glutathione Peroxidase 4 (GPX4) in humans, and a cytosolic sensor for cyclic dinucleotides (Stimulator of Interferon Genes, Sting) in Drosophila, promote Coxiella infection and increase host survival [73,74]. At least three T4SS-secreted effectors AnkG, CaeA, and CaeB inhibit intrinsic apoptosis in host cells, with CaeA also capable of inhibiting extrinsic apoptosis [75–79], and reviewed in [80]. Further, host pro-survival kinases Akt and Erk1/2 potentiate Coxiella’s anti-apoptotic activity [81]. These studies demonstrate that inhibition of programmed cell death is a key mechanism of intracellular adaptation of Coxiella. Typically, autophagy is associated with targeting pathogens for lysosomal degradation. However, autophagy and ATG gene functions are subverted by Coxiella for homotypic fusion of CCVs into a unified, spacious compartment [23,82].
Cells such as mast cells and neutrophils also contribute to bacterial control through secretion of antimicrobial peptides and control of bacterial dissemination [83,84]. Natural Killer (NK) cells limit intracellular bacterial replication and may protect from adverse histopathology, but release infectious Coxiella during degranulation [53,85].
Coxiella research: Where are we headed?
We are bearing witness to remarkable developments in the field of Coxiella research, including the formulation and refinement of defined axenic media, the availability of genomic sequences, lab-adapted strains, and the development of genetic tools [8,86–89], making Coxiella increasingly accessible to investigate fundamental host-Coxiella interaction (Figs 1 and 2). Each experimental host model is associated with distinct advantages and limitations as highlighted in Fig 1 and this extensive review [90]. This expanding landscape of experimental systems and tools is expected to facilitate addressing several key questions in future, as listed in Fig 1 and S1 Box, address public, veterinary and One Health perspectives and better modeling of disease biology in natural hosts [91].
Supporting information
S1 BoxKey open questions regarding the immune mechanisms against *Coxiella burnetii.*The expanding knowledge, experimental toolkit and models available for Coxiella research (Fig 1) is expected to facilitate addressing several key questions in future, as listed in S1 Box. These questions represent broad areas of innate immunity, organelle biology and vesicle traffic, effector biology, stress response, infection and immunity-induced cell death, adaptive immunity, disease manifestation and strain/clinical variant-specific response and more. Created with Biorender.com.(TIF)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Maurin M, Raoult D. Q fever. Clin Microbiol Rev. 1999;12((4):518–53. doi: 10.1128/cmr.12.4.51810515901 PMC 88923 · doi ↗ · pubmed ↗
- 2Eldin C, Mélenotte C, Mediannikov O, Ghigo E, Million M, Edouard S. From Q fever to Coxiella burnetii infection: a paradigm change. Clin Microbiol Rev. 2017;30(1):115–90. 10.1128/CMR.00045-16 27856520 PMC 5217791 · doi ↗ · pubmed ↗
- 3NIAID Biodefense Pathogens | NIAID: National Institute of Allergy and Infectious Diseases [Internet]. [cited 2025 Feb 15]. Available from: https://www.niaid.nih.gov/research/niaid-biodefense-pathogens
- 4Bioterrorism and Q fever: The Threat | Q Fever | CDC [Internet]. [cited 2025 Feb 15]. Available from: https://www.cdc.gov/q-fever/bioterrorism/
- 5Schneeberger PM, Wintenberger C, van der Hoek W, Stahl JP. Q fever in the Netherlands—2007-2010: what we learned from the largest outbreak ever. Med Mal Infect. 2014;44(8):339–53. doi: 10.1016/j.medmal.2014.02.006 25108615 · doi ↗ · pubmed ↗
- 6Anderson A, Bijlmer H, Fournier P-E, Graves S, Hartzell J, Kersh GJ, et al. Diagnosis and management of Q fever—United States, 2013: recommendations from CDC and the Q Fever Working Group. MMWR Recomm Rep. 2013;62(RR-03):1–30. 23535757 · pubmed ↗
- 7Moses AS, Millar JA, Bonazzi M, Beare PA, Raghavan R. Horizontally acquired biosynthesis genes boost Coxiella burnetii’s physiology. Front Cell Infect Microbiol. 2017;7:174. doi: 10.3389/fcimb.2017.00174 28540258 PMC 5423948 · doi ↗ · pubmed ↗
- 8Sandoz KM, Beare PA, Cockrell DC, Heinzen RA. Complementation of arginine auxotrophy for genetic transformation of Coxiella burnetii by use of a defined axenic medium. Appl Environ Microbiol. 2016;82(10):3042–51. doi: 10.1128/AEM.00261-16 26969695 PMC 4959063 · doi ↗ · pubmed ↗